| CATEGORII DOCUMENTE |
| Agricultura | Asigurari | Comert | Confectii | Contabilitate | Contracte | Economie |
| Transporturi | Turism | Zootehnie |
UNIVERSITATEA DE STIINTE AGRICOLE SI MEDICINA VETERINARA
A BANATULUI TIMISOARA
FACULTATEA DE HORTICULTURA
FIZIOLOGIE VEGETALA
R e f e r a t
UTILIZAREA HORMONILOR DE CRESTERE
LA SPECII HORTICOLE

CUPRINS
INTRODUCERE..................
FITOHORMONI..................
Acid abscisic..................
Auxine......................
Citokinine...................
Etilen.....................
Gibereline...................
Fitocrom.....................
RETARDANTI....................
INHIBITORI....................
1. INTRODUCERE
Actiunea constienta a omului de influentare a naturii s-a inscris, de-a lungul timpului, ca o coordonata a existentei, a deverii sale. Agricultura, ca sector de baza a activitatii umane, este chemata sa raspunda cerintelor crescande de produse, atat sub raportul cantitatii, al calitatii , cat si al diversitatii, impunandu-se ca o necesitate trecerea la forme eficiente, intensive, de mare randament, ale cultivarii pamantului, resursa limitata ca intindere, inepuizabila insa ca potential, prin forta creatoare a mintii omenesti.
Posibilitatile de crestere a productiei agricole, de reglare a structurii acesteia, evolueaza odata cu nivelul de cunostinte , cu experienta acumulata, cu perfectionarea aparaturii de cercetare si a tehnologiilor de cultura a plantelor.
Descoperirea substantelor cu actiune regulatoare asupra cresterii fitohormonilor, stimulatorii de crestere , inhibitorii, retardantii - a pus la indemana specialistilor un instrument eficace, de un deosebit rafinament,pentru dirijarea si controlul proceselor de crestere si dezvoltare ale plantei, ale productiei vegetale.
In acest context, consideram util sa nominalizam la genul proxim ca substante de reglare a cresterii toti compusii organici naturali sau sintetici, care prezinta , in cantitati mici, efecte asupra ritmului si proportiilor de crestere. Diferente specifice dintre substantele care se incadreaza in aceste categorii constau in efectul particular al actiunii (stimulare - franare, inhibitie - activare).
Fitoregulatorii de crestere sunt substante biologic active - naturale sau de sinteza - care influenteaza procesele de crestere si dezvoltare ale plantelor, marind in general rezistenta acestora la actiunea nefavorabila a daunatorilor si a unor factori de stres. Din aceasta grupa de substante vom trata fitohormonii, retardantii, inhibitorii si fitocromul.
Fitoregulatorii de crestere au un spectru larg de actiune. Pe langa reglarea proceselor de crestere si dezvoltare, ei pot influenta si alte procese biochimice si fenomene biologice (germinarea, pornirea sau inhibarea mugurilor , a tuberculilor si bulbilor, randamentul fotosintezei, circulatia apei, a sevei brute si elaborate, absorbtia si transportul substantelor anorganice si organice , rezistenta la cadere a frunzelor si fructelor , inflorirea si fecundarea, rezistenta plantelor in conditii de stres, miscarea stomatelor etc.).
(JUNG.1979, 1984; 1983; MILICA si colab, 1983: SZIRTES 1984; CARR,1972; PILET, 1961 etc).
In practica agricola, fitoregulatorii de crestere sunt utilizati in cantitati mici - de ordinul ppm.
FITOHORMONI
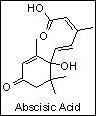 2.1. ACIDUL ABSCISIC
2.1. ACIDUL ABSCISIC
Acidul abscisic este un inhibator natural important, larg raspandit in organismele vegetale, care, impreuna cu substantele stimulatoare de crestere, in special cu auxinele, giberelinele si citochininele, regleaza procesele de crestere si dezvoltare ale plantelor (BONNER si colab.1976; MILICA si colab.1983). Aceste inhibtor engogen are o activitate biologica foarte puternica. Este initiatorul si ,,controlorul" proceselor de imbatranire la plante. Inhiba cresterea si dezvoltarea plantelor, grabeste abcizia sau caderea fructelor si a frunzelor , determina repausul mugurilor si al semintelor, inchiderea si deschiderea stomatelor etc. Inhinba biosinteza acizilor nucleici si a proteinelor. Opreste germinatia semintelor si pornirea mugurilor in vegetatie. Etc.
Descoperirea acidului abscisic s-a datorat unor cercetatori independente efectuate in principal de doua colective de cercetare; unui in S.U.A. condus de F.T. ADDICOTT si colab, iar celalalt in Anglia, condus de P.F. WAREING si colab. (MOORE 1979). Colectivul din S..U.A. a lucrat in principal pe bumbac. In 1961 , acest colectiv a extras din capsule mature de bumbac un compus cristalin, cu structura nedeterminata, denumit abscisin I, care accelereaza abscizia frunzelor de bumbac. Ulterior , acelasi colectiv in 1963, a descoperit un nuo inhibitor endogen, in fructele tinere (necoapte) de bumbac, denumit abscisin II, care accelereaza abscizia fruntelor si produce caderea fructelor tinere. Proprietatile fizice si chimice, precum si formula moleculara (C15H20O4) a acestui comus au fost determinate de OHKUMA si colab. In 1963.
La scurt timp dupa descoperirea si stabilirea proprietatilor celor doi inhibitori endogeni - abscisin II si dormin au comparat proprietatile )masa moleculara, punct de topire, spectre de absorbtie etc) dorminului extras din frunze de Acer pseudoplatanus, cu cele abscisinului II si ai constatat ca cei doi compusi sunt identici. Structura moleculara stabilita pentru abscisin II de catre OHKUMA si colab. 1965 , cu un atom de carbon asimetric la C-1 , a fost atribuita si pentru dormin. Adaptandu-se pentru ambii o denumire comuna de acid abscisic cu o prescurtare internationala de ABA. Intrucat acidul abscisic contine un atom asimetric poate forma doi enantiomeri: (+) -ABA si (-) -ABA sau (S) -ABA si ( R ) -ABA (fig.1.)
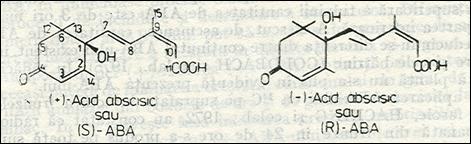
Fig.1 Enantiomeri ai acidului anscisic
Acidul abscisic natural este dextrogir (+)-ABA sau (S) -ABA. Cel obtinut prin sinteza este un amestec racemic ()-ABA sau (SR)-ABA.
Acidul abscisic este un inhibator endogen care sub aspectul structurii moleculare face parte din clasa compusilor terpenoidici, Este o sesquiterprena, cu formula moleculara C15H20O4, fiind formata din trei unitati izoprenice. Este genetic inrudit cu sterolii, pigmentii carotenoidici, cu compusii monoterpenici, diterpenici, triterpenici si politerpenici.
Proprietati
Acidul abscisic natural este o substanta solida, cristalina, solubila in apa si in solutii alcoolice, cu p.t. 160-161 0C. Acidul abcisic obtinut prin sinteza () -ABA are p.t. 191oC. In identificarea si determinarea acizilor abscisici spectrele de absorbtie in U.V. si I.R. au un rol important. Spectrul de absorbtie al ABA-ului depinde de pH-ul mediului. In mediu acid (+) -ABA prezinta un maxim de absorbtie in U.V. la 262 nm, cu o inflexiune la 240 nm. In mediu bazic, maximul de absorbtie apare la 245 nm, fiind intens cu 20% (MILBORROW, 1974). Spectrul de absorbtie al esterilor nu este influentat de pH. Puterea optica rotatorie este de asemenea influentata de pH, fapt pentru care se masoara in mediu acid, intr-o solutie de H2SO4. Dicroismul celular masurat in solutie metanolica acida de 0,005 n H2SO4 prezinta urmatoarele date:
∆ε 263 nm = +395; ∆ε 230 nm = - 34; ∆ε 318 nm = - 2,5;
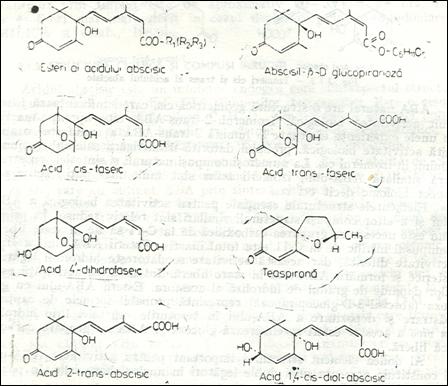
Derivati naturali ai acidului abscisic (Dupa MOORE,1979)
Mecanismul de actiune al acidului abscisic
Mecanismul molecular de actiune al acidului abscisic in organismele vegetale inca nu este pe deplin cunoscut, cu toate ca s-au obtinut progrese remarcabile in ultimele doua decenii in ce priveste raspandirea in natura, cunoasterea structurii moleculare, a rolului si a functiilor acidului abscisic in diferite plante. Se stie ca ABA este un fitohormon important care stimuleaza procesele de imbatranire, marind totodata rezistenta plantelor aflate in stare de stres. ABA este fitohormonul care stimuleaza supravietuirea plantelor aflate in conditii nefavorabile de viala. Continutul sau creste foarte mult, de zeci si sute de ori, in tesuturile sau organele lezate , sau in cele aflate in stare de stres, datorita secetei si temperaturilor ridicate, gerului , inundatiilor, lipsei din mediul de nutritie a unor elemente minerale (N, K+, Ca2+, O etc), transplantarii plantelor dintr-un loc in altul etc.
Acidul abscisic are o actiune biochimica si fiziologica contrara principalilor fitohormoni (auxine, gibereline, citochinice) care stimuleaza procesele de crestere si dezvoltare ale plantelor, fiind unul din cei mai importanti inhibitori endogeni. Cresterea si dezvoltarea plantelor este reglata de raportul existent in plante dintre continutul fitohormonilor de crestere si cel al acidului abscisic si a altor inhibitori. Aceasta ,,balanta" biochimica determina cresterea si dezvoltarea plantelor in diferite fenofaze si anotimpuri in zonele temperate.
Pe baza unor numeroase date experimentale, a functiilor sale si a efectelor ce le produce asupra unor procese biologice s-a stabilit ca ABA actioneaza in plante ca inhibitor endogen prin intermediul mai multor mecanisme biochimice.
Inhibarea cresterii si dezvoltarii plantelor se realizeaza prin aceea ca inhiba biosinteza acizilor ribonucleici, in special a m-ARN, blocand procesul de transcriptie si de translatie a informatiei genetice din molecula ADN-ului. Fiind blocate aceste procese biochimice se opreste biosinteza proteinelor si a altor compusi importanti din celule (glucide, lipide, etc.). este blocat procesul de biosinteza a enzimelor care determina si controleaza procesele biochimice din organism. In felul acesta se reduc in mod treptat procesele de fotosinteza , respiratia, fosforilarea oxidativa etc. ABA determina atat reducerea proceselor de biosinteza din organismele vegetale cat si a celor generatoare de energie. In aceste procese de inhibare ABA se comporta ca un represor care blocheaza activitatea genelor structurale din molecula ADN, inhiband intr-o proportie mai mare sau mai mica, in functie de continutul sau, procesele de biosinteza din plante.
Un alt mecanism biochimic de actiune a ABA-ului in plante il constituie reglarea fluxului ionilor anorganici prin membrane biologice, prin schimbarea permeabilitatii acestora si a potentialelor biolectrice. Acest mecanism se remarca cu usurinta in frunze, in celulele care determina inchiderea si deschiderea stomatelor prin reglarea continutului K+. Cand stomatele se inchid celulele cedeaza ionii de K+ , iar cand stomatele se deschid celulele acumuleaza ionii de K+. Ionii de Ca2+ au un rol insemnat in abcizia fructelor si a frunzelor. In perioada cand plantele se afla in stare de stres ABA se acumuleaza in radacini, determina o depolarizare partiala a membranelor, fapt ce le mareste permeabilitatea, usurand procesul de absorbtie a apei si a sarurilor minerale. ABA in aceste procese se comporta ca o pompa de electroni avand efecte contrare citochininelor. ABA stimuleaza transportul apei si a sarurilor minerale de la radacini spre varful plantei, furnizand plantelor cat mai multe elemente nutritive. Prin inchiderea stomatelor si stimularea absorbtiei apei si a sarurilor minerale ABA contribuie la supravietuirea plantelor aflate in stare de stres o perioada cat mai indelungata. In aceasta actiune ABA contribuie pe de o parte la reducerea proceselor de biosinteza, iar pe de alta parte, prin stimularea absorbtiei si a transportului substantelor minerale, alimenteaza plantele cu hrana minima pentru a supravietui starii de stres.
Plantele mutante cu continut redus de ABA endogen, in aceleasi conditii ecologicem absorb mai putin apa si saruri minerale decat plantele cu continut ridicat de ABA, fapt pentru care au o perioada mai scurta de supravietuire in conditii de stres.
Un alt mecanism de actiune al ABA-ului in procesele de inhibare a germinarii semintelor, incoltirii cartofilor si a bulbilor il constituie inhibarea unor enzime implicate in aceste procese, in special a hidrolazelor (amilaze, esteraze, proteaze, ribonucleaze etc).
Intrarea in repaus a mugurilor si inhibarea pornirii in vegetatie a acestora se datoreste acumularii acidului abscisic care determina o inhibare a proceselor de biosinteza si de fotosinteza, prin inhibarea activitatii unor carboxilaze, ribonucleaze, fosfataze, oxidoreductaze etc.
Prin tratarea mugurilor de Coleus cu ABA a scazut simtitor continutul lipidelor. Acumularea ABA in tesuturile fotosintetizante determina o degradare a clorofilelor, inhibarea procesului de fotosinteza, ingalbenirea plantelor, cresterea continutului carotenoidelor oxigenate.
Acidul abscisic prin rolul si functiile ce le indeplineste in organismele vegetale consituie un inhibitor endogen foarte important care influenteaza adaptarea plantelor la conditii de mediu schimbate, contribuind prin mecanismul sau de actiune la supravietuirea acestora in conditii neprielnice de crestere si dezvoltare.
Rolul si functiile acidului abscisic.
Acidul abscisic este un fitohormon foarte important pentru organismele vegetale, cu rol multiplu si complex, cu numeroase functii , contribuind in mod substantial la reglarea proceselor de crestere si dezvoltare ale plantelor, la cresterea rezistentei plantelor aflate in conditii de stres. ABA are un rol hotarator in determinarea repausului la muguri, seminte, bulbi, cartofi, rozomi etc. , in inhibarea germinatiei semintelor etc. Stimuleaza procesele de imbatranire, caderea si a frunzelor. Are actiune antagonista fata de cea a fitohormonilor care stimuleaza cresterea si dezvoltarea plantelor.
Grabirea proceselor de imbatranire a plantelor. Functia caracteristica a acidului abscisic este aceea de a grabi procesele de imbatranire a plantelor, scurtand astfel ciclul biologic de crestere si dezvoltare al acestora. Frunzele turgescente ale diferitelor plante tratate cu o solutie de ABA se ingalbenesc repede si in 2-3 zile se ofilesc, dupa care urmeaza moartea acestora. Daca frunzele tratate cu ABA se spala dupa 1-2 zile ele isi revin treptat la starea initiala si isi continua procesul de crestere. Pe baza acestor experiente se poate trage concluzia ca ABA nu este toxic ca atare, decat in concentratie foarte mare. Ofilirea in imbatranirea frunzelor s-a datorat inhibarii biosintezei acizilor nucleici, a proteinelor si a numeroase enzime care dirijeaza procesele metabolice din organism. In plantele netratate imbatranirea celulelor si a tesuturilor se produce in mod treptat , proportional cu intensificarea biosintezei si acumularii ABA-ului. In mediile biologice care stimuleaza degradarea acidului abscisic plantele au o crestere mai mare, intrucat actiunea inhibanta a ABA-ului este mai redusa.
Franarea cresterii si stimularea imbatranirii plantelor se produce atat la plantele intregi cat si la seminte, muguri, tesuturi si organe izolate.
Cresterea rezistentei plantelor in conditii de stres. O alta functie importanta a ABA-ului o constituie marirea rezistentei plantelor aflate in conditii de stres, indiferent de natura factorului stresat. Aceasta proproetate a sa se poate realiza pe baza modificarii permeabilitatii membranelor celulare, usurarii absorbtiei apei si a substantelor minerale si a transportului acestora prin celulele si tesuturi, reducerii la minimum a proceselor de biosinteza si degradare prin inhibitie enzimatica, oprirea pierderilor de substante din corpul plantelor prin reglarea stomatelor, inducerea repausului la muguri si seminte etc. Continutul ABA din tesuturile si organele stresante creste foarte mult atat datorita intensificarii proceselor sale de biosinteza , cat si eliberarii din compusii de depozitare (esteri, eteri etc). In plantuele de fasole aflate in stres de umiditate, alaturi de cresterea continutului ABA-ului creste in continutul acidului faseic si a acidului dihidrofaseic intr-o proportie mai mare decat a ABA-ului.
In frunzele de avocado nevatamate continutul mediu al ABA-ului este de 20 μg/kg, iar in urma vatamarii frunzelor se intensifica biosinteza si acumularea ABA-ului si intr-un timp scurt de 10 - 12 minute continutul sau este de 2600 μg/kg. In fructele de avocado care au frecvent un continut ridicat de ABA de 4000 - 6000 μg/kg continutul ABA-ului creste intr-o proportie redusa daca felii de fructe sunt puse in cinditii de stres, desi se intensifica biosinteza ABA-ului pe calea malonil CoA. In radacinile de avocado continutul acestui fitohormon este scazut de 27 μg/kg si creste putin prin efecte de stres, dar descreste rezistenta radacinilor la aborbtia apei si a sarurilor minerale protejand frunzele de deshidratare si ofilire.
Dintre elementele minerale care intarzie imbatranirea plantelor un rol mai important revine azotului si potasiului. Lipsa acestor elemente din mediul de nutritie a plantelor le intensifica procesele de imbatranire. Daca la floarea soarelui, in varsta de 7 - 8 saptamani se sustrage azotul din mediul de nutritie pentru o saptamana, continutul ABA-ului creste de 3,7 ori in frunzele batrane, de 3,1 ori in cele mature si de 1,8 ori in cele tinere , fata de proba martor. In varful tulpinii continutul ABA-ului a crescut de 2,6 ori. Intre continutul azotului si al ABA-ului exista o relatie invers proportionala.
Continutul acidului abscisic creste mult in plantulele rasadurilor care sunt transportate. La tomatele de 17 zile, chiar si la transplantarea cea mai rapida cantitatea de ABA din plantule a crescut de 10 ori. Cresterea continutului de ABA determina franarea cresterii lastarilor pana ce radacinile se regenereaza si vatamarile suferite dispar. Pe drept cuvant ABA este numit fitohormonul adaptarii plantelor si a supravietuirii acestora in conditii ecologice nefavorabile.
Mentinerea si reglarea umiditatii. Acidul abscisic are un rol important in mentinerea si reglarea umiditatii plantelor prin reglarea inchiderii si deschiderii stomatelor, ca mecanism protector fata de stresul de apa. Pe baza multor teste experimentale s-a constatat ca inchiderea si deschiderea stomatelor depinde de variatia continutului ABA-ului din celulele de garda, care prin modificarea presiunii osmotice si a columului determina inchiderea si deschiderea stomatelor, regland astfel transpiratia si eliminarea apei.
Prin stropirea unor frunte de Zea mays, Rumex obtusifolia, Beta vulgaris cu ABA exogen, in concentratie de 1 M, s-a redus transpiratia si s-au inchis stomatele in decurs de 3-9 minute de la aplicarea tratamentului. Celulele de garda din frunze sunt foarte sensibile la actiunea acidului abscisic. Ele se inchid cand ABA are o concentratie de 10 -7 M. Cresterea continutului endogen de acid abscisic creste in frunze odata cu pierderea apei si se intensifica pe masura accentuarii stresului de apa. Continutul sau creste intr-o perioada scurta de 10 - 12 minute de zeci si sute de ori inaintea ofilirii frunzelor si inaintea inchiderii stomatelor.
ABA acumulat in celulele de garda determina eliminarea ionilor de K+, stomatele se inchid , transpiratia se opreste, protejand astfel plantele de uscare. Se produce totodata o crestere a continutului de CO2. Daca planta primeste apa si stresul hidric inceteaza, scade concentratia ABA-ului in celulele de garda , ionii de K+ sunt recuperati, concentratia CO2 scade si stomatele se deschid permitand schimbul de gaze, intensificarea proceselor biologice (fotosinteza, respiratie etc).
ABA, in perioada de stres, determina inhibarea amilazelor, reducerea hidrolizei amidonului, fapt ce determina scaderea presiunii osmotice si a turgescentei in celulele stomatice.
La plantele de porumb rezistente la seceta concentratia endogena a ABA-ului este de 2,5 ori mai mare decat la plantele nerezistente fata de acest stres, iar la iarba de Sudan concentratia ABA-ului este de 2 ori mai mare la formele rezistente la seceta si arsita decat la cele nerezistente. La o mutanta de tomate ,,Flacca concentratia ABA-ului endogen este de 10 ori mai scazuta decat la soiul normal ,,Rheinlands Ruhm", din aceasta cauza activitatea citochininelor in mutanta este ridicata deoarece stomatele au ramas tot timpul deschis si n-au putut regla pierderile de apa.
Prin administrarea de ABA plantele mutante au primit aspectul unor plante normale. ABA exogen a provocat inchiderea stomatelor, care nu s-au mai supus controlului genetic. In perioada de seceta continutul ABA din frunze creste de aproape 40 de ori fata de proba martor. Marirea rezistentei la seceta a plantelor si reglarea umiditatii acestora se realizeaza prin reducerea evapotranspiratiei prin mecanismul de inchidere si deschidere a stomatelor.
Reglarea starii de repaus la muguri. Repausul la muguri, seminte, cartofi, bulbi, rizomi etc. se regleaza endogen, cu ajutorul unor inhibitori, fiind o etapa obligatorie in ciclul biologic de crestere si dezvoltare a plantelor.
Pe baza mai multor observatii si date experimentale efectuate s-a constatat ca intrarea mugurilor in stare de repaus la Fraxinus si la tuberculii de cartofi este corelata cu acumularea unor inhibitori de crestere , neidentificati sub aspectul structural. Rezultate similare s-au obtinut ulterior si la Beta pubescens si la alte specii de plante lemnoase. Dupa descoperirea acidului, abscisic s-a constatat ca acest fitohormon are rolul principal, hotarator in determinarea starii de repaus la muguri si seminte. Intrarea mugurilor in stare de repaus se datoreste intensificarii biosintezei ABA in frunze, in concitii de iluminare de zi scurta de toamna, migrarea si acumularea acestuia in mugurii latenti si lastari. Prin faptul ca ABA inhiba procesele de biosinteza a acizilor ribonucleici si a proteinelor si blocheaza activitatea a numeroase enzime, in muguri se instaleaza starea de repaus cand procesele metabolice fundamentale sunt reduse la minimum. Perioada de repaus la plantele perene din zona temperata dureaza si pana in a doua jumatate a iernii (luna februarie ). Revenirea din starea de repaus necesita ca plantele sa suporte in aceasta perioada de latenta temperaturi joase, la multe plante perene intre 1-10C, de la 260 la 1000 de ore. Frigul este necesar pentru mentinerea si intreruperea starii de repaus la muguri, iar temperaturile ulterioare mai ridicate sunt necesare pentru pornirea in vegetatie dupa perioada de repaus. La sfarsitul lunii ianuarie si in luna februarie datorita temperaturilor scazute si umiditatii ridicate se produce o degradare treptata a ABA-ului si a altor inhibitori engogeni si o crestere a continutului stimulatorilor de crestere , in special a giberelinelor. Perioada de repaus se intrerupe mai tarziu, plantele raman intr-o stare de post repaus pana la inceputul primaverii, cand continutul ABA-ului s-a redus foarte mult, fapt ce permite pornirea mugurilor in vegetatie. In perioada de post repaus raportul dintre continutul stimulatorilor de crestere (gibereline, auxine, citochinine) si continutul ABA devine supraunitar. Repausul plantelor este un proces endogen supus controlului genetic. El nu se datoreaza modificarilor ciclice care apar la unii factori naturali.
La Betula pubescens si Acer pseudoplatanus repausul poate fi provocat de fotoperioade scurte si nopti lungi si inceteaza prin expunerea plantelor la fotoperioade lungi si nopti scurte, fara sa fie supuse plantele temperaturilor scazute. In perioadele de zi scurta creste continutul ABA din frunze si muguri. Extractul din frunze de mesteaca (Beta pubescens) tinute in conditii de zile scurte s-a aplicat pe mugurii si frunzele aceleiasi specii de mesteacan mentinute in conditii de iluminare de zi lunga, determinand oprirea cresterii si intrarea jugurilor in stare de repaus si la aceste plante . in extractul de mesteacan se gasea acid abscisic care are un rol important in reglarea cresterii si a repausului la plante. Starea de repaus a mugurilor este determinata de continutul ridicat al ABA si de nivelul redus al stimulatorilor de crestere, situatia inversa determina incetarea starii de repaus.
Plantele lemnoase din zonele temperate isi formeaza mugurii de repaus la sfarsitul verii si toamna, cand planta intreaga intra in stare de repaus, datorita acumularii in frunze si muguri a acidului abscisic. Repausul mugurilor apare si la plantele tropicale, la care fotoperioada se schimba insesizabil si schimbarile de temperatura sunt blande. Prezenta repausului mugural si la plantele tropicale si din alte zone geografice demonstreaza ca repausul este o stare fiziologica obligatorie in viata plantelor, ca starea de repaus nu este un raspuns al plantelor la schimbarile fizice ale mediului, ca repausul este un fenomen adaptiv care pregateste plantele pentru supravietuirea lor in timpul conditiilor de viata nefavorabile.
Reglarea starii de repaus la seminte. Repausul seminal este determinat de acumularea acidului abscisic in fructe si in seminte. Semintele multor specii de plante nu germineaza imediat dupa coacere chiar daca sunt puse in conditii favorabile.
ABA, printr-un mecanism similar determinarii repausului la muguri, mentine starea de repaus a semintelor si impiedica germinarea prematura a acestora. In cazul semintelor , ABA inhiba atat biosinteza acizilor nucleici a proteinelor, cat si activitatea amilazelor, orpindu-se astfes hidroliza amidonului.
La fel ca si la muguri starea de repaus la seminte face parte din ciclul biologic de crestere si dezvoltare a plantelor si asigura supravietuirea acestora. Impermeabilitatea cojii fata de apa si schimbul de gaze, nematuritatea embrionului, necesitatea ,,coacerii ulterioare" in depozite speciale, rezistenta mecanica a cojii, prezenta inhibitorilor in coaja, in embrion sau in masa fructului, necesitatea luminii sau a intunericului; necesitatea temperaturilor joase si a umiditatii etc.
Reglarea caderii fructelor si a frunzelor. Una dintre functiile importante ale acidului abscisic, de la care ii deriva numele, este aceea a determinarii absciziei fructelor si a frunzelor. Pe baza mai multor date experimentale s-a stabilit existenta unei corelatii directe continutul ABA-ului, abscizia fructelor tinere si a dehiscentei fructelor matura. Fructele care cad au continut mai mare de ABA decat fructele rezistente la cadere. In abscizia fructelor ABA este activ numai concentratii mari. Abscizia fruntelor este stimulata intr-o proportie insemnata de etilena. Continutul etilenei creste concomitent cu imbatranirea frunzelor. Daca antitatea de apa din plante scade de la 20% la 50% continutul etilenei creste de 7 ori , iar la 2 zile de la udarea plantei scade la loc. In zona de abscizie a frunzelor scade continutul ionului de calciu cu putin timp inainte de caderea lor.
In producerea absciziei la fructe si frunze ABA stimuleaza formarea stratului de separare care determina caderea fructelor si a frunzelor. Se intensifica activitatea celulazelor si a pectinazelor si se produce degradarea lamelei mediene din membrana celulara. Si in acest rolul important il are raportul dintre continutul inhibitorilor si a stimulatorilor de crestere. Abscizia fructelor si a frunzelor este influentata nu numai de actiunea factorilor interni ci si a celor externi (vant, brume , ger etc).
Tratarea pomilor fructiferi in diferite fenofaze cu solutii de ABA ar contribui la rarirea chimica a florilor si a fructelor , fapt ce ar asigura uniformizarea marimii fructelor, cresterea calitatii acestora, sporirea productiei la hectar, evitarea rodirii periodice.
ABSCISINE.
Abscisinele constituie un grup de compusi naturali si sintetici care sunt inruditi structural si functional cu acidul abscisic. Ele inhiba procesele de crestere si dezvoltare ale plantelor, maresc rezistenta acestora in conditii de stres, contribuie la determinarea starii de repaus la muguri si seminte, inhiba germinatia etc. Ele indeplinesc in plante, intr-o proportie mai mare sau mai mica toate functiile acidului abscisic. Dintre abscisinele naturale, care in procesul de biosinteza a ABA-ului apar ca precursori, un rol important revine xantoxinului.
XANTOXINUL
Xantoxinul este un inhibitor endogen puternic , cu structura si activitate biologica similara acidului abscisic, dar cu o raspandire mai limitant in organismele vegetale. Se gaseste in cantitate mai mare predominant in lastarii tineri. Mecanismul intim de biosinteza a xantoxinului in diferite plante nu se cunoaste, dar numerosi cercetatori afirma ca acest compus este precursorul direct al acidului abscisic.
2.2. AUXINE
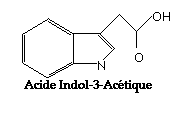
Cu toate ca nu se poate face o ierarhizare generala a importantei unui anume tip de fitohormon, auxinele prezinta un interes aparte fiind prima clasa de substante stimulatoare de crestere descoperita - prin reprezentantul cel mai edificator - acid 3-indolilacetic.
Numele de auxina deriva de la grecescul auxo - auxein, care se traduce prin: a face sa creasca.
Structura si proprietati
Acidul 3-indolilacetic este un derivat heterociclic al acidului acetic. Prima sinteza a fost realizata in anul 1904 de catre Ellinski care a pornit de la fenilhidrazona esterului al acidului b-formilpropionic, care in conformitate cu metoda Fischer de sinteza a derivatilor indolici, se va cicliza, rezultand esterul metilic al acidului 3-indolilacetic, care prin hidroliza va elibera acidul 3-indolilacetic. Fig.3.
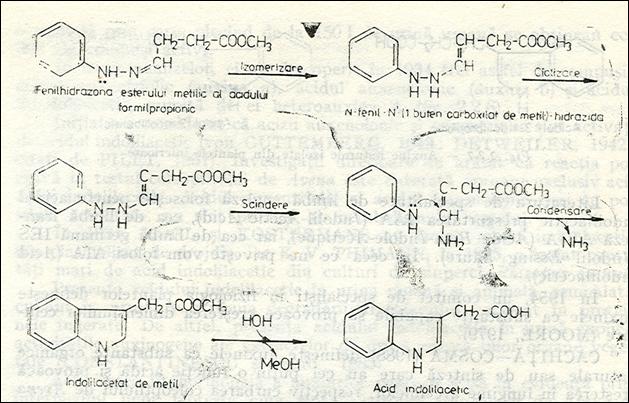
Fig.3. Sinteza Fischer a acidului indolilacetic realizata de ELLINGER in 1904.
Acidul 3-indolilacetic sau scatol carboxilic este un acid indolic cu gruparea carboxilica situata in afara nucleului. Formula moleculara este C10H9O2N, iar masa moleculara 175,18 UMA. Se prezinta sub forma de cristale aciculare la temperatura normala, al caror punct de topire este diferit in functie de mediul de recristalizare folosit : 164-165 la recristalizare din benzen , 167-168 la recristalizare dintr-un amestec de alcool etilic si clorura de etilen si 168-169 la recristalizare din clorura de etilen.
La incalzire peste temperatura de topire are loc descompunerea sa prin decarboxilare, la scatol, un compus de catabolism al triptofanului.
Acidul 3-indolilacetic este greu solubil in apa dar se solva usor in solventi organici precum clorura de etilen, alcool etilic, acetona, cloroform si eter etilic. Solubilitatea in n-butanol si benzen este notabila doar la cald. Pe aceasta solubilitate diferentiata se bazeaza si purificarea acidului 3-indolilacetic prin recristalizare.
Prezenta gruparii carboxilice confera activitate moleculei, care in solutie va pune in libertate ioni de hiddrogen. Totodata, in molecula se mai gasesc o grupare bazica, reprezentata de azotul indolic. In consecinta, , molecula va avea o comportare amfotera. Punctul izolectric situat la pH 4,78 arata ca gruparea acida este mai puternica decat cea bazica.
Prezenta celor doua grupari active, carboxil si amino, precum si proprietatile conferite de nucleul indolic, fac din molecula de acid 3-indolilaceticun substrat foarte activ pentru reactii biochimice, care duc la inactivarea rapida a acestuia, atat ,,in vivo" cat si ,,in vitro".
Raspandire, forme libere si legate, transport.
Concentratiile extrem de scazute la care se gasesc acidul indolilacetic si derivatii sai au creat mult timp dificultati, atat in identificarea, cat si in dozarea acestora.
Majoritatea cercetarilor au folosit metoda extractiei substantei din planta proaspata cu ajutorul alcoolului, eterului etilic sau al acetatului de etil, sau determinari spectroluorimetrice si spectrometrice de masa.
Se poate afirma ca acidul indolilacetic este o substanta ubicvitar raspandita in lumea vegetala. S-au pus in evidenta auxine in toate organele plantelor superioare, dar concentratia acestora variaza in special cu varsta, dar si cu tesutul sau organul cercetat. Concentratii ridicate de auxina gasim la plante tinere.
In organismele vegetale auxinele se acumuleaza mai mult in tesuturile si organele cu activitate biochimica si fiziologica mai intensa, cum sunt: mugurii foliari si florali, frunzele tinere, varfurile tulpinilor si a radacinilor, polen, seminte imature, ovar fecundat, cotiledoane etc., predominant in partea aeriana a plantelor.
Acidul indolialactic se gaseste in plante fie ca atare, fie sub forma conjugata (condensata) cu alte molecule cu masa mare. Se pare ca aceadta forma de existenta reprezinta o modalitate de reglare a concentratiei auxinei active in plante. In cazul existentei unui exces de acid indolilacetic, o parte din acesta se stocheaza temporar in compusi conjugati mai putin , sau de loc activi. Pe masura ce nevoile plantei devin imperioase, prin procese hidrolitice va avea loc eliberarea dirijata de auxine.
Acidul indolilacetic se gasesc in mai multe stari chimice in tesuturile vegetale, sub forma libera, sub forma legata si in amestec cu diferiti alti fitoregulatori de crestere.
Rolul si functii biologice
Fitohormonii auxinici au un rol multiplu si complex in organismele vegetale si indeplinesc numeroase functii biologice. Auxinele actioneaza in toate etapele de crestere si dezvoltare ale plantelor.
Asa dupa cum s-a prezentat la mecanismul de actiune, auxinele au un rol esential in alungirea si cresterea celulelor prin marirea plasticitatii si permeabilitatii membranelor si a peretilor celulari. Prin modificarea permeabilitatii membranelor celulare se intensifica absorbtia apei si a unor ioni, in special a celor de K+, fiind favorizata desorbtia ionilor de H+ . uxinele determina atat alungirea membranelor celulare cat si ingrosarea acestora, prin depunerea de noi substante formative. In procesul de alungire a celulelor, sub influenta auxinelor, se produc modificari in ce priveste elasticitatea, plasticitatea si permeabilitatea membranelor celulare, orientarea microfibrilelor de celuloza.
Prin experiante cu butasi de mazare s-a aratat ca auxina stimuleaza formarea calusului, deci este stimulata diviziunea celulara si activitatea cumbiului. Auxina stimuleaza de asemenea lignificarea tesuturilor. Ea actioneaza in organismele vegetale atat singura cat si in asociere cu alti fitohormoni.
Tratamentele cu auxine maresc presiunea osmotica a sucului celular, absorbtia apei si a unor ioni, scad intensitatea solutiilor protoplasmatice, intensifica miscarea browniana, mobilitatea curentilor citoplasmatici etc. Auxinele stimuleaza reactiile de hidroliza ale poliglucidelor (amidon, celuloza, hemiceluloza, substante pectice) si ale propteinelor; influenteaza de asemenea biosinteza unor vitamine.
In functie de concentratie, de natura plantei si a fitohormonului, auxinele pot stimula sau frana germinatia semintelor. Ele stimuleaza germinarea semintelor pastrate un timp mai indelungat si a celor cu germinatie intarziata. Intervin de asemenea, in procesul de formare a organelor plantelor.
Auxinele naturale si cele de sinteza contribuie la cresterea si formarea radacinilor adventive. Ele se acumuleaza la partea bazala a butasilor si determina formarea radacinilor secundare.
Auxinele stimuleaza ,,germinarea " polenului, cresterea tuburilor polenice, fecundarea cosferei, stimularea diviziunii oului, formarea fructelor inclusiv a celor partenocarpice. Continutul ridicat de auxine din unele organe mentin mugurii pomilor, tuberculii cartofilor, rizomii si bulbii plantelor in stare de latenta.
In ce priveste inflorirea plantelor, s-a stabilit ca, in general, tratamentele cu auxine stimuleaza inflorirea plantelor de zi lunga si inhiba inflorirea speciilor de zi scurta.
La numeroase plante (mar, par, cais, tomate, vinete, pepeni, fragi, struguri, afine etc) tratamentele cu auxine sintetice aplicate in pasta de lanolina, determina formarea de fructe partenocarpice, fara seminte , la care nu s-a produs fenomenul de polenizare.
Auxinele au un rol insemnat in miscarile de crestere si in tropisme.
Auxinele franeaza caderea frunzelor si a fructelor, sunt implicate in corelatiile dintre diferite organe si tesuturi. Caderea frunzelor si a fructelor se produc in perioadele in care continutul auxinelor endogene scade foarte mult.
Raportul dintre continutul auxinelor si al giberelinelor influenteaza sexul florilor. Daca raportul dintre continutul gibereline/auxine este mare se produc flori mascule, iar daca acest raport este mic apar flori femele.
Auxinele stimuleaza fotosinteza, fixarea si transformarea CO2 in substante organice, migrarea sevei elaborate din frunze in alte organe ale plantelor.
Aplicatii practice.
Auxinele au numeroase si variate aplicatii practice atat in ce priveste cultura cerealelor, a legumelor si fructelor , care reprezinta baza alimentara pentru lumea intreaga, cat si pentru flori, plante tehnice, arbori si arbusti, plante ornamentale etc.
La mazare, tratamentele cu auxine aplicate foliar inainte de inflorire, in concentratie de 10-4M , determina o marire a volumului pastailor si boabelor, creste continutul de azot si fosfor in boabe si pastai, fara sa influenteze forma acestora.
Prin tratarea mazarei cu solutii apoase de acid indolilacetic, de 0,1 - 20 ppm, se produce o crestere accentuata a tulpinii si chiar a radacinii.
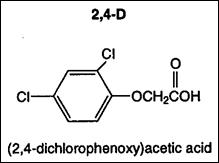
Umectarea boabelor de fasole cu o solutie de 10 mg/l AIA are ca efect franarea cresterii radacinilor in primele 10 zile de la aparitia acestora, dar sporeste cresterea perisorilor absorbtivi radiculari . Prin tratarea plantuelor de fasole cu solutii diluate de 2,4 - D, in medie de concentratie 1 ppm, se obtin sporuri de productie si creste continutul vitaminei C in pastai in boabe.
La tomate , acidul indolilacetic in concentratie mica de 1-10 ppm, aplicat dupa germinarea semintelor, determina alungirea radacinii principale si stimuleaza formarea radacinilor adventive. Prin stropirea plantulelor de tomate cu sarea sodica a acidului 2,4 -D, in concentratie de 20 - 100 ppm, se intensifica culoarea verde a frunzelor, dar este franata alungirea internodurilor. Daca se folosesc 250 l/ha, din solutia mentionata , in concentratie de 20 ppm, se reduce de sase ori caderea bobocilor si a florilor din primele doua inflorescente , in comparatie cu martorul. Prin stropirea inflorescentelor de tomate cu solutii apoase de 2,4-D, in concentratie de 5-10 ppm, se obtin sporuri la productia de fructe cu 40-90% formarea de fructe partenocarpice si cu numar redus de seminte;se produce maturarea fructelor cu 5-12 zile mai devreme. Efectele similare se obtin si prin tratarea rasadurilor de tomate, inainte de plantare, cu solutii apoase de 2,4 -D , in concentratie de 15-20 ppm.
Tomatele cultivate in sera, datorita temperaturii ridicate si a luminozitatii reduse, prezinta o sterilitate accentuata la primele inflorescente. Daca florile acestor plante sunt tratate in fazele initiale, in special in perioada deschiderii florilor, cu solutii de acid 2,4,5 - triclorfenoxiacetic sau cu acid naftaxiacetic se obtin fructe fara seminte.
Prin tratarea mugurilor florali de Solanum melangena (vinete) cu solutii apoase de acid b-naftaxiacetic, in concentratie de 10-15 ppm creste procentul de legare a fructelor si a productiei de vinete cu 20-30% . Sporuri insemnate de productie de 35-40% se obtin si la Capsicum annuum (ardei) prin stropirea frunzelor si a inflorescentelor cu solutii apoase de acid naftalacetic, in concentratie de 200 ppm.
Prin utilizarea a 30 l la tona de tuberculi de cartofi , solutie apoasa de AIA, de 50 ppm, se intensifica hidroliza substantelor de rezerva (amidon, proteine, lipise etc), creste continutul de monoglucide si de aminoacizi, fapt ce stimuleaza formarea lastarilor , cresterea tulpinilor si sporirea productiei la hectar cu 15-18%. Prin tratarea cartofilor timpurii inainte de plantare, cu sarea de sodiu a acidului 2,4-D, in concentratie de 50 ppm, se obtine un spor de productie de 6340 kg/ha .
La Daucus carota (morcov) sporul productiei la radacini creste in medie cu 12% prin inmuirea semintelor inainte de semanat , in solutie de AIA 10 ppm, timp de 18 ore. Daca in solutia de AIA se adauga si unele microelemente (B, Cu, Co, Zn, etc) sporul de productie a radacinilor de morcov va fi si mai mare.
Prin tratarea butasilor de crizanteme cu 5-7 noduri, cu acid indolilacetic, sub forma de solutii apoase de 100 - 400 ppm, se realizeaza franarea formarii mugurilor florali, fapt ce duce la intarzierea infloririi crizantemelor.
Tratatamentele cu solutii apoase de AIA, in concentratii forte variate, cuprinse intre 10 si 200 ppm, realizate prin scufundarea bulbilor in solutie determinata stimularea cresterii si imbunatatirea calitatii florilor de gladiole.
Acidul indolilacetic in amestec cu acidul indolilbutric, dispersat in pudra de talc, in concentratie de 5000 ppm, stimuleaza inradacinarea butasilor de Ficus de aproape 100%.
Fitohormonii auxinei au numeroase utilizari practic in floricultura si dendrologie, atat asupra cresterii, inradacinarii cat si a infloririi a numeroase specii de arbusi si arbori fructiferi. Daca se introduce capatul inferior al butasilor in solutii de auxina, sau se acopera cu pudra de talc care contine auxina, se realizeaza o inradacinare mai bunp a plantelor tratate fata de martor. Apar un numar mai mare de radacini adventive intr-un timp relativ scurt.
Tratamentele
cu acid ![]() -naftilacetic 1% sub forma de sare sau de ester etilic,
efectuate la meri, determina franarea cresterii lastarilor
lacomi si obtinerea de lastari scurti, roditori,
imbracati cu numerosi muguri de rod. Prin stropirea pomilor
fructiferi, in special a merilor, cu auxine artificiale, in concentratie
de 100 g/ha , cu o lunp inainte de recoltare , se evita intr-o proportie
insemnata scaderea prematura a fructelor. Pentru meri se
folosesc stropiri cu acid
-naftilacetic 1% sub forma de sare sau de ester etilic,
efectuate la meri, determina franarea cresterii lastarilor
lacomi si obtinerea de lastari scurti, roditori,
imbracati cu numerosi muguri de rod. Prin stropirea pomilor
fructiferi, in special a merilor, cu auxine artificiale, in concentratie
de 100 g/ha , cu o lunp inainte de recoltare , se evita intr-o proportie
insemnata scaderea prematura a fructelor. Pentru meri se
folosesc stropiri cu acid ![]() -naftilacetic, iar pentru lamai si portocali acidul
2,4-D. numeroase fructe , mai cu seama portocalele si lamaile,
invelite in hartie imbibata cu auxine, se pastreaza mai bine
si au un colorit mai stabil. Grabirea maturarii fructelor la
caisi si piersici se realizeaza prin stropiri cu acidul
4-clorfenoxiacetic.
-naftilacetic, iar pentru lamai si portocali acidul
2,4-D. numeroase fructe , mai cu seama portocalele si lamaile,
invelite in hartie imbibata cu auxine, se pastreaza mai bine
si au un colorit mai stabil. Grabirea maturarii fructelor la
caisi si piersici se realizeaza prin stropiri cu acidul
4-clorfenoxiacetic.
Unele
auxine de sinteza (acidul 2,4,5 - triclorfenoxiacetic, acidul
2,4,5-triclorfenoxipropionic) se utilizeaza pentru a rari
numarul florilor, prin stimularea caderii acestora, cand florile sunt
in numar prea mare, asigurandu-se astfel o marire uniforma a
fructelor. Acidul ![]() -naftilacetic, in solutii concentrate este utilizat
pentru a impiedica incoltirea cartofilor, a bulbilor de ceapa,
rizomilor si pornirea mugurilor la materialul saditor.
-naftilacetic, in solutii concentrate este utilizat
pentru a impiedica incoltirea cartofilor, a bulbilor de ceapa,
rizomilor si pornirea mugurilor la materialul saditor.
Rarirea
fructelor de par se realizeaza prin stropirea pomilor cu solutii de acid ![]() -naftilacetic, in concentratie de 25 ppm. Tratarea
pomilor fructiferi cu preparate au inice, pentru prevenirea scaderii
fructelor si evitarea aparitiei unor efecte secundare nedorite, se
fac cu solutii de acid
-naftilacetic, in concentratie de 25 ppm. Tratarea
pomilor fructiferi cu preparate au inice, pentru prevenirea scaderii
fructelor si evitarea aparitiei unor efecte secundare nedorite, se
fac cu solutii de acid ![]() -naftilacetic, in concentratii de 10-20 ppm, iar cele cu
acid 2,4,5-triclorfenoxiacetic si 2,4,5-triclorfenoxipropionic cu
solutii avand concentratia intre 20 si 40 ppm.
-naftilacetic, in concentratii de 10-20 ppm, iar cele cu
acid 2,4,5-triclorfenoxiacetic si 2,4,5-triclorfenoxipropionic cu
solutii avand concentratia intre 20 si 40 ppm.
Tratamentele
cu fitoregulatori au efecte pozitive si in ce priveste calitatea strugurilor.
Daca se fac stropiri cu solutii de acid ![]() -naftilacetic in perioada maturarii strugurilor,
creste continutul de glucide, scade aciditatea acestora, iar raportul
dintre substantele solide/aciditate are valori mai mari. Se stie ca pentru
mentinerea caracteristicilor de soi, vita de vie se
inmulteste pe cale vegetativa, prin butasi de tulpina
si nu prin seminte. Stimularea inradacinarii
butasilor se poate realiza prin tratamente cu acid indolilacetic, in
concentratie de 5-30 ppm si cu acid
-naftilacetic in perioada maturarii strugurilor,
creste continutul de glucide, scade aciditatea acestora, iar raportul
dintre substantele solide/aciditate are valori mai mari. Se stie ca pentru
mentinerea caracteristicilor de soi, vita de vie se
inmulteste pe cale vegetativa, prin butasi de tulpina
si nu prin seminte. Stimularea inradacinarii
butasilor se poate realiza prin tratamente cu acid indolilacetic, in
concentratie de 5-30 ppm si cu acid ![]() -naftilacetic 5 ppm- aplicate timp de 24 ore, prin
scufundarea in solutie a partii inferioare a butasului.
-naftilacetic 5 ppm- aplicate timp de 24 ore, prin
scufundarea in solutie a partii inferioare a butasului.
Auxinele de sinteza , in concentratie mai mare (10-1 - 10-3M/l) sunt utilizate ca erbicide pentru distrugerea buruienilor de dicotiledonate din culturile de graminee. O mare parte din buruieni sunt distruse cu acid 2,4-diclorfenoxiacetic, utilizand 1-1,5 kg substanta activa la hectar. Prin utilizarea unor doze mai mici de 2,4-D buruienile nu sunt distruse, dar sunt franate in crestere, astfel incat cultura principala va umbru buruienile.
2.4. CITOKININE
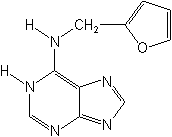
Citokininele sunt fitohormoni care deriva de la adenina. Ei au un rol foarte important in stimularea diviziunii celulelor vegetale mature nemeristematice.
Cresterea si dezvoltarea plantelor este influentata de fitohormoni si de numerosi alti compusi fitoregulatori, care stimuleaza sau inhiba cresterea si dezvoltarea organismelor vegetale.
Descoperirea definitiva a noului compus endogen, care induce puternic diviziunea celulelor s-a realizat in 1955 de catre C.O. MILLER, F.SKOOG, M.H.VON SALTZA si F.M. STRONG, in laboratoarele Universitatii Wisconsin, S.U.A., iar dupa efectul sau specific asupra citokinezei i s-a atribuit de kinetina. Sub aspectul chimic kinetina este 6-flurfuriladenin (sau 6-furfurilaminopurina), cu formula moleculara C10H9N5O.
Dupa extragerea si identificarea kinetinei s-au descoperit si alte substante naturale, cu structura si efecte similare acesteia, stabilindu-se ulterior existenta unei clase de compusi naturali cu efecte asupra citokininei, atribuindu-li-se denumirea generala de kinine.
Kinetina este un derivat al adeninei care este o baza purinica importanta, raspandita in natura, ce se gaseste atat in stare libera, cat mai cu seama sub forma de compusi (acizi nucleici, coenzime, compusi macroergici etc.).
Prin denumirea de kinetina se intelege 6-furfuriladenina sau 6-furfurilaminopurina. Restul compusilor chimici care stimuleaza diviziunea celulelor (cytokinesis) si contribuie la reglarea cresterii si dezvoltarii plantelor.
Rolul si functii fithormonale
Citokininele sunt fitohormoni care au in organismele vegetale un rol multiplu si complex. Ele au numeroase functii prin care influenteaza crestereasi dezvoltarea plantelor, luand parte in toate fazele importante ale acestor procese; de la germinare, la maturare, imbatranire si moartea plantelor. Reglarea proceselor metabolice de crestere si de dezvoltare a organismelor vegetale se realizeaza prin influenta ce o au citokininele asupra metabolismului acizilor nucleici, a biosintezei proteinelor, influentei asupra activitatii enzimelor, cat si prin translocatia si acumularea compusilor anorganici si a celor organici rezultati in urma proceselor de biosinteza.
Citokininele libere prezinta activitate biochimica si fiziologica independenta de orice asociere a lor cu sARN sau cu alti compusi.
Activitatea fitohormonala de baza a citokininelor consta in stimularea biosintezei acizilor nucleici (sARN , mARN) si a proteinelor, activarea sau inhibarea activitatii unor enzime, procese care inhiba imbatranirea celulelor si a tesuturilor vegetale.
In ce priveste mecanismul de actiune a citokininelor s-a stabilit existenta unei mari asemanari cu mecanismul de actiune a hormonilor in organismele animale, prin stabilirea unor legaturi intre hormoni si receptori intracelulari. In cazul actiunii citokininelor receptorul lor este o proteina allosterica, cu care citokininele stabilesc legaturi de hidrogen sau legaturi ionice. Ca urmare a stabilirii acestor legaturi, receptorul poate sa formeze un complex receptor - hormon, care sa stimuleze activitatea hormonului sau poate receptorul sa-si modifice conformatia , sa influenteze activitatea hormonala.
Aplicatii practice ale citokininelor
Aplicatii practice ale fitoregulatorilor, inclusiv ale citokininelor, sunt mai pe larg prezentate, in ultimul deceniu, in literatura romana.
Citokininele naturale , cat mai cu seama cele de sinteza, au numeroase aplicatii practice in cultura cerealelor, in legumicultura, pomicultura, floricultura, viticultura, silvicultura, in cultura plantelor tehnice etc. Prin aplicarea unor tratamente foliare cu solutii diluate de citokinine la grau, orz si secara, in diferite stadii de dezvoltare, creste randamentul fotosintezei, continutul clorofilelor, a azotului si a substantei uscate; creste de asemenea rezistenta plantelor la stresul salin, la rugina si la intarzierea senescentei frunzelor.
Prin tratarea semintelor in repaus, de orez rosu cu solutii 10-3 M de kinetina, benziladenina sau izopenteniladenina, la 300C si pH acid, se produce o germinare a acestor seminte in proportie de 80-90%.
Kinetina in solutie apoasa, in concentratie de 10-4M, aplicata pe frunze de mazare inainte de inflorire mareste substantial volumul pastailor si al boabelor, favorizeaza acumularea de azot si fosfor in boabe, formarea nodozitatilor, contribuind atat la sporirea productiei de mazare cat si la imbunatatirea calitatii acesteia. Daca plantele de Phaseolus radiatus sunt tratate cu solutii de kinetina, in concentratie de 50-100 mg/l , se produce o inhibare a cresterii radacinilor si a tuplinii, datorita probabil inhibarii biosintezei substantelor proteice. Prin tratarea frunzelor de soia, dupa inflorirea plantelor, cu o solutie de benziladenina, in concentratie de 2 x 10-3 M, se produce o crestere remarcabila a productiei de boabe, in comparatie cu lotul martor.
Citokininele sunt capabile sa intrerupa starea de latenta a tuberculilor de cartofi si a legumelor bulboase. Prin tratarea soiului de cartof Majestic cu solutii de kinetina si zeatina , in concentratii de 10 - 100 ppm, timp de 6-24 ore, se realizeaza pornirea mugurilor dupa doua zile de la imceperea tratamentelor. Cresterea productiei de cartofi se poate realiza prin tratarea acestora cu kinetina si benziladenina, in concentratie de 2,5 -25 ppm, realizandu-se totodata o marime apropiata, mijlocie , a tuberculilor de cartofi.
Tratamentele cu citokinine (benziladenina, kinetina, 6-fenilaminopurina, zeatina, etc), in diferite concentratii , mentin o perioada indelungata ridichile in stare proaspata, se asigura un grad ridicat de turgescenta si fragezime, reducandu-se totodata procesele de degradare a clorofilelor si a carotenoidelor. Pentru pastrarea proaspata a ridichilor, sfeclei, morcovilor etc, tratamentele cu citokinine sunt mai eficiente daca se efectueaza impreuna cu unii retardanti (Alar, CCC, Fosfor-D).
Citokininele in solutii diluate, de 1 ppm, prin tratamente de scurta durata determina cresterea plantelor de ceapa, usturoi, praz etc. , datorita stimularii diviziunii celulare, accelerarii biosintezei acizilor nucleici, cresterea permeabilitatii membranelor plasmatice celulare.
Citokininele au largi aplicatii practice si la legumele frunzoase si varzoase, atat pentru sporirea productiei, imbunatatirea calitatii acestora cat si pentru pastrarea prospetimii partilor frunzoase pe o perioada indelungata , dupa recoltare. Tratamentele cu citokinine s-au dovedit extrem de eficiente, pentru pastrarea prospetimii si a culorii frunzelor la salata, spanac, marar, patrunjel, telina, varza, varza de Bruxelles, conopida, sparanghel etc, fapt ce asigura consumul acestor legume in stare proaspata o perioada indelungata. Tratamentele se efectueaza de obicei cu citokinine sintetice, in concentratie de 5-10 ppm, prin pulverizarea masei aeriene a plantelor cu cateva zile inaninte de recoltare sau imediat dupa recoltare. La varza, conopida si salata tratamentele efectuate dupa recoltare se realizeaza prin scufundarea capatanilor in solutie de benziladenina, in concentratie de 10 - 30 ppm. Daca plantele sunt mentinute la temperaturi scazute, dupa 20 de zile dupa aplicarea tratamentului continutul clorofilelor este de 2- ori mai mare decat la lotul martor, netratat cu benziladenina. Cresterea continutului de clorofila totala la varza, prin tratare cu kinetina sintetica se prezinta in tab.1.
Tab.1.
Continutul clorofilei totale la varza sub influenta tratamentelor cu benziladenina
|
Benziladenina ppm |
Continut de clorofila totala mg/g sus. proaspata |
La varza de Bruxelles prospetimea verzisoarelor se realizeaza prin scufundarea acestora timp de 1 minut in solutii apoase de benziladenina, in concentratie de 10 ppm , in amestec cu solutii de acid naftalenacetic. De25 ppm. Franarea imbatranirii si a ingalbenirii frunzelor se realizeaza la numeroase plante prin efectuarea unor tratamente mixte cu citokinine si auxine sintetice, in concentratie de 20 - 25 ppm. Franarea imbatranirii frunzelor, sub actiunea tratamentelor cu citokinine, se realizeaza atit prin stimularea biosintezei acizilor nucleici, a proteinelor, clorofilelor, carotenoidelor etc., cat si prin reducerea activitatii respiratorii, franarea degradarii enzimatice a clorofilelor, a proteinelor etc. Prin utilizarea de 14C s-a remarcat ca in frunzele de vara (Brassica oleracea var.italica), in urma tratamentelor cu kinetina creste continutul acidului orotic ca precursor l bazelor piramidinice din molecula acizilor nucleici si a leucinii, care intra in constitutia proteinelor nou sintetizate.
In tabelul 2. se prezinta continutul 14C-acidului orotic incorporat in ARN si a 14C-leucinei incorporata in proteine.
Tabel.2.
|
Tratament |
C -acid azotic |
C -leucina |
|||
|
Absorbtie neta |
Incorporare |
Absorbtie neta |
Incorporare in proteine |
||
|
Lot de control - 3 ore | |||||
|
Kinetina 3 ore | |||||
|
Lot de control 6 ore | |||||
|
Kinetina 6 ore | |||||
Semintele de salata, care la umezeala si la 350C nu germineaza de loc, daca sunt tratate cu solutii de citokinine (kinetina, benzilaminopurina) in concentratie de 100 ppm, timp de 3 minute, germineaza im proportie de aproximativ 67%.
Prin tratamente fizice (ultrasunete, temepraturi joase, presiune scazuta, lumina intensa etc) asemenea seminte germineaza in proportie mica, deaproximativ 30%.
M.L. GORLOVA (1969) a cercetat actiunea stimulatoare a citokininelor asupra germinarii semintelor la numeroase plante (marar, patrunjel, telina, spanac, morcovi, varza, conopida, gutui etc). Tratamentele cu citokinine la aceste plante nu numai ca au sporit procentul de germinatie a semintelor , dar s-a produs si o reducere a perioadei de vegetatie cu 4-7 zile, a crescut productie cu 5-19%, a srescut continutul vitaminei C, a glucidelor solubile, azotului, substantei uscate etc. Cresterea fitomasei plantelor se realizeaza prin tratarea frunzelor cu kinetina sau benziladenina, in concentratie de 12-50 ppm.
Citokininele au numeroase si variate aplicatii practice in floricultura. Despre rolul si mai cu seama frumusetea florilor C.I. MILICA si colab. 1983 mentioneaza ca ,,din toate timpurile florile au intuchipat frumusetea, perfectiunea creatiei, armonia si linistea sufleteasca."
Benziladenina prelungeste timpul de pastrare a garoafelor taiate cu 3-5 zile fata de proba martor. Garoafele vor fi scufundate in solutii de bnziladenina de diferite concentratii si mentinute submers un timp invers-proportional cu concentratia solutiei. Astfel in solutie de concentratia 10-3M vor fi mentinute doar doua minute, in solutie de concentratie 10-4M vor putea fi mentinute 50 de minute, iar daca solutia este foarte diluata durata tratamentului poate fi de 12 ore.
La trandafiri, care se inmultesc prin butasi, citokininele stimuleaza formarea radacinilor adventive, stimuand astfel cresterea plantelor.
In pomicultura, la arborii fructiferi continutul citokininelor din lastari are valori ridicate in timoul infloririi depline si scade la valori minime in perioada verii cand inceteaza cresterea lastarilor. Mugurii terminali datorita faptului ca au o vascularitate mai buna primesc o cantitate mai mare de citokinine, fapt ce va determina formarea de primordii florale, iar mugurii laterali vor ramane vegetativi. Daca in seva xilemului continutul de citokinine are valori corespunzatoare concentratiei moderate, o mare parte din mugurii axilari devin floriferi. De altfel diferentierea mugurilor florali de pe lastarii lungi ai pomilor (mar, par, prun, cires etc.) depinde de raportul dintre continutul citokininelor si al giberelinelor, iar de pe formatiile scurte, de balanta dintre gibereline si auxine. La arborii fructiferi tratamentele cu citokinine determina iesirea din repaus a muguilor , substituind partial nevoia de frig a pomilor. Intreruperea repausului la mugurii de mar si piersic se realizeaza eficient prin tratarea acestora cu solutii de benzilaminopurina, in concentratie de 100 - 2000 ppm.
La vita de vie continutul maxim al citokininelor din lastari se obtine in perioada cresterii boabelor de struguri , iar frunze , in timpul maturarii boabelor. In inflorescente continutul citokininelor are valori ridicate in perioada infloririi si a maturarii boabelor. In cadrul boabelor mature continutul citokininelor in ordine descrescanda se prezinta in coji , pulpa si seminte. Si la vita de vie tratamentele cu citokinine stimuleaza inflorirea, precum si conversia sexelor, influentand tranformarea florilor hermafrodite in flori femele, iar a florilor mascule in flori hermafrodite.
2.4. ETILEN
Etilenul este primul fitohormon gazos cunoscut pana in prezent, cu un larg spectru de actiune biochimica si fiziologica asupra proceselor de crestere si dezvoltare ale plantelor.
Sub aspectul chimic etilenul este o hidrocarbura aciclica nesaturata, cu doi atomi de carbon (CH2 = CH2), fiind primul component al seriei alchenelor. Este o substanta gazoasa, incolora, inodora, foarte putin solubila in apa, usor solubila in numerosi solventi organici. Are masa moleculara 28,03 p.t. - 169,4 0C (ALBU si colab, 1974).
In natura etilenul, desi are o concentratie mica, are o raspandire relativ mare intrucat este produs de aproape toate organismele vegetale si animale, fiind un compus metabolic endogen.
Rolul si functiile etilenului
Pe baza a numeroase si variate date experimentale s-a stabilit ca etilenul are in organismele vegetale un rol multiplu si complex. In functie de concentratia sa, de relatia doza - raspuns, etilenul determina actiuni de stimulare sau de inhibare a diferitelor procese biochimice si fiziologice din plante. Prezinta de asemenea interrelatii sinergice sau antagonice cu ceilalti fitohormoni, participand activ la reglarea endogena a cresterii si dezvoltarii plantelor. Dintre principalele functii ale etilenului, care cintribuie la reglarea unor procese biochimice si fiziologice la plante se pot mentiona coacerea fructelor, senescenta (imbatranira) , abscizia frunzelor si a fructelor, epinastia frunzelor, strangerea carceilor, elonagrarea, hipertrofia, expansiunea frunzelor, inductia inflorescentei , expresia sexului, exudatia, inhibarea cresterii si ingrosarea radacinilor si a tuplinilor, inhibarea fiviziunii celulelor si a biosintezei acizilor nucleici etc.
Concentratia de etilen necesara pentru a produce efecte de pornire intr-o varietate de raspunsui fiziologice este de 0,01 ppm. Raspunsuri maxime apar la o concentratie optima de aproximativ 0,1 ppm; iar orprirea raspunsurilor (saturatia) apare la o concentratie maxima de 10 ppm.
Nivelul minim de pornire, de actiune a etilenului depinde de natura plantei, de organele si tesuturile asupra carora actioneaza , de starea fizilogica , fenofaza etc.
In comparatie cu alte gaze fiziologic active, etilenul este de multe ori mai eficient in producerea raspunsurilor vizibile la plante. Astfel, concentratia minima a etilenului, efectiva in producerea inclinarii tulpinilor de mazare si a epinastiei petiolurilor de tomate este de 0,2 ppm, a acetilenei este de 250 ppm, a propilenei de 1000 ppm, a monoxidului de carbon de 5000 , in timp ce butilena este aproape inactiva. Etilena aflat in atmosfera in concentratie mai mare, prin activitatea fiziologica intensa si gama larga de efecte pe care le are poate cauza pagube mari la cereale, fructe, legume , struguri etc.
Coacerea fructelor
Etilenul este considerat principalul fitohormon care declanseaza coacerea si maturitatea fructelor la numeroase specii de plante (tomate, mar, par, vita de vie), usurand totodata desprinderea fructelor de pe plante . etilenul impreuna cu alti fitohormoni cu actiune similara sau contrara cu a sa determina cresterea respinratiei, ingalbenirea pigmentilor, modificarea permeabilitatii membranelor protoplasmatice, biosinteza unor enzime , inmuierea cojii etc.
In procesul de coacere a fructelor oxigenul stimuleaza actiunea etilenului, in timp ce CO2 se comporta ca un inhibitor. Pentru pastrarea fructelor un timp mai indelungat este necesar ca aceastea sa fie depozitate in camere cu atmosfera mai bogata in CO2 (5-10%), scazuta in O2 (1-3( si foarte redusa in etilen. Depozitarea fructelor este bine sa se faca in camere aerisite, la temperaturi scazute, prevazute cu filtre de carbune brominat, pentru fixarea etilenului produs de fructe . Continutul oxigenului va scadea datorita respiratiei materialului vegetal, iar continutul CO2 va creste, fapt ce va influenta negativ biosinteza etilenului.
In timpul coacerii fructelor se produc schimbari in intensitatea respiratiei , asociata cu biosinteza unor proteine enzimatice, hidroliza materialelor intercelulare care contribuie la cimentarea tesuturilor vegetale, inmuierea pulpei fructelor, schimbari in culoarea , aroma si parfumul fructelor. In timpul coacerii fructelor se intensifica procesele de glicoliza. Creste de 10 ori continutul esterului fructozo-1,6-difosforic si se intensifica de 5 ori activitatea fosfofructokinazei si a piruvatkinazei. Concomitent se produce o scadere a continutului esterului fructozo-6-fosforic si a acidului enolpiruvic. Coacerea fructelor poate fi privata ca un caz special al senescentei organelor. Fructele carnoase sunt mult mai complexe in procesul de senescenta decat alte organe. La unele specii de plante se produce o scadere a intensitatii respiratiei in fructele mature, dar urmeaza o crestere semnificativa a respiratiei in timpul coacerii (climactericul respirator), dupa care din nou se produce o scadere a respiratiei in perioada de imbatranire a fructelor. Dintre fructele cu climacteric respirator fac parte marul, parul, bananele, avocado, mango etc.
Abscizia frunzelor
Un alt efect important al etilenului endogen si a celui exogen consta in determinarea caderii fructelor, florilor, frunzelor prin grabirea absciziei pedunculilor foliari, florali si a coditei fructelor. Caderea frunzelor, sub actiunea etilenului, a fost remarcata in 1917 de catre S.L. DOUBT.
Abscizia frunzelor se datoreste in mare parte reducerii transportului de fitohormoni (auxine, gibereline, citokinine) de la limbul foliar spre peduncul, precum si cresterii continutului si activitatii unor hidrolaze ,,localizate in membranele celulare din zona de desprindere a petiolului". Scaderea continutului fitohormonilor de crestere mentionati duce la o imbatranire a celulelor din petiol. Celulele specializate in abscizia frunzelor si a fructelor, sub influenta etilenului endogen sau exogen, produc hidrolaze care distrug peretele celular. Dintre aceste enzime hidrolitice cea mai specifica si mai importanta este celulaza. Actiunea etilenului in abscizia frunzelor la fasole se produce in doua faze. In prima faza se accelereaza maturitatea celulelor din zona de abscizie, iar in faza a doua se intensifica biosinteza celulazei si a altor β-glicozidaze (pectinaze, tanaze etc.) care slabesc peretele celular- Faza a doua este mai scurta si are rolul hotarator in producerea absciziei. La tutun , abscizia peduncului floral se produce la 9 ore de la aplicarea etilenului exogen. Efectele etilenului pot fi inhibate prin tratamente cu auxina, intrucat etilenul are o actiune reversibila. Caderea frunzelor se poate evita aproape integral prin stropirea plantelor cu acid naftalenacetic in concentratie de 250 ppm.
Inhibarea alungirii radacinilor si a tulpinilor
Etilenul actioneaza ca un inhibitor al alungirii radacinilor si a tulpinilor, iar in unele cazuri ca stimulator al dezvoltarii laterale, fapt ce determina ingrosarea organelor tratate cu aceste fitohormon. Etilenul inhiba de asemenea dezvoltarea mugurilor si a lujerilor. Tratamentele cu etilen exogen la bob, mustar, ridichi, rosii, grau, soia, mazare etc. Inhiba alungirea radacinilor si stimuleaza ingrosarea si strambarea acestora.
Prin tratarea tesuturilor mersitemice din radacini cu etilen se produce o reducere semnificativa a diviziunii celulare si a biosintezei de ADN. La mazare, aceste efecte negative , pot fi indepartate prin tratarea plantelor cu solutii netoxice care contin ioni de Ag+. Inhibarea cresterii radacinilor sub actiunea etilenului se produce si la plantele cultivate ,,in vitro".
Tratamentele cu etilen exogen inhiba cresterea frunzelor, stimuleaza biosinteza acidului abscizic, reduce activitatea axinelor, giberelinelor si a citokininelor.
Pentru stimularea cresterii radacinilor doza de etilen trebuie sa fie la tomate sub 0,02 ppm, la mazare sub 0,15 ppm, la orez sub 1 ppm, la laptuca sub 0,1 ppm, La un continut mai mare a etilenului fata de aceste valori se produce o inhibare ireversibila a cresterii radacinilor.
Etilenul are un rol important si in protectia varfurilor sensibil al lastarilor de invatamare in timpul germinarii si a rasaririi, atat la plantele cu germinatie hipogena (Pisum sativum), la care cotiledonul ramane sub pamant cat si la cele cu germinatie epigena (Phaseolus vulgaris), la care hipocotilul elongheaza, formand un carling sau arc, varful lastarului fiind astfel protejat in timp ce este impins spre suprafata. In ambele cazuri s-a remarcat o crestere simetrica in lastar, ceea ce cauzeaza indepartarea carligului epicotilului sau hipocotilului, dupa ce rasadul ajunge la suprafata pamantului. Productia de etilen in cazul lastarului de mazare etiolat este de 61/kg-1 ora-1. aceasta cantitate de etilen este suficienta pentru formarea si mentinerea carligului in rasadurl etiolat. Dupa ce rasadul etiolat rasare, natural din sol, creste continutul de etilen in regiunea carligului si se stabilizeaza cresterea simetrica a lastarului. Dupa inverzirea etiolat acesta devine din ce in ce mai sensibil la actiunea etilenului produs.
Determinarea sexului
Etilenul determina la unele plante, in special la cele din familia Cucurbitaceae (castravete, dovleac, pepene etc.) o crestere a florilor femele, concomitent cu scaderea florilor mascule. Tratamentele cu etilen, exogen, stimuleaza de asemenea formarea florilor la plantele din familia Bromeliaceae. La alte familii si genuri de plante (Perilla, Xanthium, Chrysanthemum), tratamentele cu etilen inhiba complet formare florilor.
De mare importanta practica este utilizarea etilenului pentru stimularea infloririi la mango si la ananas.
Alte efecte ale etilenului
Etilenul in concentratii mai mari determina o crestere (ingrpsare) anormala a tlpinilor si a radacinilor, stimuland formarea radacinilor adventive. Expansiunea radiala a celulelor, care in mod normal se extind prin alungire, este corelata cu alterarea de catre etilen a orientarii fibrelor de celuloza in peretele celular.
Asa dupa cum tesuturile fructelor raspund la etilen producand enzime caracteristice coacerii si absciziei, o mare varietate de tesuturi raspund la actiunea etilenului prin biosinteza unor enzime caracteristice. In cateva tesuturi inhibitori i biosintezei ARN blocheaza si actiunea etilenului.
La mazare toate partile rasadului raspund la actiunea etilenului. Cresterea tulpinii si a radacinii incetineste, se strang carceii, subapexul se mareste (bombeaza) si se inclina orizontal, se mareste zona de elongare a radacinii, se formeaza peri radiculari etc. Dupa 3-4 ore de la aplicarea etilenului se produce dezvoltarea subapicala care necesita biosinteza de proteine. Se produce o scadere substantiala in incorporarea peptidelor din peretele celular care contin hidroxiprolina. Colchina si vinblasina determina marirea subapexului si inhiba depunerea peptidelor cu hidroxiprolina in peretii celulari.Celulele subapicale se maresc si sub influenta kinetinei, benzimidazolului, benzadeninei etc.
Etilenul influenteaza comportamentul tropistic si epinastic al plantelor. El determina strangerea carceilor la rasadurile etiolate, epinastia frunzelor, inclinarea orizontala a tulpinii, plagiotropismul la radacini etc. Toate aceste efecte se datores in mare parte inhibarii de catre etilen a transportului lateral de auxine.
La cartof , dupa un anumit timp de la aplicarea tratamentului cu etilen se produce o crestere a continutului si a activitatii peroxidazei, fenoloxidazei, fenilalanindezaminazei in radacini.
2.5. GIBERELINE
Giberelinele sunt fitohormoni naturali, derivati izoprenici, larg raspanditi in natura, care stimuleaza diviziunea si elongatia celulelor vegetale. Au efecte importante, adesea spectaculoase, asupra cresterii si dezvoltarii plantelor, fapt pentru care au numeroase aplicatii practice in ameliorarea plantelor, in obtinerea unor noi linii , hibrizi si soiuri de plante, putand fi utilizati pentru dezvoltarea intensiva a agriculturii , horticulturii, silviculturii, a plantelor medicinale etc.
Prescurtarea denumirii clasei giberelinelor se face pe plan international prin notarea (simbolul) GA, iar denumirea prescurtat[ a diferitelor gibereline cu GA1, GA2, GA3..,GA60. Dintre giberelinele descoperite pana in prezent acidul giberelinic, notat cu GA3, este principalul fitohormon din aceasta grupa, fiind cel mai raspandit, cu mare activitate biologica.
Structura chimica si clasificare
Sub aspectul structurii chimice giberelinele sunt derivati diterpenici care au ca structura comuna de baza un unel giberelinic (nucleu fluorenic), pe care sunt grefate diferite grupari functionale hidroxilice (-OH), carboxilice (-COOH), metilice (-CH3), metenilice (=CH2), aldechidice , cetonice (C = O), lactonice etc. Proprietatile diferitelor gibereline depind in mare aarte de felul , numarul , de configuratia si conformatia gruparilor functionale, precum si de forma sub care se gasesc in diferitele tesuturi si organe ale plantelor.
Rol, functii si utilizari
Fitoreceptori ti ciclul acidului adenilic (cAMP).
Giberelinele au un rol important in reglarea procesului de crestere si dezvoltare a plantelor, au numeroase functii si sunt tot mai mult utilizate cu succes in practica agricola si horticola.
Pentru ca celulele si tesuturile organelor vegetale sa reactioneze la actiunea giberelinelor endogene, sau a celor exogene, trebuie sa existe un receptor hormonal, in cazul tuturor hormonilor in organisme vegetale. Actiunea hormonilor nu se poate manifesta (celulele, tesuturile, organele si planta intreaga nu raspund la actiunea hormonului) pana ce hormonul nu este ,,recunoscut" de receptorul sau. Receptorii hormonali au o mare sensibilitate si specificitate in recunoasterea hormonilor, necesitand cantitati foarte mici de hormoni. Ei sunt capabili sa distinga deosebiri mici in ce priveste structura moleculara si activitatea hormonilor. Dintre substantele existente in celulele proteinele sunt cele mai potrivite pentru recunoasterea hormonilor si a activitatii lor specifice. Cercetarile pentru descoperirea unor receptori hormonali proteici sunt de mare actualitate si importanta pentru cunoasterea mecanismului si a functiunilor fitohormonilor.
Stimularea proceselor de crestere
Stimularea proceselor de crestere prin diviziunea si alungirea celulelor constituie cea mai evidenta actiune fiziologica, care se remarca prin alungirea tulpinilor, cresterea laterala a frunzelor si prin schimbarea habitusului general al plantelor.
Giberelinele sunt fitoregulatori de crestere care stimuleaza atat elongatiacelulelor , deci procesele de auxeaza, cat si diviziunea celulei, deci procesele de mereza, fie ambele procese la aceeasi specie. In cazul rasadurilor de fasole ambele procese au fost stimulate prin aplicarea giberelinelor exogene, dar efectul marcat s-a observat la alungirea celulelor. In schimb , efectul major sub actiunea giberelinelor consta la mazarea pitica, in meristemele aplicale la Hyosciamus niger si Samolus parviflorus in diviziunea celulelor. Pe baza mai multor date experimentale s-a observat ca diviziunea si alungirea celulelor depind de varsta si faza de dezvoltare a plantelor, de soi si numarul tratamentelor, de evolutia conditiilor meteorologice. Celulele tinere raspund in general la actiunea giberelinelor prin diviziune iar cele mai batrane prin alungire. Diviziunea celulra singura nu influenteaza semnificativ cresterea plantelor. Acest proces trebuie sa fie corelat cu o elongare normala sau intensificata a celulelor, pentru a remarca cresteri vizibile la plante.
In procesele de auxeaza (alungire celulara) giberelinele actioneaza indirect asupra celulelor, prin stimularea biosintezei triptofanului, care este precursorul principal al auxinei si prin inhibarea actiunii auxinoxidozelor care degradeaza fitohormonii auxinici. Giberelinele activeaza, de asemenea, biosinteza difenolilor, care se comporta ca inhibitori intracelulari ai ai auxinoxidazelor. Numerosi cercetatori sustin ca alungirea celulelor sub actiunea giberelinelor, se datoreste in mare parte actiunii auxinelor.
In procesele de mereza (diviziunea celulara) giberelinele actioneaza tot indirect, prin stimularea biosintezei ARN si ADN, care determina biosinteza unor proteine functionale (enzime) si de structura. Giberelinele exogene nu au nici o actiune asupra radacinilor.
Sub actiunea giberelinelor sunt afectate toate tesuturile si organele de crestere ale plantelor, internodurile si lastarii se alungesc de 3-4 ori mai mult decat probele martor. Se alungeste hipocotilul si epicotilul, celulele epidermice, perenchimatice si meristemice, se mareste volumul celular prin extensie. Giberelinele actioneaza asupra alungirii celulelor la concentratii mici (pana la 200 ppm) si intr-un timp scurt. La concentratii mari giberelinele la fel ca si alti fitohormoni devin inhibitori, dar nu si toxici.
Sub actiunea giberelinelor, limbul frunzelor la monocotiledoane creste mai mult in lungime, iar la dicotiledoane creste semnificativ si in latime , dubland uneori suprafetei frunzei. Frunzele tratate cu gibereline au o culoare mai deschisa datorita reducerii continutului de clorofile si de xantofile, pana la refacerea acestor pigmenti sub actiunea elementelor minerale.
Cresterea plantelor pitice de mazare, fasole, porumb etc. Este mult stimulata sub actiunea giberelinelor. La mazare (Pisum sativum), culturile pitice ajung la maturitate la o inaltime de 30 cm si chiar mai putin, iar plantele normale sunt inalte, de peste 1 m.
Giberelinele influenteaza fenomenul de dominanta apicala, care consta in inhibarea mugurilor apicali, pe cale fitohormonala , de catre mugurele terminal din varful tulpinii. Mugurele terminal produce auxina, care stimuleaza biosinteza de etilen in mugurii axialu, fapt ce inhiba cresterea acestora. Prin aplicarea giberelinelor exogene, acestia pornesc in vegetatie, dand numeroase ramificatii laterale plantelor tratate.
Giberelinele stimuleaza germinatia semintelor si prin accelerarea proceselor de stratificare a semintelor la buruieni, plante spontane, la unele plante lemnoase etc, scurtand perioada necesara de pastrare a acestora inainte de germinare. De asemenea giberelinele substituie efectele luminii la plantele fotoblastice care nu germineaza la intuneric. Semintele unor soiuri de salata nu germineaza decat prin expunerea acestora in stare umeda cateva ore la lumina. Giberelinele insa inlocuiesc aceasta necesitate a semintelor de salata.
Iesirea din repaus mugurilor plantelor lemnoase se face sub actiunea temperaturilor joase si uneori sub actiunea zileleor lungi. Arborii tratati cu gibereline nu intra in repaus, sau intra mult mai tarziu.
Se
poate afirma ca giberelinele anuleaza multe tipuri de inhibari
ale cresterii. ![]()
Stimularea infloririi, fructificarii si a altor procese fiziologice ale plantelor.
Plantele tratate cu gibereline infloresc si fructifica mai devreme decat plantele de control. Stimularea infloririi plantelor sub actiunea giberelinelor s-a remarcat la mazare, morcovi, fasole, tomate, varza, telina , cereale paioase, porumb etc. Giberelinele determina grabirea formarii florilor, marimea acestora, germinarea polenului, influenteaza raportul dintre florile mascule si cele femele etc. GA3 stimuleaza la plantele din familia Cucurbitaceae formare florilor femele si reducerea numarului celor mascule.
Prin stropirea cu solutii diluate de gibereline se determina inflorirea plantelor bianuale (sfecla, morcov, varza, nap, patrunjel, maselatita etc.), in primul an de la recoltare, fara ca aceste plante sa fie supuse actiunii temperaturilor joase. De asemenea pot determina inflorirea plantelor de zi lunga, in conditii de iluminare redusa, caracteristica plantelor de zi scurta. Giberelinele anuleaza de asemenea efectul de franare a cresterii sub actiunea luminii rosii. La lolium perene si phleum pratense s-a remarcat un continut de gibereline la inceputul formarii inflorescentei, dupa care cantitatea de gibereline scade treptat cu maturarea florilor.
Tratamentele cu gibereline stimuleaza formarea fructelor partenocarpice, cu mare importanta economica. La tomate creste frecventa partenocarpiei de 500 ori. La mar, par, struguri tratamentele cu gibereline constituie singura cale eficienta de obtinere a fructelor partenocarpice.
Continutul giberelinelor din fructe si seminte precum si din alte organe corespunde cu perioadele de crestere intensiva a fructelor, semintele , florilor, tulpinilor, mugurilor etc. Continutul giberelinelor este mai mare in boabe strugurilor cu seminte decat in boabele celor fara seminte, indicand ca semintele strugurilor sunt surse bogate in gibereline. De asemenea sub actiunea giberelinelor alungirea celulelor din strugurii ce contin boabe fara seminte este cu mult mai mare decat in cazul strugurilor ce contin boabe cu seminte. Nu se stie precis daca giberelinele din seminte contribuie la cresterea fructelor carnoase. Scaderea giberelinelor endogene influenteaza favorabil coacerea fructelor carnoase.
Prin tratamente cu gibereline se pot obtine boabe de struguri fara seminte destinate consumului. Se pot determina cresteri anuale mari la puietii de arbori si arbusti. Se poate provoca germinarea semintelor de buruieni, care sunt in repaus, realizandu-se astfel distrugerea lor mai usoara, fara cheltuieli economice prea mari. Prin stropiri cu solutii diluate de giberelina se poate stimula cresterea gazonului in perioadele de toamna si primavara timpurie, cand temperaturile sunt mai scazute. Tot cu gibereline se poate provoca ramificarea crizantelelor si a altor plante, grabirea infloririi la plantele dependente de temperaturi mai joase si de fotoperioade lungi.
Prin tratarea tomatelor, la varsta plantelor de 14-25 zile, cu solutii de acid giberelic in concentratie de 100 ppm se asigura cresterea masei aeriene (tulpini, frunze, muguri , flori).
Prin stropirea plantelor cu solutii apoase de gibereline se grabeste inflorirea legumelor cu 4-5 zile fata de probele martor, iar florile sunt mai mari si mai larg deschise. Tratarea plantelor cu GA3, inainte de inflorire stimuleaza legarea si cresterea fructelor cu peste 50%.
Prin scufundarea tuberculilor de cartofi, timp de 3-10 min. Intr-o solutie de GA3 (1-8 ppm), in care se poate introduce si fungicide pentru combaterea ciupercilor, se intrerupe repausul obligatoriu , indiferent de perioada fizilogica in care se gasesc tuberculii, producandu-se totodata incoltirea cartofilor. Pentru plantarea de vara a cartofilor, imediat dupa recoltarea culturii anterioare, tuberculii de cartofi vor fi imbaiati in solutii de gibereline de concentratie de 15 ppm timp de 30 minute.
Stropirea plantelor cu solutii de gibereline, inaintea formarii florilor va permite utilizarea acestor hormoni pentru substituirea castrarii manuale la hibridare, la crearea unor noi linii valoroase.
Prin tratarea repetata a plantelor de conopida cu GA3, in concentratii de 50 ppm, la intervale de 5-7 zile, in perioada cand inflorescentele aveau un diametru de 2-5 cm, s-a produs o crestere semnificativa in productia de inflorescente, a crescut, de asemenea, continutul substantei uscate.
Calitatea si cantitatea productiei de telina, patrunjel, morcov, gulii, varza, spanac, porumb, cereale paioase, sfecla de zahar, tutun etc, este stimuleaza favorabil prin aplicarea unor tratamente cu gibereline exogene.
2.6. FITOCROM
Fitocromul este un pigment cromoproteic care influenteaza cresterea si dezvoltarea plantelor, fapt pentru care mai nou este denumit fitohormon. Ca aspect caracteristic si specific pentru acest compus este ca indeplineste rolul de fotoreceptor in toate fotoreactiile reversibile ale plantelor, care au loc in zona rosie si rosu indepartate a spectrului de absorbtie a luminii solare. Fitocromul interactioneaza cu alti fitohormoni, influentand cresterea si dezvoltarea plantelor de la formaea semintei , germinatie si pana la inflorirea.
Rol si functii
Fitocromul se comporta cu un fotoreceptor in toate fotoreactiile reversibile ce au loc in zona rosie (rosu-deschis, rosu-inchis) de absorbtie a luminii. Prin absorbtia luminii fitocromul inlesneste utilizarea energiei luminoase pentru stimularea sau inhibarea a numeroase procese biologice.
Fitocromul are in organismele vegetale un rol multiplu si complex. El intervine ca factor de control in germinatia semintelor, in raspunsurile fotoperiodice si fotomorgogenetice ale plantelor, in biosinteza pigmentilor (clorofile, carotenoide, flavonoide etc.), cresterea frunzelor si a tulpinilor, formarea tuberculilor, inverzirea plantelor, biosinteza enzimelor din cloroplaste, formarea giberelinelor, contribuie la reglarea hormonala a infloririi plantelor etc. Ansamblul fenomenelor ce determina modificari in structura si culoarea unor compusi , celule sau tesuturi , sub influenta luminii constituie fotomorfogeneza. Rolul morfogenetic al luminii se deosebeste de actiunea sa in fotosinteza sau in fotocromul este fotoreceptorul principal in fotoperiodism. Morfogenezele determinate de fitocrom pot fi pozitive, cand sunt stimulate procesele de crestere , biosinteza etc. Si negative, cand diversele procese asupra carora actioneaza lumina absorbita de fitocrom sunt inhibate. Plantele raspund in mod diferential la actiunea fitocromului, in functie de actiunea altor factori, de fenofaza, starea fiziologica etc.
Fotoraspunsurile plantulelor de Sinapis alba produse sub influenta fitocromului P730, sdupa H.MOHR, 1966 se prezinta astfel:
inhibitia alungirii hipocotilului;
inhibitia translocatiei din cotiledoane;
marirea cotiledoanelor;
desfacerea lamei cotiledoanelor;
formarea trihomilor pe hipocotil;
inceputul curburii hipocotilare;
formarea de prmordii foliare;
diferentierea primelor frunze;
cresterea reactivitatii geotropice negative a hipocotilului;
formarea traheelor;
diferentierea stomatelor in epiderma cotiledoanelor;
schimbarea intensitatii respiratiei celulare;
biosinteza antocianilor;
intensificarea biosintezei acidului ascorbic si a clorofilelor;
cresterea biosintezei ARN si a proteinelor in cotiledoane;
schimbarea proportiei de mobilizare a lipidelor si a proteinelor de rezerva.
Pe langa fotoactiunile mentionate la Sinapis alba se cunosc numeroase alte fotoraspunsuri ale plantelor la actiunea fitocromului. Modul de actiune mai intim al luminii in diferitele fotoprocese este inca prea putin cunoascut. In procesul de biosinteza a antocianilor la Brassica oleracea si B. Alba se produc doua fotoreactii. Prim este determinata de rolul fotoreceptor al fitocromului si transformarea sa in cele doua forme izomere (P660 si P730) sub actiunea luminii si una determinata de predominarea radiatiilor rosii - inchise (700 - 800 nm) si albastre (400 - 480 nm), dependenta de densitatea fluxului de lumina. Aceasta actiune poarta denumirea de ,,reactie de inalta energie" Radiatiile albastre , cele din regiunile rosu-deschis si rosu -inchis actioneaza sinergic in ce priveste biosinteza antocianilor si a acidului folic, stimularea cresterii frunzelor, inhibarea cresterii hipocotilului etc. Apare insa un autogonism intre reactia de inalta energie si fitocrom in miscarile de nutatie si germinatie.
S-a mentionat ca fitocromul este fotoreceptorul pentru toate fotoreactiile reversibile ale plantelor care au loc in zona rosie si rosu indepartat a spectrului de absorbtie a luminii, c (MOORE , 1979) T.C. MOORE (1979) clasifica fotoreactiile determinate de lumina prin intermediul fitocromului in trei grupe majore. In prima grupa se incadreaza raspunsurile fotoperiodice, cum sunt raspunsurile fotomorfogenetice ca de exemplu: inductia florala, inhibarea mugurilor iarna, formarea tuburilor polenice etc. In grupa a doua se incadreaza fotoreactiile nefotoperiodice, cum sunt: inverzirea plantulelor etiolate, germinarea diferitelor tipuri de seminte, alungirea rizoizilor etc. In grupa a treia se incadreaza fotoreactiile nefotoperiodice, cum sunt: inverzirea plantulelor etiolate, germinarea diferitelor tipuri de seminte, alungirea rizoizilor etc. in grupa a treia se incadreaza fotoraspunsurile nemorgogenetice, cum sunt biosinteza antocianilor, stimularea biosintezei enzimelor cloroplastice etc.
Mecanismul intim de actiune al fitocromului, in fotoraspunsurile plantelor pe care le mediaza, este putin cunoscut. Exista conceptii si opinii ca fitocromul are un singur mecanism de actiune in toate fotoraspunsurile plantelor in care este fotoreceptor, si alti opinii care presupun existenta a doua sau mai multe mecanisme de actiune. Cercetarile ulterioare, prin folosirea unor tehnici mai superioare, mai sensibile si a unui material biologic mai bogat, vor aduce fara indoiala date noi, certe in ce priveste mecanismul de actiune a fitocromului in diferite procese metabolice pe care le influenteaza.
Fitocromul stimuleaza biosinteza giberelinelor in cloroplaste I.L. STODDARD (1968) a constatat ca extracte din fractiunile cloroplastelor din fractiunile cloroplastelor din frunzele de Brassica oleracea si hordeum vulgare au o activitate biologica similara cu a giberelinelor a caror biosinteza compartimentalizata este in cloroplaste si in alte plastide. In vitro lumina altereaza biosintezasi interconversiunile giberelinelor, in timp ce fitocromul stimuleaza aceste procese. Fitocromul stimuleaza transportul diferitelor substante prin intermediul membranelor celulare . Biosinteza si activitatea giberelinelor in plantulele etiolate depinde de continutul fitocromului. Numerosi cercetatori sustin ca fitocromul mediaza si controleaza transportul giberelinelor prin membranele toplastelor stimuland biosinteza acestui fitohormon de crestere. Fitocromului i se atribuie un rol important in realizarea ritmurilor circadiene la plante. Se stie ca la multe plante anumite procese biologice prezinta o ritmicitate sub influenta conditiilor variate din mediu natural. Aceste ritmuri pot fi anuale sau sezoniere, zilnice sau cotidiene. Ritmicitatea zilnica a proceselor biologice nu dureaza exact 24 de ore, ci intre 21 si 28 de ore, fapt pentru care ,,ritmul zilnic" sau ,, cotidian" se numeste ,,ritm circadian" (circa o zi) , ritm de aproximativ o zi, Fiecare planta se comporta ca un ,,ceas fiziologic" sub influenta diferitilor stimuli. Ritmurile endogene sunt initiate de anumiti stimuli (intensitatea si durata luminii, tempetatura , compusi chimici, tensiunea O2 etc.) dintre care alternarea perioadelor circadiene poate fi usor modificata in functie de factorii mediului ambiant. La Phaseolus multiflorus radiantii luminoase cuprinse intre 610 si 680 nm maresc, iar cele intre 690 si 850 nm reduc perioadele circadiene . la Glycine hispida intreruperea la orice timp cauzeaza o crestere a continutului de fitocrom. P660, care va fi transformat in P730. La plantele angiosperme, in cadrul ritmurilor circadiene, fitocromul are un rol insemnat in determinarea )distribuirea, delimitarea) perioadelor de lumina si intuneric, la Phaseolus multiflorus, ritmul miscarii frunzelor este stimulat de lumina rosie - deschis (600 - 700nm) si inhibat de lumina rosie -inchis (700 - 800 nm). Reactiile fotobiologice depind de pigmentii care raspund de captarea fotonilor. Astfel , daca se determina spectrul de actiune a luminii, stabilindu-se regiunile cele mai active pentru fenomenul cercetat, si se compara cu spectrul de absorbtie al diferitilor pigmenti existenti in plante, pigmentul al carui spectru de absorbtie coincide cu spectrul de actiune studiat este responsabil pentru captarea fotonilor care initiaa raspunsul plantelor. La plantele inferioare spectrul de actiune prezinta maxime in zonele albastra si rosi - deschis. La unele ciuperci fotoraspunsurile sunt determinate de zona albastra a spectrului.
Pe baza a numeroase date experimentale se poate afirma ca fitocromul este un fotoreceptor important pentru producerea ritmurilor la plantele superioare.
RETARDANTI
Retardantii fac parte dintre compusii chimici de sinteza cu ajutorul carora oamenii pot sa influenteze si chiar sa dirijeze procesele de crestere, dezvoltare si de fructificare a plantelor. Ei au actiune complexa si specifica asupra diferitelor organe si tesuturi ale plantelor. Sbstantele retardante determina o franare (reducere) temporara a cresterii organelor vegetative, in special a tulpinilor si a lastarilor laterali, stimuland in acelasi timp, in majoritatea cazurilor inflorirea si fructificarea, obtinandu-se importante sporuri de productie , atat sub aspect calitativ, cat si cantitativ. Franarea cresterii tulpinilor si a lastarilor se realizeaza prin reducerea intensitatii diviziunii celulare si a elongatiei celulelor din tesuturile meristemice subapicale. Retardantii se deosebesc de inhibitori prin faptul ca ei ,,nu blocheaza ireversibil procesele metabolice vitale ale plantelor, nu provoaca malformatii la nivelul diferitelor organe vegetative sau reproductive, iar variabilitatea intregii plante nu este diminuata".
Retardantii dupa absorbtie se acumuleaza in tesuturile tinere, in special in cele meristemice din zonele de crestere si in nodurile tulpinilor. Actiunea lor de inhibare a diviziunii si alungirii celulelor din tesuturile in care se acumuleaza se manifesta pana cand enzimele din frunze sau alte organe si tesuturi metabolizeaza retardantii si-i transforma in compusi inactivi sau in produsi finali de metabolism, care vor fi eliminati din plante. Retardantii acioneaza eficient la un nu,ar mult mai mare de plante dicotiledonate, in comparatie cu cele monocotiledonate. Actiunea lor principala, caracteristica , consta in reducerea inaltimii tulpinilor si a lastarilor prin scurtarea lungimii internodurilor, mentionandu-se pozitia verticala a plantelor.
Numerosi retardanti maresc rezistenta plantelor la actiunea nefavorabila a unor factori fizico-chimici si biologici. Ei maresc rezistenta plantelor la actiunea bolilor si a numerosi daunatori , la actiunea toxica a unor produse fitofarmaceutice si a unor metale grele. Un rol insemnat revine unor retardanti in marirea rezistentei plantelor la ger si iernare, seceta si arsita, exces de umiditate si inundatii, in soluri saraturate etc.
Prin utilizarea retardantilor in procesul de ameliorare a plantelor se vor putea obtine noi forme (linii, hibrizi, soiuri) cu potential de productie ridicat, rezistente la actiunea bolilor si daunatorilor, a factorilor pedoclimatici, cu continut ridicat in substante biologic active.
ALAR- 85 N-DIMETILAMINOSUCCINAMIC
Activitatea retardanta a unor derivati ai acizilor maleic si succinic a fost pusa in evidenta in a962 de catre J.A. RIDDEL si colab. Acidul N-dimetilaminomaleic s-a dovedit a fi cel mai activ, cu actiunea retardanta puternic asupra cresterii legumelor, vitei de vie, cartofilor, plantelor ornamentale, dar care are dezavantajul de-a fi instabil in mediul apos. Acest retardant poarta denumirea comerciala de CO-11
In a963 G.DAHLGREN si N.L. SIMMERMAN au reusit sa obtina prin sinteza acidul N-dimetilaminosuccinamic , care are mai multe denumiri comerciale dintre care cele mai uzuale sunt: Alar-85; B995; B9; SADH; Dominozid etc. Acest compus este stabil in solutii apoase si are o actiune retardanta similara cu a acidului N-dimetilaminomaleic, avand efect similar asupra acelorasi specii de plante.
Proprietati
Alar-85 este o substanta solida, putin higroscopica, foarte putin solubila in apa (aproximativ 5%); solutiile apoase sunt insa stabile. Are masa moleculara 160, iar persistenta sa activa in sol este de 3-4 saptamani. Este absorbit cu usurinta de plante, fapt pentru care se utilizeaza predominant sub forma de solutie, de concentratii diferite, intre 100 si 5000 ppm, prin stropire foliare, extraradiculare. Actioneaza asupra unui mare numar de plante cerealiere, plante legumicole, pomicole. Aplicat pe sol, in concentratie mai mare prezinta unele efecte de toxicitate. Foarte active sunt preparatele cu Alar-85 sub forma de sprayuri.
Functii si utilizari
La plante Alar-85 actioneaza atat in ce priveste inhibarea temporara a cresterii tulpinilor si lastarilor, cat si transformarea mugurilor vegetativi in muguri florali. Produce de asemenea intarzierea infloririi plantelor cu cateva zile, fapt ce protejeaza plantele de ingheturi tarzii si primavara. Previne caderea fiziologica a fructelor din luna iunie si inainte de recoltare, stimuleaza coacerea uniforma a fructelor, acumularea pigmentilor si a unor substante biologice active. Mareste perioada de pastrare a fructelor, creste rezistenta la actiunea nefavorabila a unor boli si daunatori, precum si a factorilor fizico-chimici nefavorabili. Nu prezinta efecte inhibitoare asupra enzimelor implicate in procesul de biosinteza a giberelinelor si a altor fitohormoni.Legumele sunt plantele cele mai sensibile la actiunea Alarului-85. nu are actiune asupra germinarii plantelor.Stimuleaza formarea florilor femele.
Actiunea asupra organelor vegetative.
Alar-85 este un retardant activ care poate fi utilizat atat singur, cat si in amestec cu alti retardanti (Ethrel, CCC etc; defoliatori, erbicide, insecticide, ingrasaminte etc). La plantele pomicole acest retardant este mai eficace la meri , iar CCC la peri. Este bine ca tratamentele cu Alar-85 la pomii fructiferi sa nu se faca mai tarziu de doua luni inainte de recoltarea fructelor. Reactia pomilor este maxima cand tratamentele incep sa se efectueze de la caderea petalelor , pana la aproximativ 4 saptamani dupa inflorire. In aceasta perioada optima reducerea cresterii tulpinilor si a lastarilor este direct proportionala cu cresterea concentratiei retardantilor. Dupa un an de la aplicarea tratamentelor, daca retardantii sunt catabolizati pomii revin la o cresetere normala, iar in caz contrar apar fenomene de remanenta.
Tratamentele cu Alar-85 , efectuate pe meri tinere, neintrati pe rod, au ca efect reducerea cresterii tulpinilor cu 15-38%, iar a diametrului coroanei cu 18-33% in functie de natura soiului. Efectul retardantului se resimte la aproximativ 2 saptamani de la tratament si se mentine pana spre sfarsitul perioadei de vegetatie. Daca stropirile s-au facut cu solutii mai concentrate de 2000 ppm, se produce o scurtare a tulpinilor si a lastarilor cu 50-70%, iar masa radacinilor scade cu 64%,
Tratamentele cu Alar-85 si alti retardanti , aplicate la plantele samburoase, determina reducerea cresterii tulpinilor, stimuleaza formarea mugurilor florali, fapt ce determina obtinerea unor pomi cu talie mica (pruni, ciresi, visini, piersici, caisi etc.), cu intrare timpurie pe rod, cu productii de fructe mari la hectar. Foarte eficace sunt in acest sens tratamentele cu solutie de Alar-85 de 2000 ppm, aplicate in cires si prun la sfarsitul lunii iulie. Daca se aplica la cires tratamente combinate, Alar-85 impreuna cu CCC, solutii de 2000 ppm, se realizeaza o reducere a masei ciresilor cu 30-60%, iar frunzele devin mai goase cu 10-15%. La prun, piersici, caisi etc. Tratamentele cu solutii de Alar-85 de concentratie 2000 ppm, determina o reducere a taliei pomilor cu 15-22%.
La ridiche, tratamentele cu solutii de Alar-85 , de 5000 ppm, aplicate la aparitia primei perechi de frunze adevarate, stimuleaza szbstantial cresterea radacinilor. Tratamente similare efectuate cu solutii de Alar -85, de 4000 - 8000 ppm, aplicate pe plantule de varza de Bruxelles, au ca rezultat cresterea numarului de capatani comestibile, care vor putea fi recoltate mai timpuriu decat probele martor.
La crizantemele si dalii, tratamentele cu solutii apoase de Alar-85 , de 2000 ppm, sunt utilizate frecvent pentru stimularea inradacinarii butasilor, prin formarea unor numar cat mai mare de radacini adventive. Prin Tratarea plantulelor de crizanteme, cu solutii de Alar-85, de concentratie 2,5-5% se obtin plante cu flori dense. Astfel, crizantemele din soiul Tokyo , care au inaltimea tulpinii de 80 cm, prin tratamente cu Alar-85, cu solutii de 5-15%, devin plante cu tulpini de 15-20 cm. Foarte eficiente sunt tratamentele cu solutii de Alar-85 si franarea si reglarea cresterii tuplinilor la diferite specii de Calistephus, Lilium, Gladiolus, Antirhinum, Petunia, Euphorbia, Pelargonium, Azalea, Dianthus etc. Alar-85 , aplicat sub forma de spray foliar, franeaza cresterea internodurilor la numeroase plante de cultura, indiferent de perioada de expunere a plantelor la lumina, fara sa provoace intarzierea infloririi plantelor.
Actiunea asupra organelor de reproducere.
La par , tratamentele cu Alar-85 desi nu au mare efect asupra franarii cresterii tulpinilor, ele stimuleaza substantial formarea mugurilor florali, initierea florala. La piersic, daca tratamentele cu Alar-85 se fac in perioada de repaus, se produce o intarziere a deschiderii mugurilor florali cu 6-8 zile, fapt pozitiv, care diminueaza efectele ingheturilor tarzii de primavara. Tratamentele cu solutii de Alar-85, de 2000 - 4000 ppm, aplicate la zmeura sunt foarte eficiente pentru sporirea mugurilor de rod pentru anul urmator.
La tomate , stimularea infloririi se realizeaza prin stropirea plantulelor, la aparitia primei frunze adevarate, cu solutii de Alar-85, de 1000 -2500 ppm. Prin tratamente cu solutii de Alar -85, de 2500 ppm, aplicate pe flori taiate sau ghivece, creste perioada de pastrare a florilor cu 5 zile fata de martor. La azalee, tratamentele cu solutii de 0,75-1% de Alar-85 , aplicate la plante tinere, determina o crestere a mugurilor florali de 5-10 ori.
Actiunea asupra fructificarii, maturarii si coacerii fructelor. Tratamentele cu Alar-85, aplicate la plante pomicole determina o acumulare a pigmentilor , vitaminelor, glucidelor si aminoacizilor, fapt ce contribuie la cresterea calitatii fructelor. Este indicat ca aceste tratamente sa nu se faca dupa inflorire, cel mai tarziu inainte cu doua luni de la inceperea recoltarii fructelor. Prin utilizarea unor solutii de Alar-85, de 2000 ppm, aplicate anual la meri din soiul Golden Delicicus, in fenofaza cresterii intense a lastarilor, rezulta o productie de fructecu 170% mai mare, comparativ cu plantele netratate. Nu este indicat a se aplica , la pomii tineri la 2-3 ani de la plantare, tratamente consecutive mai multi ani, cu solutii de Alar-85, de concentratii mai mari de 1000 ppm. Prin utilizarea unor solutii de Alar-85 de 2000 ppm, aplicate cu 70-80 de zile, inaintea recoltarii la soiul Ionathan, se produce o mai buna uniformizare a marimii fructelor si o mica reducere a dimensiunii lor, fapt ce prelungeste perioada la pastrare a merelor in depozite, reducandu-se pierderile prin prabusire , putrezire, evaporare etc. Se produce o reducere a respiratiei fructelor, scaderea cu 20% a formarii etenei, reducerea sticlozitatii fructelor, cresterea fermitatii pulpei; se reduc procesele biochimice care determina coacerea si caderea fructelor.
La cires, visin si cais, tratamentele cu solutii de 2000 ppm, de Alar-85, efectuate la 2-3 saptamani dupa inflorirea pomilor, determina o grabire a coacerii fructelor, cu 4-6 zile, iar caderea fructelor coapte creste cu 20-30% fapt ce usureaza recoltarea mecanica a fructelor. Daca ciorchinii de struguri sunt scufundate dupa recoltare in solutii de 500 - 600 ppm de Alar-85 , creste perioada de pastrare la rece a strugurilor , scade aciditatea titrabila, se mentine buna culoarea fructelor, a caror calitate este superioara fata de probele martor. Solutiile de Alar-85 , in concentratii mai mici, de 1000 - 25000 ppm grabesc fructificarea tomatelor, iar in concentratii mai mari, de 5000 ppm, intarzie acest proces.
Transformari metaboolice, rezistentei plantelor tratate la actiunea nefavorabila a unor factori fizico - chimici si biologici.
Tratamentele cu solutii de Alar-85, de 2000 ppm, , maresc rezistenta plantelor pomicole , legumicole si a cerealelor la actiunea prelungita a secetei, prin reducerea numarului de stomate, cresterea grosimii frunzelor si intarzierea imbatranirii lor etc. Prin tratarea merilor tineri cu solutii de Alar-85, de 2500 - 3000 ppm, se produce hidroliza mai accentuata a amidonului la inceputul perioadei de repaus, fapt ce determina acumularea glucidelor solubile care maresc rezistenta plantelor la actiunea gerului. Daca tratamentele se fac cu solutii mai concentrate, de 5000 6000 ppm de Alar-85 , la inceputul lunii octombrie, creste rezistenta la ger a mugurilor de rod, fapt ce va contribui in anul urmator la sporirea productiei. Prin stropirea salatei recoltate cu solutii de Alar-85 creste perioada de pastrare in stare proaspata a acestei legume.
Prin tratarea plantelor de varza de 4 saptamani , cu solutii de 2500 - 5000 ppm de Alar-85 creste rezistenta plantelor la temperaturi scazute si ger, creste continutul azotului neproteic si a proteinelor solubile, a amidonului si a glucidelor solubile; creste concentratia si presiunea osmotica a sucului celular, in schimb scade azotul anorganic, in special azotatii.
AMO -1618, CLORURA DE 2-IZOPROIL-E-4-TRIMESTIL-AMONIN-5-METILFENIL-1-PIPERIDIN CARBOXILA
Acest retardant , denumit prescurtat Amo-1618, a fost descoperit in 1950 de catre J.W.WIRWILLE si J.W. MITCHEL, fiind selectat ca cel mai activ retardant dintr-o serie de aproximativ 80 de carbanati de amoniu quaternari, incluzand derivati de morfolina, picolina, piperidina, piridina, quinaldina, quinolina etc.
Structura moleculara
Sub aspectul structural Amo-1618 este un carbomat de amoniu quaternar, care contine in molecula doi atomi de azot: unul carbamic si un azot de amoniu quaternar. Indepartarea acestor atomi de azot din molecula determina disparitia activitatii retardante.
In componenta structurii moleculare se mai gasesc: un carbamat de amoniu derivat din piperidina, un nucleu benzenic, pe care este grefat la C-2 un radical izopropilie, la C-4 clorura de trimetilamoniu, iar la C-5un radical metilic.
Functii si utilizari
Amo-1618 franeaza cresterea tulpinii si a lastarilor la fel ca si retardantii anteriori, fara sa apara malformatii pe frunze, tulpini, radacini, flori sau fructe. Mo-1618 , similar CCC-ului, mareste rezistenta plantelor de tomate si vinete la temperaturi negative sub -50C, la stres hidric, la boli si daunatori, fapt ce contribuie la marirea perioadei de pastrare a fructelor.
Raspunsul plantelor la tratamentele cu Amo-1618 este diferit, in functie de natura speciei si chiar a varietatilor. In timp ce solutiile Amo-1618 franeaza substantial cresterea tulpinilor de dovleac si de castravete , asupra tulpinilor pe pepene, au un efect slab. Fasolea si castravetii sunt foarte sensibili, chiar la aplicarea unor tratamente cu solutii diluate , de 10 ppm, de Amo-1618. prin inmuierea unor plante de crizanteme, in solutii de Amo-1618, a craror radacini au fost scurtate prin taiere, s-a observat histologic ca retardantul dupa patru zile inhiba diviziunea si alungirea celulelor din meristemul al tulpinii. Meristemul apical continua sa se dezvolte, celulele se inmultesc si se alungesc in prezenta retardantului. Prin aceasta actiune selectiva a retardantului Amo-1618 in ce priveste diviziunea celulelor din zona subapicala si apicala a meristemelor se poate realiza franarea cresterii tulinilor si a lastarilor, precum si stimularea formarii mugurilor florali, stimularea infloririi, grabirea fructificarii, a maturizarii si coacerii fructelor etc. In cazul frunzelor, solutiile de Amo-1618 determina la inceput o franare a cresterii acestora, dar dupa cateva zile de tratament frunzele cresc si ajung la maromea normala. La fasole cresterea celulelor din primul interval scade de 69% la tratamentele cu Amo-1618, in comparatie cu martorul. Prin sectiune transversala se observa ca celulele din cambiu se divid centripetal. Prin tratarea frunzelor mature cu solutii de Amo-1618 se produce ingrpsarea acestora cu 20%. La mazare, dupa 1 zile de la aplicarea tratamentelor cu Amo-1618 se remarca o scurtare puternica a internodurilor tulpinii, in special a internodului 3.
CCC; CYCOCEL; 2-CLORETIL-TRIMETILAMONIU
Clorura de clorcolina este unul din retardantii cei mai importanti, care are un larg spectru de actiune, avand numeroase aplicatii in agricultura, horticultura, silvicultura, pentru cultura plantelor ornamentale de apartament etc. Datorita efectelor remarcabile pe care le produce asupra cresterii si dezvoltarii plantelor, clorura de (2-cloretul) -trimetilamoniu, se produce in numeroase tari, in cantitati mari, sub diferite denumuri comerciale, dintre care cele mai uzuale sunt cele de CCC, Cycocel, Tur, Batur, Sur, ABM, Clormequat, retacel, Mirocil, Cycogan, Clorvylegac, Stabilin, R-41 etc.
Strucutra moleculara
Sub aspectul structurii moleculare CCC este un derivat al colinei, la care gruparea hidrocilica este substituita cu clor.
+
Cl-CH2-CH2-N- (CH3)3Cl-
Cycocel (CCC) , 2-cloretil-trimetilamoniu
Colina este un aminoalcool care intra in constructia lecitinelor. In organismele vegetale si animale colina se obtine din serina, printr-un proces de decarboxilare si metilare, conform urmatorului mecanism.
Functii si utilizari.
CCC are numeroase functii si variate aplicatii practice in cresterea si dezvoltarea diferitelor plante de cultura.
Actiunea asupra organismelor vegetative ale plantelor. Asa dupa cum s-a mentionat , efectul principal al retardantilor, deci si al CCC-ului , consta in franarea temporara a cresterii tulpinilor si a lastarilor. Actiunea retardantilor se exercita cu arecadere la nivelul internodurilor, care in momentul aplicarii tratamentelor trebuie sa se gaseasca in faza cea mai intensa de crestere si alungire. Prin tratamente repetate, in diferite fenofaze, se poate reduce substantial inaltimea tulpinilor si lungimea lastarilor. Asupra radacinilor CCC are in general un efect stimulator, iar asupra frunzelor, dupa 2-3 zile de la aplicarea tratamentului se produce o ingrosare a limbului foliar, creste continutul clorofilelor si al carotenoidelor, a azotului total si a proteinelor, fapt ce stimuleaza fotosinteza si biosinteza substantelor organice, care vor fi utilizate intr-o proportie mai mare de organele de reproducere ale plantelor.
Prin aplicarea retardantilor in pomicultura se urmareste reducerea cresterii pomilor in inaltime, scurtarea si ingrosarea internodurilor, scurtarea ramurilor, ingrosarea frunzelor, cresterea productiei de fructe atat cantitativ cat si sub aspect calitativ, diferentierea mugurilor de rod pentru anul urmator tratamentului etc.
Prin tratamente cu CCC, in concentratie de 1000 - 5000 ppm, aplicate la 2-6 saptamani dupa inflorirea parului se produce o substantiala reducere a cresterii vegetative si o stimulare puternica a formarii viitorilor muguri de rod pentru anul urmator. Daca CCC se aplica in concentratie de 2000 ppm, se produce o reducere a taliei perilor cu 11-40%, iar sporurile de productie cresc cu 20-40%.
La vita de vie, tratamentele cu CCC, in concentratie de 100 - 1000 ppm, aplicate cu 15 zile inainte de inflorire, au redus sintitor alungirea internodurilor, cresterea frunzelor si a carceilor, favorizand formarea unui numar mai mare de inflorescenta. CCC, la fel ca alti retardanti , controleaza repartitia substantelor nutritive intre organele vegetative si cele de reproducere pe care le favorizeaza.
Foarte eficace sunt tratamentele cu CCC la tomate. Prin utilizarea unor solutii de concentratie de 1500 - 3000 ppm, se produce, o reducere a inaltimii tulpinii si internodurilor cu 10-15%, iar in unele cazuri cu 40%.
Daca se trateaza solul cu solutii CCC, cu concentratie relativ mica de 1000 ppm, de 2-5 ori pe saptamana, se intensifica biosinteza si acumularea clorofilei, fapt ce duce la intensificarea culorii verde-inchis a limbului foliar si implicit la intensificarea procesului de fotosinteza.
La cartofi , tratamentele cu CCC, Alar, Ethrel, Fosfon-D, inhiba cresterea tulpinilor si a lastarilor laterali, favorizandu-se formarea stolonilor si a tuberculilor.
Prin stropiri repetate a garoafelor, la 8-10 saptamani dupa ciupire, cu solutii CCC de 2000 ppm, se produce o scurtare a tulpinilor cu 20% , a internodiilor cu 20% si se realizeaza concomitent o indesare a florilor. Rezultate similare cu a garoafelor, prin stropiri cu solutii de CCC; se obtin si la alte plante ornamentale: la Fresia, Zinnia, Tagetes, Salvia, primula etc. In cazul trandafirilor, prin utilizarea unor solutii 0,25% - 0,5% de CCC, cate 100 ml la un ghiveci, se reduce la jumatate lungimea tijelor florale, concomitent cu cresterea desimii, marimea si calitatea florilor.
Prin tratamente cu CCC si alti retardanti se realizeaza la numeroase plante legumicole cresterea numarului de flori femele, care vor deveni purtatoare de fructe, in detrimetul florilor mascule. Retardantii au deci aplicatii importante in determinarea si cresterea mugurilor florali, a florilor. Solutiile diluate cu CCC stimuleaza intr-o mare proportie inflorirea plantelor de tomate, cresterea numarului de flori la primele inflorescente scazand simtitor procesul de avortare a organelor femele.
In cultura gladiolelor, tratamente cu CCC aplicate pe sol , in concentratie de 8000 ppm, determina o crestere a numparului mediu de flori de la 7 la 10 , florile apar trei zile mai devreme , fiind mai mari, mai grele si mai viu colorate decat la lorul martor.
La Azalee, tratamentele cu CCC determina o puternica stimulare a mugurilor florali, paralel cu reducerea lastarilor laterali. Solutiile 1% de CCC determina o dublare a mugurilor florali.
Actiunea asupra fructuficarii , maturarii si cresterii productiei.
La cereale, plante pomicole, legumicole, la vita de vie etc. Retardantii determina o mai buna fructificare. Creste calitatea fructelor prin acumularea unor substante biologic active; se mareste perioada de pastrare; creste productia de fructe, legume si cereale la hectar, atat prin cresterea masei boabelor, fructelor si legumelor , cat si prin diminuarea caderii fructelor tinere din luna iunie sau inainte de recoltare. In perioada de crestere a fructelor scade continutul de auxine si creste continutul de etilen din fructe, factori care usureaza procesele de abscisie, datorita formarii unui strat de separare intre penduncul si fructe, frunze, legume etc. La par, tratamentele cu solutii de CCC, de 100 ppm, aplicate cu 70 - 80 de zile inaintea recoltarii, reduc aproape la jumartate fructele care cad in perioada ce precede culesul.
La diferite soiuri de vita de vie, prin tratamente cu CCC, in concentratie de 100 - 1600 ppm, aplicate inainte de inflorire este stimulata foarte mult legarea fructelor. Spre deosebire de efectele unor stimulatori, tratamentele cu CCC determina o marire a numarului de fructe cu seminte, iar in unele cazuri produc o mica reducere a greutatii acestora. Tratamentele cu CCC efectuate la vita de vie inainte si dupa inflorire stimuleaza fructificarea, cresterea boabelor si a ciorchinelor la majoritatea soiurilor realizandu-se sporuri substantiale de productie. La soiul Cocur stropirile cu solutii de CCC de 5000 ppm, au produs o crestere a productiei de struguri cu 40-60%. Tratamentele cu CCC aplicate la vita de vie determina nu numai o crestere a productiei de struguri , dar creste si calitatea acestora. In urma tratamentelor cu retardanti creste in boabe continutul substantei uscate, se acumuleaza o cantitate mai mare de glucide solubile, antociani, substante aromate , saruri minerale etc., concomitent cu scaderea aciditatii.
Cresterea rezistentei plantelor la actiunea factorilor fizico -chimici si biologici nefavorabili.
Una din caracteristicile importante ale tratamentelor cu diferiti retardanti, in special cu CCC, Alar, Ethrel, Fosfon-D, o consituie cresterea rezistentei acestora la actiunea nefavorabila a unor factori fizico-chimici (ger, seceta, salinitate, toxicitatea pesticidelor, exces de umiditate, influenta metalelor grele) si biologici (boli si diferiti daunatori).
Pentru a mari rezistenta plantelor la actiunea nefavorabila a temperaturilor scazute din timpul iernii, a brumelor de primavara si de toamna, este necesar sa se faca tratamente preventive cu retardanti pentru a se acumula substante osmotice active in sucul celular (aminoacizi, glucide solubile, polialcooli, substante minerale, vitamine etc.), care maresc presiunea osmotica si micsoreaza puntele de solidificare ale solutiilor citoplasmatice si a celor vacuolare. Tratamentele cu CCC, aplicate la sfarsitul verii si inceputul toamnei, maresc rezistenta pomilor fructiferi la temperaturi cu 2-3oC mai joase, comparativ cu probele martor. Tratamentele cu solutii 1% de CCC, aplicate la perii tineri primavara, maresc rezistenta pomilor la ger in timpul iernii si a florilor in anul urmator. Tratamentele cu CCC determina, de asemenea , o rezistenta mai mare a plantelor la actiunea nefavorabila a secetei si arsitei, prin creseterea concentratiei sucului celular si a continutului de apa legata, marirea presiunii osmotice, scaderea permeabilitatii membranelor protoplasmatice, inchiderea stomatelor, reducerea transpitatiei etc.
2,4-DNC; CLORURA DE 1 (2-4 DICLORBENZEN) -1-METIL-2-PIRIDIN-PIROLIDINICA
Sub aspectul structural acest retardant este format din 2,4 -diclorbenzil, un nucleu tetrahidropirolic metilat si un nucleu piridinic. Ca element activ un cation de azot quaternar. Atomul de azot din nucleul piridinic are valenta 5, iar cel din nucleul piridinic are valenta 3. Partea activa a moleculei, alaturi de azotul quaternar, o constituie cei doi atomi de clor substituiti pe nucleul benzenic la C-4 si C-2.
Alti compusi cu structura similara, cu atomi de clor la C-3 si C-4 sau numai la C-4 sunt mai putin activi. De asemenea compusii cu nucleu de benzen liber, fara clor sau alti substituenti sunt mult mai putin activi decat orice derivat halogenat al benzilului.
Determina franarea tulpinilor si a lastarilor la nemeroase specii de plante, fara sa apara efecte secundare negative la nivelul frunzelor, florilor, tulpinilor sau fructelor. Foarte sensibile la actiunea acestui retardant sunt plantulele de fasole, a caror internodii, in urma tratamentelor se scurteaza foare mult, in timp ce frunzele se ingroasa, biosintetizand o cantitate mai mare de clorofile si de carotenoide. Majoritatea tratamentelor cu acest retardant se fac sub forma de pasta de lanolina, in care retardantul este incorporat in proportie de aproximativ 1%. Cercetarile ulterioare au aratat ca si compusii picolinici si piridinici, care contin azot quaternar, sunt activi, franand temporar alungirea tulpinilor de fasole.
ETHREL; ACIDUL 2-CLORETILFOSFONIC
Acidul 2-cloroetilfosforic este unul din retardantii cei mai eficienti si mai frecvent utilizati in agricultura , care poarta denumirea comerciala de Ethrel. A fost sintetizat pentru prima data in 1967 de catre firma AMCHEN din Ambler - Pensylvania S.U.A. . are urmatoarea formula generala:
O
||
Cl - CH2 - CH - P - OH
|
OH
Acest retardant este un genetrator de etilen. Are actiunea retardanta asupra plantelor atat datorita actiunii sale directe asupra celulelor vegetale, cat si datorita etilenului care se fomraza prin degradarea sa- Printr-o reactie de hidroliza Ethrelul se transforma in acid clorhidric, acid fosforic si etena.
Functii si utilizari
Actiune asupra organelor vegetative.
Solutiile de Ethrel, de 200 ppm sunt utilizate frecvent pentru defolierea pomilor in pepiniera. Acest retardant grabeste caderea frunzelor, fara sa manifeste toxicitate asupra mugurilor si scortei. Tratamentele se aplica de obicei la sfarsitul lunii octombrie. Foarte eficace sunt amestecurile solutiilor de ethrel de 2000 ppm, cu S2-4 0,75% sau MgCl2 0,25% , iar raport de 2: 1 in colum.
La vita de vie, stropirea cu solutii diluate de Ethrel au determinat reducerea cresterii lastarilor, la diferite soiuri , cu 27% - 53%. La ridiche, solutiile de Ethrel, sub 10 ppm stimuleaza formarea radacinilor. La ceapa tratamentele cu solutiile de Ethrel , de 1000 - 5000 ppm au grabit formarea bulbilor cu 1-2 saptamani mai devreme decat probele martor. Tratamentele cu Ethrel, pe sol sau extraradicular, au redus mult inaltimea unor plante de gura leului, gladiole, stanjeni, petunii, crini, margarete etc,. Si au intarziat inflorirea acestor plante cu cateva zile.
Actiunea asupra organelor de reproducere
La castravete, solutiile de ethrel, de 50 - 500 ppm, sunt frecvent utilizate pentru stimularea formarii florilor femele, care sunt geberatoare de fructe. Stropireile se efectueaza cand plantulele au 1-4 frunze adevarate. In unele experiente, stropirile cu solutii de Ethrel , de 1000 ppm, au determinat o crestere substantiala a florilor femele, concomitent cu reducerea de 10 ori a florilor mascule. Daca raportul dintre florile femele si cele mascule la plantele netratate era de 1:8, la cele tratate au ajuns la 30:1. ca aspect pozitiv merita semnalata constatarea ca tratamentele cu Ethrel determina o inflorire stimulanta a florilor femele, grabeste productia timpurie la castraveti , usurandu-se recoltarea mecanica a acestora.
La crizanteme, solutiile diluate de Ethrel intarzie deschiderea mugurilor florali si prelungeste perioada de crestere a plantelor.
Actiunea asupra fructificarii , maturarii, coacerii fructelor si semintelor.
Prin tratamente cu solutii de Ethrel , de 500-750 ppm, aplicate cu 10-14 zile inainte de recoltare, se produce la merele de vara grabirea coacerii, intensificarea culorii fructelor si se usureaza mult desprinderea fructelor de pe pom. La par dupa tratamente similare cu solutii de 500 - 2000 ppm, apar si aspecte negative care constau in cresterea procentului de fructe prabusite , dupa 50 de zile de pastrare la temperatura de 0oC.
Pentru a usura recoltarea mecanica a fructelor de prun, cires, visine,coacerea negre etc se fac stropiri cu Ethrel, cu 7-14 zile inaintea recoltarii. La visini se recomanda cu solutii sa aiba o concentratie de 500 ppm, la cires de 750 ppm, iar la coacaze de 300 ppm.
La vita de vie , tratamentele cu solutii de Ethrel, de 500 - 1200 ppm au grabit maturarea fructelor, a crescut continutul pigmentilor , a glucidelor solubile, masa boabelor si a scazut aciditatea.
Prin efectuarea unor stropiri cu Ethrel pe frunze de tomate in concentratie de 250 - 500 ppm, se realizeaza o crestere a productiei cu 56%. Se produce atat grabirea cat si uniformizarea coacerii, fapt ce determina ca 85-90% din fructe sa fie de calitate superioara. Prin stropirea cu solutii de Ethrel , de 250 - 200 ppm, efectuate la tomate in diferite faze de formare a fructelor, se asigura o coacere simultana a fructelor , care au o culoare intensa, o forma usor alungita, creste continutul de vitamina C, se usureaza recoltarea mecanica, creste sintitor productia de fructe. Daca stropirile se fac cu 11-14 zile inainte de inceperea recoltarii se grabeste maturarea fructelor cu 7-12 zile. Coacerea fructelor de tomate este stimualata si de etena care se formeaza in procesul de degradare a acestui retardant.
FOSFON-D; CLORURA DE 2,4-DICLORBENZIL-TRIBUTILFOSFONICA
Activitatea retardanta a acestui compus, denumit prescurtat Fosfor D a fost anuntata in 1995. acest retardant este clorura unui cation de fosfoniu quatrenar, care formeaza legaturi covalente cu trei radicali de butil si unul de 2,4-diclorbenzil.
Functii si utilizari.
Asa dupa cum s-a mentionat , tratamentele cu Fosfon D sunt utilizate frecvent la plantele floricole. Activitatea selectiva a acestui retardant se manifesta chiar si la plantele din acelasi gen. Astfel, prin efectuarea unor stropiri cu Fosfor D la Lilium longiflorum se produce franarea tulpinii fara sa apara efecte secundare negative la nivelul frunzelor. Prin stropiri efectuate cu aceeasi solutie la L.speciosum, apar fenomene toxice, de cloroza in imbrunare a frunzelor. La ambele specii de Lilium se produce o ingrosare a membrenele celulare, astfel incat plantele tratate sunt mai rezistente la cadere.
Fosfor D stimuleaza formarea si dezvoltarea mugurilor florali la speciile Rhadodendron , ilex etc. Grabeste inflorirea , cresterea si dezvoltarea florilor. La tomate si vinete tratamente cu Fosfor D si fosfon S maresc rezistenta plantelor la temperaturi scazute, la actiunea unor boli si daunatori la stresul hidric etc, asigurand o perioada mai mare de pastrare a fructelor. Tratamentele cu Fosfor D la cartofi determina inhibarea cresterii tulpinilor aeriene si a lastarilor, in schimb stimuleaza formarea stonilor si a cartofilor. Creste continutul substantei uscate din cartofi, in special continutul amidonului, proteinei brute, iar in frunze creste continutul pigmentilor asimilatori. La plantele tratete procesul de respiratie a cartofilor este mai redus, fapt ce asigura o perioada mai mare de pastrare a acestora.
La ceapa, prin tratarea plantelor cu Fosfor D, cu solutii de 480 ppm, se reduc inaltimea lujerilor (tulpini florifere), fapt ce usureaza recoltarea mecanica a inflorescentei.
β-HIDROXIETILHIDRAZINA (BOH)
Aceste retardant a fost descoperit in 1955, de catre GOWING D.P. li LEEPER R.W. , care au observat ca acest compus grabeste inflorirea la ananas.
Ca aspect caracteristice in ce priveste structura moleculara se remarca lipsa unui inel homociclic sau heterociclic si lipsa legaturilor nesaturate carbon-carbon, duble sau triple.
Acest produs nu prezinta activitate retardanta in ce priveste franarea cresterii tulpinilor la mazare si nici in ce priveste producerea curburii coleoptilelor la ovaz. Este activ insa in franarea cresterii tulpinilor la tomate, fasole, castraveti, dovleac, pepene etc. Avand efecte pozitive asupra organelor generative.
INHIBITORI DE CRESTERE
Inhibirotii naturali si cei obtinuti prin sinteza sunt substante care inhiba cresterea tulpinilor, lastarilor, radacinilor, germinatia semintelor, cresterea si dezvoltarea mugurilor, incoltirea cartofilor, a bulbilor etc.
Inhibitorii de crestere pot actiona atat asupra plantelor intregi, cat si numai asupra unor organe si tesuturi , determinand inhibarea unor importante procese metabolice, dintre care mentionam : biosinteza acizilor nucleici, biosinteza si degradarea proteinelor, fotosinteza cloroplastelor, respiratia celulelor, fosforilarea oxidativa etc, precum si procesele fiziologice de organogeneza, diviziunea si alungirea celulelor, germinarea semintelor, pornirea si deschiderea mugurilor, incoltirea cartofilor etc. Tratamentele cu inhibitori se fac atat in vivo, pe planta integrala sau pe anumite parti ale acesteia , cat si in vitro, pe sectiuni din diferite organe si tesuturi ale plantelor, culturi de celule si tesuturi. Etc.
Din nefericire , majoritatea covarsitoare a experientelor privind stabilirea efectelor biochimice si fiziologice ale inhibitorilor de crestere a mecanismelor de actiune, s-au determinat pe baza unor teste biologice, cu inhibitori extrasi din material vegetal (celule, tesuturi, organe) si nu interiorul plantelor , in celulele si tesuurile intacte, fapt ce poate cauza aparitia unor modificari conformationale ale inhibitorilor, care sa determine aparitia unor centre metabolice diferite de cele care actioneaza in celulele naturale intacte. Din aceasta cauza se pot savarsi interpretari gresite in ce priveste mecanismele intime de actiune.
Intr-o importanta lucrare de sinteza , din 1971 V.I. KEFELI si C.Sh. KADAYROV au prezentat urmatoarele caracteristici generale ale inhibitorilor naturali de crestere:
continutul inhibitorilor creste in timpul perioadei de depresie a cresterii si descreste in perioada activa de crestere a plantelor;
inhibitorii sunt capabili sa inhibe cresterea unor organe, tesuturi ti sectiuni izolate;
ei nu poseda activitate antihormonala specifica , dar ei toti pot inhiba activitatea de crestere a plantelor determinata de fitohormoni;
inhibitorii nu sunt capabili sa inhibe toate formele de crestere ale plantelor;
ei nu determina o stimulare specifica pentru auxine, gibereline si citokinetine;
care regula generala inhibitorii naturali de crestere se gasesc in tesuturile verzi tinere, impreuna cu auxine, gibereline si citokininele; pe cand inhibitorii singuri se acumuleaza in absenta fitohormonilor, in organele de repaus si in cele imbatranite;
intre inhibitorii de crestere si fitohormoni se produc interactiuni care asigura cresterea normala a plantelor;
inactivitatea si degradarea inhibitorilor se produce in perioada activa de crestere a plantelor:
inhibitorii se acumuleaza in cantitate mai mare in plantele lemnoase, in perioadele de repaus ale acestorA;
biosinteza intensa a inibitorilor se produce in tesuturile verzi;
organele si tesuturile aflate in stare de repaus isi pierd capacitatea de a distruge inhibitorii naturali;
degradarea inhibitorilor de crestere se produce inaintea germinarii semintelor, deschiderii mugurilor, incoltitii cartofilor etc;
ei au capacitatea de a inhiba germinatia semintelor, pornirea si deschiderea mugurilor, incoltirea cartofilor, a cepei, cresterea dreapta a coleoptilelor etc.
Prezenta in plante a inhibitorilor endogeni s-a pus in evidenta destul de tarziu, doar in ultimele 3-4 decenii, datorita prezentei concomitente in acelasi tesuturi si organe vegetale, atat a inhibitorilor cat si a stimulatorilor de crestere, cu functii antagoniste. Plantele, la fel ca si animalele, pot realiza fenomene de autoinhibare totale, care cuprind intregul organism, sau numai partiale, care cuprind unele tesuturi sau organe, diferite procese biocimice si fiziologice.



B I B L I O G R A F I E
DAVIDESCU V - ,, Produse fitofarmaceutice" - Editura Ceres - Bucuresti 1970 p.119
CHERGHI A, BURZO I, MARGINEANU L, DENES S, DOBREANU M, PATTATYUS K - ,, Biochimia si fiziologia legumelor si fructelor" - editura Acad.R.S.R. - Bucuresti - 1983 -
DAVIDESCU VELICICA - ,,Fitohormoni sintetici cu aplicatii in agricultura" - Redactia revistelor Agricole - Bucuresti - 1969 . p.4.
NEAMTU G. - ,,Biochimie vegetala" - Editura ceres - Bucuresti - 1960 - p.208.
Revista ,,Profitul Agricol" - nr.3 (18 ian 2006) p.15
Revista ,,Profitul Agricol" - nr.28 (15-30 octombrie 2005) p.17
Revista ,,Profitul Agricol" - nr.16 (22 - 31 mai 2005) p.15
Revista ,, Profitul Agricol" - nr.3 (18 ian 2006) p.16
Revista ,,Profitul Agricol" - nr.4 (20 - 28 februarie 2005) p.14
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 9130
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved