| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
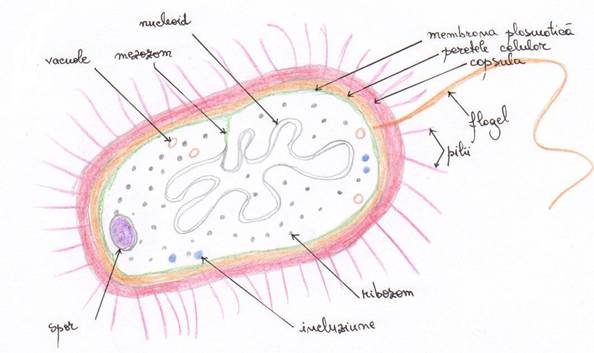
CELULA PROCARIOTA
Celula procariota este caracteristica bacteriilor si algelor-verzi (cianobacteriile). Celula procariota are dimensiuni microscopice si prezinta cateva caracteristici proprii.
materialul genetic (ADN) nu este delimitat printr-o membrana, fiind localizat direct in citoplasma;
ADN-ul nu este asociat cu proteine de tipul histonelor
peretele celular contine aproape intotdeauna mureina (un peptidoglican=polizaharid)
se multiplica, de obicei prin sciziparitate (impartire in doua)
Descrierea constituentilor structurali ai celulei bacteriene a fost posibila datorita perfectionarii tehnicilor de studiu in special cele de citochimie si microscopie electronica.
Elementele structurale ale celulei bacteriene pot fi clasificate in doua categorii:
elemente structurale constante intalnite la toate bacteriile: perete celular, mambrana plasmatica, citoplasma, material nuclear, organite citoplasmatice si incluziuni citoplasmatices
elemente structurale inconstante intalnite numai la unele specii bacteriene: capsula, glicocalixul, sporul bacterian, flagelii, pilii si cromatoforii.
Peretele celular - este situat in afara membranei plasmatice
Este alcatuit dintr-o retea de molecule de mureina (peptidoglican), care poate fi inconjurata de o membrana lipoproteica, puternic incarcata negativ. Aceasta membrana are rolul de a emite flageli. Mureina intervine in procesele de imunitate si patogenitate.
Exista si procariote lipsite de peretele celular (micoplasmele). Peretele celular este bine definit structural si cu o consistenta rigida; creste prin extindere moleculara.
Examinat la microscopul electronic, peretele celular are aspectul unui sac gol cu o constitututie asemanatoare unei plase. Grosimea lui variaza intre 15-35nm. Este in general mai subtire la toate celulele tinere care cresc rapid.
La celulele tinere cu citoplasma abundenta, compacta si omogena, peretele celular adera intim de continutul celular subadiacent. La celulele batrane, in curs de liza (plasmoliza), peretele celular devine evident, fiind indepartat de citoplasma.
Semnificatia biologica
conserva integritatea celulei fata de cresterea presiunii osmotice interne, asigurand protectie fata de socul osmotic. Presiunea osmotica interna este in jur de 5-6 atm (ex: E. coli) dar poate fi si de 20-30 atm (ex: Staphilococ). Fara suportul rigid al peretelui celular, membrana plasmatica nu ar putea rezista acestei presiuni.
peretele celular reprezinta sistemul de sustinere mecanica al intregii arhitecturi celulare. Datorita rigiditatii sale, determina forma caracteristica a celulelor.
participa la procesele de crestere si diviziune celulara, urmand membrana plasmatica in formarea septurilor transversale care vor separa celula mama in 2 celule-fiice.
participa indirect la deplasarea bacteriilor ciliate, constituind suportul rigid pe care se sprijina flagelii in contractiile lor; este suport de fixare pentru cili si flageli.
Plasmalema sau membrana plasmatica acopera citoplasma celulei separand-o de suprafata interna a peretelui celular de care este de obicei strans lipita. Este alcatuita din fosfolipide si proteine, dupa modelul "mozaicului fluid" (asemanator celui de la eucariote) . Are grosime fixa 7,5-10 nm, nu prezinta pori, nu permite endosimbioza, este impenetrabila, isi mareste suprafata prin pliere si invaginare de unde se diferentiaza mezozomii.
Semnificatia biologica
- este o componenta permanenta a celulelor cu rol de delimitare a spatiului intracelular; ea reprezinta o bariera osmotica a celulei;
- este semipermeabila, asigurand o rapida echilibrare a ionilor de Na intre interiorul si exteriorul celulei, o concentrare intracelulara a ionilor de K si Mg precum si a unor metaboliti chiar daca concentratia lor extracelulara este mai mica.
- transportul prin membrana a substantelor nutritive sau a celor rezultate din metabolism e realizat in mod activ de sistemul permeazelor.
- membrana plasmatica este sediul a numeroase enzime respiratorii, reprezentand o importanta structura cu rol energetic.
- coordoneaza cresterea si diviziunea celulelor initiind replicarea cromozomului bacterian si asigurand separarea cromozomilor-fii prin formarea septului transversal de diviziune (prin unirea a doi mezozomi)
Membrana plasmatica la bacterii se deosebeste de cea a celulelor eucariote pentru ca prezinta numeroase invaginari de diferite forme: viziculare, lamelare, tubulare care patrund in citoplasma. Aceste invaginari se numesc mezozomi (la bacteriile G+ si impreuna cu membrana plasmatica formeaza sistemul membranar unic sau unit-membrana.
Mezozomii sunt invaginari ale membranei si pot fi de 3 tipuri interconvertibili:
mezozomi lamelari rezultati prin plierea membranei plasmatice
mezozomi tubulari de forma unor tubusoare lungi
mezozomi veziculari sau saculiformi sub forma unor vezicule aproximativ sferice.
Mezozomii sunt implicati in metabolism si procesele de multiplicare celulara. Ei stimuleaza formarea septului transversal in timpul diviziunii si fixarea/ancorarea ADN-ului bacterian de membrana plasmatica si apoi separarea acestuia in doua entitati ce vor forma materialul nuclear al celulelor-fiice.
Pot avea si alte functii (respiratie), pe ele localizandu-se enzime ce intervin in sinteza de ATP. De asemenea pot fi implicati si in secretia unor enzime celulare/al penicilinazei.
Citoplasma este un sistem coloidal complex (3000-6000 macromolecule) care se comporta ca un gel. Cuprinde: proteine, lipide, glucide, apa, saruri minerale si substante de alta natura.
In general e lipsita de curenti citoplasmatici, deci elementele componente au pozitii fixe. Nu are o origine definita similara celei celulei eucariote ce contine organite diferentiate si delimitate de membrana.
Aspectul ei e variabil funtie de varsta celulei si de conditiile de mediu:
la celulele tinere si in conditii normale de dezvoltare, citoplasma adera de peretele celular si se prezinta ca o masa densa, omogena si intens colorabila;
la celulele batrane, citoplasma isi pierde treptat afinitatea tictoriala, se indeparteaza de peretele celular si capata o structura granulara cu multe vacuole.
In interiorul citoplasmei se gaseste material nuclear (citoplasma mentine structura si localizarea nucleoidului), incluziuni, vacuole si alti constituenti.
O caracteristica a citoplasmei bacteriilor este prezenta unei mari cantitati de ARN care-i confera caracterul de bazofilie si care e mai intens la celulele tinere.
Nucleoidul (procarion, genofor)
Spre deosebire de celulele eucariote ce au un nucleu cu structura bine definita, marginit de membrana si continand un numar finit de cromozomi capabili de diviziune meiotica, materialul nuclear bacterian are forma primitiva de organizare:
- e lipsit de membrana nucleara;
- este situat in partea centrala a celulei;
- nu sufera modificari de tip mitotic in cursul ciclului de dezvoltare.
Datorita caracteristicilor sale particulare, aceasta structura a fost desemnata cu numerosi termeni: nucleosid, nucleosom, material nuclear, nucleoplasma, cromozom bacterian.
Din punct de vedere chimic, materialul nuclear contine: 60% ADN, 30% ARN si 10% proteine (NU histone). ARN-ul si proteinele contribuie la conservarea unitatii structurale a nucleoplasmei in lipsa unei membrane proprii.
Evidentierea materialului nuclear la microscopul fotonic se face numai prin colorare selectiva dupa indepartarea ARN-ului citoplasmatic ce se poate realiza prin hidroliza acida sau enzimatica (se folosesc ribonucleaze). Cu aceste tehnici, materialul nuclear apare sub diferite forme: granule sferice sau ovalare, haltere, bastonase izolate sau in "V", filamente, etc.
Pe micrografiile electronice se constata localizarea materialului nuclear in partea centrala a celulei. El se prezinta ca o zona clara cu o densitate medie mai mica decat a citoplasmei inconjuratoare. La celelalte celule (eucariote), materialul nuclear e intotdeauna mai dens decat citoplasma.
Pe sectiuni ultrafine s-a observat ca regiunea nucleara e plina cu filamente foarte fine aranjate in siruri ondulate, paralele, care seamana cu o jurubita de ata cateodata rasucita. Aceste filamente sunt formate din ADN si corespund cromozomului bacterian.
Prin metode speciale, cromozomul bacterian a putut fi extras si e alcatuit dintr-o singura molecula gigant de ADN de forma circulara cu capetele unite si cu o lungime de 1400m si cu diametrul de 2,5nm.
Cromozomul bacterian este puternic rasucit si spiralat, superrasucirea moleculei de ADN se poate face in sens orar - superrasucire pozitiva si in sens anti-orar - superrasucire negativa.
Superrasucirea pozitiva consta in doua randuri de bucle. Buclele mari pot fi in numar de 40→100, fiecare bucla se pliaza apoi a doua oara, fiecare rasucire secundara, fiind formata din 400-500 perechi de nucleotide. Aceasta duce la concentrarea moleculei de ADN la dimensiuni subcelulare. Astfel, o molecula de ADN cu un diametru de 300 micrometri poate ajunge sa atinga o lungime de 3-5 micometri si un diametru de 1,5-2 micrometri.
Superrasucirea negativa apare in regiunea furcii de replicare si determina desrasucirea celor doua catene care se dispun astfel paralel pe o mica distanta. Procesul este important in initierea sintezei proteice.
Dimensiunile nucleoidului bacterian oscileaza intre 5-16% din volumul celulei. El este mai mic in faza de repaus, cand celulele au un singur cromozom, deci sunt uninucleate.
In faza de crestere activa, in culturi tinere, pe medii optime, ele apar multinucleate, avand 2-4 cromozomi identici genetic, din care cauza, indiferent de aspectul morfologic al materialului nuclear, din punct de vedere genetic, bacteriile sunt organisme haploide.
Existenta a doi sau mai multi cromozomi intr-o celula bacteriana e explicata prin faptul ca replicarea cromozomului precede diviziunea celulara - deci e rezultatul unei lipse de sincronizare dintre ritmul de crestere si diviziunea celulara.
Pe cromozomul circular bacterian se dispun cateva mii de gene, care formeaza o singura grupa de linkage. Punctul de pornire al cromozomului bacterian este pe membrana celulara si se numeste mezozom.
Semnificatia biologica este data de prezenta ADN-ului = purtator al informatiei genetice. El asigura transmiterea caracterelor geno- si fenotipice la celulele rezultate in urma diviziunii, favorizand perpetuarea speciei pe termen indefinit - pana la aparitia mutatiilor.

Ribozomii (granulele lui Palade) sunt particule nucleoproteice citoplasmatice de forma aproximativ sferica, alcatuiti din ARN (75%) si proteine (25%). Se pot caracteriza dupa constanta lor de sedimentare la ultracetrifuga si prin capacitatea lor de a participa in vitro la sinteza proteinelor. In general constanta de sedimentare este de 70S. La procariote in faza de crestere activa se gasesc in citoplasma 1500-100 mii (in medie 20-30 mii/celula)
Ribozomii sunt componentii esentiali ai sinstemului de traducere a informatiei genetice reprezentand adevarate fabrici de proteine. Au rol in sinteza proteinelor realizata pe baza informatiei din ADN copiata de catre ARNm.
Aparatul fotosintetic este prezent la unele bacterii, care pot realiza fotosinteza oxigenica sau anoxigenica. Acest aparat este reprezentat de catre cromatofori = vezicule relativ sferice, lamele de forme si marimi diferite sau tilacoide = saci membranosi (cianobacterii)
Cromatoforii sunt organite citoplasmatice intalnite la bacteriile fotosintetizante purpurii. Ei contin totalitatea bacterioclorofilelor, ficocianine, ficoeritrine, pigmenti carotenoizi, proteine, citocromi, fosfolipide si o cantitate de Fe neheminic. Cercetarile electrono-microscopice au evidentiat forma vaziculara sau lamelara a cromatoforilor care sunt an contact direct cu membrana plasmatica din care deriva, facand parte din unit-membrana. Numarul cromatoforilor este in raport invers proportional cu intensitatea luminii si direct proportional cu cantitatea de bacterioclorofila.
La bacteriile fotosintetizante verzi aparatul fotosintetic este total diferit: celulele contin un numar mare de vezicule de cholobium, alungite, dispuse in citoplasma periferica aproape de membrana plasmatica dar fara a fi in contact cu ea. Fiecare vezicula prezinta o membrana proprie si fina unistratificata si care nu face parte din unit-membrana.
Intreaga cantitate de bacterioclorofila este localizata in aceste vezicule. Astfel aceste bacterii fotosintetizatoare verzi sunt singurele organisme fototrofe la care aparatul fotosintetic nu este legat de unit-membrana.
Incluziunile sunt formatiuni strcturale ce cuprind o serie de substante inerte depuse in citoplasma celulei si cu rol de substanse de rezerva. Dupa structura lor evidentiata la microscopul electronic incluziunile citoplasmatice sunt de 2 categorii:
A. incluziuni inconjurate de membrane in care se includ:
a) incluziuni de poli-β-hidroxibutirat
b) incluziuni de sulf
c) carboxizomi
B. incluziuni inconjurate de membrana, in care sunt incluse:
d) incluziuni de polimeri anorganici
e) incluziuni polizaharidice
f) rhapidozomi
g) magnetozomi
A. a) Incluziuni de poli β-hidroxibutirat se prezinta in celulele vii sub forma unor granulatii rotunde foarte refrigerente si reprezinta un material de rezerva tipic pentru procariote
b) Incluziuni cu sulf (granule cu sulf) apar sub forma unor corpusculi stralucitori la 2 grupe ecologice de bacterii: bacterii sulfuroase purpurii (ex: G.Chromatium) si bacterii sulfuroase filamentoase nepigmentate (ex: G. Beggiatoa, Thiothrix) Se gasesc in cantitati mari la bacteriile care cresc in medii bogate in sulf, respectiv H2S si reprezinta un depozit pentru ca ele dispar prin oxidarea sulfului daca aceste bacterii sunt transferate pe un mediu sarac in H2S.
c) Carboxizomii sunt incluziuni cu un profil poliedric in numar variabilde 1-200 si sunt prezenti la bacteriile care fixeaza CO2 pe calea pentozofosfatilor. Sunt localizate in vecinatatea nucleoplasmei si reprezinta locul de depozitare a enzimelor cu activitate in fixarea CO2 (ex: ribulozo-1,5-difosfat carboxilaza)
B. d) Incluziunile de polimeri anorganici (incluziuni de volutina) apar sub forma unor granule sferice, fara structura interna si situate in regiunea nucleoplasmalemei. Granulele de volutina reprezinta o rezerva de fosfor si de energie intracelulara pentru celula bacteriana.
Datorita continutului mare de polifosfati anorganici produc efecte metacromatice (schimba culoarea colorantului). Ele se coloreaza cu albastru de metilen in rosu-violet, motiv pentru care au fost denumite granulatii metacromatice.
e) Incluziunile de polizaharide pot fi incluziuni de glicogen si incluziuni de amidon si se intalnesc ca substante de rezerva la bacteriile aerobe sporulate (ex: G. Bacillus). La microscopul fotonic, aceste incluziuni pot fi observate dupa o prealabila colorare cu I2 in KI (solutie Lugol) si care se coloreaza cele de glicogen in brun-roscat iar cele de amidon in albastru in contrast cu citoplasma care se coloreaza in galben-pal.
f) Rhapidozomii sunt particule ribonucleoproteice sub forma de bastonase scurte, rigide; se gasesc intra sau extracelular si pot fi eliberate din celula prin liza ei. Exista la numeroase specii bacteriene (ex: G. Pseudomonas, Bacillus, Actinomyces si mixobacterii). Contin proteine si ARN in procent de 2:1 iar semnificatia lor biologica nu este inca cunoscuta.
g) Magnetosomii contin Fe si au rol in orientarea si migrarea unor bacterii sub influenta campurilor magnetice slabe. Exista la unele bacterii magnetoelectrice din sedimentele marine sau de apa dulce. Nu se gasesc dispersati in celula, ci formeaza 1-2 formatiuni intracelulare care sunt compuse la randul lor din 5-41 particule de forma cubica sau octaedrica si se gasesc in aproape toata suprafata interna a membranei plasmatice. Majoritatea contin Fe sub forma de magnetita si actioneaza in celula bacteriana asemanator unei busole biomagnetice asigurand orientarea si deplasarea bacteriilor in camp magnetic.
Vacuolele sunt structuri relativ sferice cu diametrul de 0,3-0,5m mai putin refrigerente decat citoplasma si care apar in faza de crestere activa a celulei. Numarul lor este variabil in raport cu conditiile mediului, fiind cuprins in medie 6-20. In interiorul lor exista diferite substante dizolvate in apa si sunt inconjurate la periferie de un invelis lipoproteic unistratificat, numit tonoplast.
Dupa unii autori, ele ar constitui o entitate permanenta absolut necesara celulei procariote; dupa altii ar fi o stare temporara care apare sub influenta conditiilor de mediu. Ele indeplinesc rolul de reglare a presiunii osmotice in raport cu mediul si de asemenea sunt depozite de substante de rezerva.
In afara de aceste vacuole la procariote mai intalnim un tip de vacuole numite vacuole cu gaz, aerosomi sau corpi de flotatie care sunt structuri prezente doar la procariote fiind intalnite la bacteriile imobile acvatice (ex: cianobacterii si bacterii fotosintetizatoare rosii si verzi). La microscopul electronic ele apar ca un organit compus dintr-o serie de vezicule cu gaz al caror numar, aranjare si localizare determina marimea si forma vacuolei. Bacteriile care contin vacuole cu gaz au o densitate globala mai mica decat apa si de aceea plutesc la suprafata acesteia.
La celulele care plutesc la suprafata apelor, aceste vacuole joaca si un rol de protector de ecran, avand un efect de dispersie a luminii si apara astfel structurile fotosensibile de expunere la intensitate daunatoare a luminii. Aceste vacuole cu gaz regleaza raportul suprafata/volum a celulelor bacteriene.
Sporii La unele specii bacteriene forma vegetativa (forma de crestere si multiplicare a unei bacterii) se poate transforma in spor. Aceste bacterii se numesc sporogene.
Sporul are o ultrastructura, compozitie chimica si enzime diferite fata de formele vegetative, precum si o rezistenta deosebita la conditiile nefavorabile de mediu. Aproape toate bacteriile sporogene sunt Gram+
Sporul tipic bacterian este o formatiune de forma sferica sau ovala, refrigerenta cu dimensiunile variabile functie de specie si in cadrul aceleiasi specii functie de varsta culturii si conditiile de mediu.
La unele specii diametrul sporului poate depasi diametrul transversal al celulei: sporii defromanti. In medie acesta are dimensiunile intre 0,5-0,9/1-1,5m.
Asezarea sporului in celula vegetativa este diferita si reprezinta un criteriu de identificare si clasificare a speciei. Astfel sporul poate ocupa pozitie centrala (Bacillus anthracis), pozitie subterminala (B. cereus) sau pozitie terminala (Clostridium tetani). Datorita acestui fapt speciile cu sporul central au forma de fus, de racheta de tenis daca sporul are pozitie subterminala sau de bat de toba ori ac cu gamalie cand sporul etse terminal.
La majoritatea speciilor sporulate sporul este unic si numai exceptional s-au descris mai multi spori in aceeasi celula bacteriana (aceasta situatie se datoreaza unei nesincronizari ce are loc in timpul sporogenezei.
Stuctura sporului este cu unele mici variatii aceeasi la toate bacteriile. Microscopia electronica a pus in evidenta unrmatoarele elemente structurale:
tunicile (invelisurile sporale) formate din 3 straturi suprapuse
a) stratul extern (exina)
b) stratul mijlociu
c) stratul intern (intina)
Aceste straturi elecrono-opace reprezinta 20-30% din greutatea uscata a sporului si contin 80% din proteinele sporale.
cortexul este o zona elecronotransparenta situata sub tunica interna sporala si reprezinta 10-20% din greutatea uscata a sporului. Acesta contine o substanta spacifica numita acid dipicolinic responsabil de termorezistenta sporilor.
membrana sporoplasmei = membrana plasmatica a celulei vegetative
sporoplasma (inima sporului) reprezinta 60-70% din greutatea uscata a sporului si contine elemente biologic active. In ea se disting ribozomii si nucleoplasma.
ADN-ul sporal (32-54%) in medie 50% din cantitatea existenta in celula vegetativa de origine. Aici se mai gasesc ARN ribozomal si lipseste ARN mesager. In afara tunicilor sporale se gaseste un strat extern mucos de forma neregulata numit exosporium. Natura chimica si functiile acestei formatiuni nu se cunosc inca.
Compozitia chimica Sporul contine in general aceleasi componente chimice ca si celula vegetativa, dar in raporturi diferite. Sporul este mai bogat in ioni de Ca, Mg si mai sarac in K si P. Caracteristica sporului este prezenta acidului dipicolinic sub forma de dipicolinat de Ca si care ii confera rezintenta. Sporul este mai sarac in enzime decit celula vegetativa, iar enzimele existente sunt inactive. Sporul este metabolic inactiv.
Semnificatia biologica Sporul reprezinta o etapa normala a ciclului de dezvoltare a bacteriilor sporogene. El se deosebeste de celula vegetativa prin mai multe caracteristici si se comporta ca mo forma de rezistenta ce pastreaza intacte toate caracterele genotipice si fenotipice ale speciei. Sporul nu este o forma de multiplicare ca la fungi. Rezistenta speciei bacteriene sporulate la factorii nefavorabili din mediu este mare. Sporii unor bacterii rezista cateva ore la 120ºC (caldura umeda) si 200º (caldura uscata). S-au gasit spori viabili dupa mai bine de 100 de ani in sol (ex: Bacillus anthracis)
Glicocalixul se prezinta ca o masa de filamente polizaharidice atasate de lipopolizaharide prezente pe suprafata bacteriilor Gram-negative, formand in ansamblu o structura asemanatoare unei pasle pe suprafata celulei. Asigura fixarea ferma si adesea specifica a celulei de alte celule. Cateodata diviziunea celulei in interiorul glicocalixului poate produce o microcolonie de celule acoperite de un glicocalix comun. Glicocalixul este absent in culturile pure de laborator. El se formeaza numai la celule aflate in conditii naturale.
Semnificatia biologica Prezenta glicocalixului care determina uneori formarea unei microcolonii creeaza conditii favorabile de nutritie prin mentinerea si concentrarea enzimelor degradative eliberate de catre bacterii si active pe subtratul de fixare. In acelasi timp el creeaza un rezervor de hrana pentru bacteriile eliberate prin digestia enzimatica a celulelor moarte.
In cazul bacteriilor patogene prezenta glicocalixului poate determina un efect protector complex pentru celule, asigurand fixarea si mentinerea acesteia in mediile naturale sterile precum si o rezistenta marita fata de bacteriile pradatoare, bacteriofagi si protozoare.
Prezenta glicocalixului impiedica fagocitarea bacteriilor de catre leucocite, precum si fixarea anticorpilor din organismele infectate. In natura, in apele repezi de munte, glicocalixul asigura ancorarea bacteriilor de diferite substrate unde bacteriile formeaza adevarate depozite vascoase.
Capsula (statul mucos) Unele bacterii au proprietatea de a elabora in anumite conditii de mediu substante macromoleculare cu caracter vascos, gelatinos, mucos. Aceasta proprietate ar exista potential la toate bacteriile dar s-ar manifesta regulat doar la unele specii in timp ce la altele s-ar traduce prin prezenta unei formatiuni foarte organizate dar invizibila cu mijloacele uzuale de cercetare.
In functie de structura si raportul cu celula bacteriana substanta mucoida se poate prezenta sub urmatoarele forme:
- microcapsula cu o grosime de 0,2 m. Substanta mucoida alcatuieste un strat foarte fin in jurul celulei bacteriene. Nu poate fi obserata la microscop, dar e detectabila prin procese imunologice.
- macrocapsula cu o grosime ce depaseste 0,2 m. Se prezinta ca o formatiune morfologica distincta si care inveleste de jur imprejur fiecare celula sau pereche de celule.
- zooglea sau masa zoogleica in care stratul mucos leaga intre ele intr-o singura masa mai multe celule formand adevarate colonii mucilaginoase de bacterii
Prin coloratii uzuale (coloratie Gram) capsula nu se coloreaza, dar apare sub forma unui halou necolorat in jurul bacteriei respective. Fiind putin opaca la electroni capsula nu apare clar sau este invizibila chiar la microscopul elecronic.
Din punct de vedere chimic capsula si stratul mucos sunt de natura polizaharidica.
Originea capsulei nu e inca bine lamurita desi capacitatea de a elabora capsula sau stratul mucos constituie un caracter ereditar, determinat genetic, ea poate sa nu se manifeste atunci cand conditiile de mediu nu sunt adecvate sau poate fi pierduta prin mutatii.
Semnificatia biologica Capsula nu este o parte integranta a celulei bacteriene ci un produs inert ce rezulta din activitatea ei metabolica si care poate fi indepartat, celula bacteriana pastrandu-si viabilitatea. Protejeaza celula bacteriana de efectul nociv al desecarii (eliminarii apei) datorita proprietatii sale higroscopice si reprezinta un rezervor pentru stocarea substantelor nutritive sau a celor rezultate din metabolism.
La bacteriile patogene capsulate, virulenta acestora esta mare pentru om si animale.
Cilii.Flagelii sunt apendice filamentoase unice sau multiple la suprafata bacteriilor, reprezentand organite de locomotie caracteristice procariotelor.
Lungimea cililor variaza in functie de specie, dar an general este mai mare decat a celulei care ii poarta. Acestia sunt mai lungi la bacteriile batrane si la cele care traiesc in medii lichide, iar la aceeasi celula pot avea lungimi diferite in functie de varsta lor.
La unele bacterii (G. Vibrio) flagelii sunt acoperiti cu o teaca de invelis. Au forma cilindrica, pot fi observati la microscopul fotonic dar numai dupa o prealabila colorare specifica. Aranjamentul flagelilor pe suprafata celulei, precum si numarul lor ce variaza (1-100/cel) sunt caractere de specie.
Bacteriile care nu poarta flageli sunt imobile si se numesc atrihe (ex: cocii). Celelalte de numesc bacterii flagelate (ciliate) si pot avea un numar diferit de cili si diferiti si ca pozitie. Astfel bacilii se numesc:
bacterii monotrihe - au un singur cil (Vibrio cholerae)
bacterii amfitriche - prezinta cate un cil la ambele capete (Bacillus megaterium)
bacterii lofotriche - prezinta cate un smoc de cili la unul din capete (Spirillum marinum)
bacterii peritriche - cu cilii pe intreaga suprafata a corpului (Rhizobium leguminoserum)
Utrastructura flagelilor - prezinta 3 parti: corpuscul bazal, o articulatie = carlig si un filament sub forma unui helix rigid format din proteine.
Corpusculul bazal este portiunea cea mai importanta pentru ca functioneaza asemeni unui motor rotativ al carui ax de transmitere este conectat cu filamentul helical printr-un carlig care serveste ca o articulatie flexibila universala.
Articulatia flexibila face legatura dintre axul de transmisie al rotorului si filamentul flagelului.
Din punct de vedere chimic flagelii sunt alcatuiti din polipeptide (cel putin 11 tipuri de polipeptide) componentul principal al filamentului flagelar fiind flagelina = o proteina majora cu o compozitie in aminoacizi variabila de la o specie la alta.
Deplasarea celulei bacteriene se face prin invartirea flagelului in jurul axului asemanator unei elice propulsand bacteria inainte cu o viteza cuprinsa intre 20-30 m/sec ceea ce corespunde cu de 2-100 ori lungimea celulei. Din punct de vedere energetic, celula consuma pentru mobilitate ≈ 0,1% din totalul energiei consumata pentru crestere. In ce priveste energia necesara deplasarii, bacteriile mobile pot sa tranforme energia chimica data de curentul de protoni ce traverseaza membrana plasmatica, curent ce se transforma in lucru mecanic.
Bacteriile se mai pot deplasa si prin alunecare sau glisare, miscare ce are loc intr-o directie paralela cu axul longitudinal al celulei, deplasare ce se realizeaza doar pe suporturi umede (ex. Myxobacteria si Cianobacteria - alge albastre-verzi)
O alta modalitate este prin tactism; daca o suspensie bacteriana este supusa actiunii unor factori fizici sau chimici se observa o anumita orientare a deplasarii celulelor bacteriene, fenomen numti tactism. Daca substanta sau factorul este o substanta chimica avem de-a face cu chimiotactism. Daca factorul este lumina, fenomenul se numeste fototactism si se intalneste la bacteriile fototrofe.
Pilii (fimbriile) sunt apendici filamentosi prezenti in numar mare pe suprafata unor bacterii. Desi cei doi termeni sunt considerati ca fiind sinonimi, altii apreciaza ca exista diferente semnificative de structura, functie si determinism genetic intre pili si fimbrii (Zarnea 1983). Pilii si fimbriile au o serie de caracteristici comune. Au fost descrisi mai ales la speciile de bacili Gram negativi, mai rar la speciile Gram pozitive. Pe baza caracterelor morfologice, biochimice si imunologice a fost descrisa o mare varietate de pili in timp ce in raport cu functionalitatea lor, acestia se impart in: - pili de conjugare sau de sex.
pili comuni sau somatici
Pilii de conjugare sunt implicati in procesele de conjugare bacteriana. Sunt in numar de 1-10/celula si asigura legarea dintre celulele bacteriene mascule (F+) si celulele bacteriene femele (notate F-) si prin aceasta, pilii sunt organite esentiale pentru transferul ADN-ului cromozomial sau plasmidic in cursul conjugarii actionand fie ca o conducta, fie ca o banda transportoare. Acestia sunt intalniti la Escherichia coli - tulpina K12, Proteus mirabilis.
Pilii comuni sunt dispusi pericelular putand fi polari sau bipolari. Numarul lor variaza in functie de specie 1-1000/cel.
Semnificatia biologica a pililor Nu reprezinta structuri comune tuturor bacteriilor. Daca sunt indepartati mecanic, ei se refac cu usurinta. Prezenta lor la un numar mare de specii ce habiteaza in medii naturale indica faptul ca indeplinesc unele functii importante ca: initierea procesului de conjugare a bacterii (pilii de sex).
De asemenea, pilii mediaza aderenta unor specii bacteriene la hematii, inducand fenomenul de hemaglutinare.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 6535
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved