| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Catabolismul alcoolului etilic
Unele levuri (Debaromyces, Hansenula, Pichia) si unele bacterii metabolizeaza complet alcoolul etilic, pana la CO2 si apa. Reactia are loc in doua trepte: in prima treapta rezulta aldehida acetica.
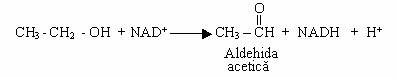
In treapta a II-a, aldehida acetica este oxidata la acetil-CoA si in conditii de aerobioza o incorporeaza in ciclul Krebs;
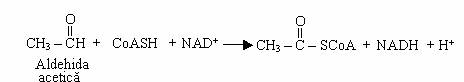
Bacteriile acetice (Acetobacter, Gluconobacter) oxideaza incomplet alcoolul etilic, cu acumularea acidului acetic, ca produs final, deoarece acetaldehida este oxidata direct la acid acetic:
![]()
Oxidarea acetaldehidei este o reactie aeroba si este baza producerii industriale a otetului alimentar, prin diferite metode. Dupa epuizarea alcoolului, unele bacterii acetice oxideaza acidul acetic la CO2 si H2O, prin intermediul acetil-CoA.
Bacteriile acetice sunt aerobe si se deosebesc de bacteriile acetogene anaerobe, prin aceea ca nu oxideaza complet sursa lor energetica. Oxidarea alcoolului etilic are loc numai pana la acid acetic, care se acumuleaza in mediu si pH scade. Bacteriile acetice sunt acido-tolerante.
Reactiile de anabolism
Reactiile de anabolism se desfasoara in sensul utilizarii metabolitilor intermediari ai cailor centrale, pentru sinteza constituientilor proprii celulei bacteriene. Procesele anabolice evolueaza in doua faze, care se desfasoara in sens invers in raport cu cele catabolice. Rezultatul lor este sinteza constituientilor celulari.
Celulele bacteriene sintetizeaza doua tipuri de macromolecule:
macromolecule informationale, codificate de mesaje genetice, cu caracter de specificitate, de importanta biologica fundamentala;
macromolecule de rezerva (de depozit), cu o structura in general uniforma, formate prin legarea unor monomeri, in polimeri de diferite marimi.
Sinteza macromoleculelor informationale se realizeaza cu o mare eficienta, deoarece, in procesele metabolice, intr-o faza initiala, nespecifica, sunt furnizate subunitatile de constructie: aminoacizi, baze purinice si pirimidinice. In cea de a II-a faza, controlata genetic se desfasoara procesele specifice de biosinteza a macromoleculelor informationale, care poarta denumirea generica de diataxie, in cursul careia subunitatile specifice sunt polimerizate intr-o ordine riguros exacta, conforma mesajului genetic.
Caile amfibolice. Sub aceasta denumire sunt reunite caile centrale ale metabolismului, care, simultan au rolul de a elibera energie si de a produce molecule precursoare pentru biosinteze.
Caracterul amfibolic este conferit de faptul ca energia si anumiti intermediari ai cailor catabolice sunt utilizati in reactii de anabolism, dupa urmatoarea schema generala:
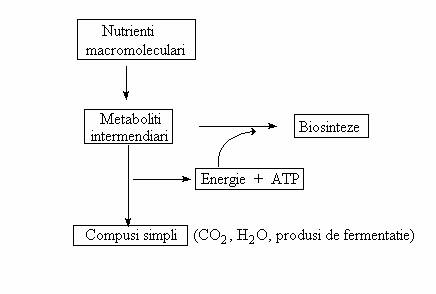
Existenta cailor amfibolice este expresia interactiunii dintre caile catabolice si anabolice, care functioneaza simultan in citoplasma necompartimentata a celulei bacteriene.
Caile anaplerotice (sau de aprovizionare) sunt cai metabolice auxiliare. Nevoia existentei lor deriva din faptul ca nutrientii din mediu sunt degradati progresiv pentru eliberarea energiei sau sunt folositi in reactiile de biosinteza. Calea metabolica principala ce furnizeaza metabolitii intermediari, esentiali pentru catabolism si anabolism trebuie reaprovizionata cu compusi care provin din alte cai metabolice, ce se desfasoara simultan in celula. Astfel, se asigura functionarea indelungata si la o rata optima a caii metabolice principale.
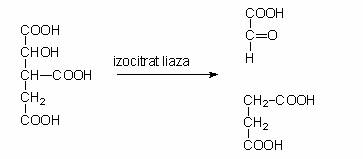
Calea anaplerotica a glioxilatului, are ca rezultat final, regenerarea oxaloacetatului, molecula acceptoare a acetatului in ciclul Krebs. Daca oxaloacetatul este intens folosit in biosinteze, functionarea ciclului acizilor tricarboxilici va diminua. Pentru a evita scaderea ratei de producere a energiei, se sintetizeaza suplimentar un compus cu 4 atomi de C. Daca substratul energetic este un glucid, compusul cu 4 atomi de C se formeaza prin carboxilarea piruvatului. Daca substratul energetic este acetatul sau un acid gras, microorganismele aerobe nu reduc acetatul la piruvat. Acetatul intra in ciclul Krebs, dar izocitratul nu este decarboxilat succesiv prin cele doua trepte, pana la succinat. In celula bacteriana este indusa sinteza a doua enzime care sunteaza cele doua reactii de decarboxilare intre izocitrat si succinat.
Izocitratul este scindat de izocitrat-liaza si rezulta glioxilat si succinat. Glioxilatul se combina cu o alta molecula de acetat din acetil-CoA si rezulta malat, care restabileste ciclul. Procesul ciclic in cursul caruia acetatul nu este convertit la CO2 poarta denumirea de ciclul glioxalic.
Bacteriile si plantele sintetizeaza acetil-CoA din acetat si coenzima A, intr-o reactie consumatoare de ATP, catalizata de ATP-sintetaza.
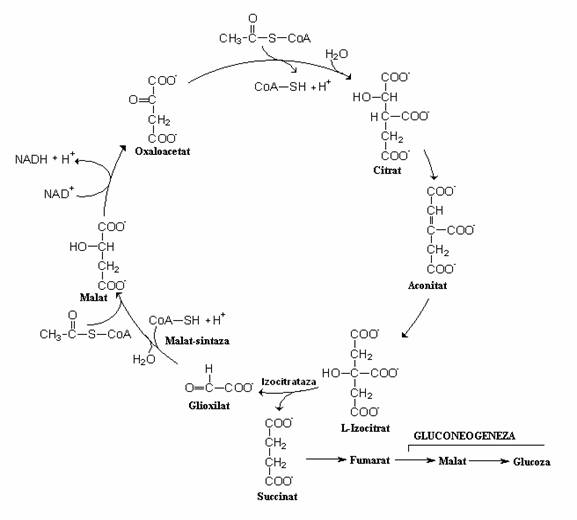
Fig. 73. Suntul glioxilatului si raporturile sale cu reactiile ciclului acizilor tricarboxilici. Reactia esentiala a ciclului este clivarea izocitratului in glioxilat si succinat. Malataul se sintetiteaza din glioxilat si acetil-CoA. Celelalte reactii ale ciclului sunt aceleasi ca si in ciclul Krebs. Ciclului glioxilatului este activ cand substratul energetic este catabolizat la acetat si nu functioneaza cand sunt degradate glucidele, deoarece sunt catabolizate la acid piruvic.
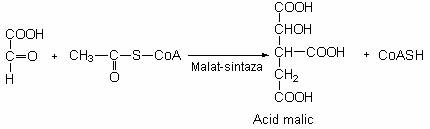
Intr-o a II-a reactie, acidul glioxilic este condensat cu acetil-CoA si formeaza acidul malic.
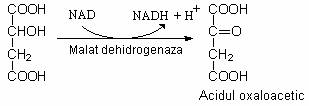
Acidul malic este oxidat si rezulta acidul oxaloacetic:
In concluzie, reactiile metabolice ale celulelor bacteriene, ca mecanism general de desfasurare sunt similare cu cele care au loc in celelalte sisteme vii, dar se deosebesc prin faptul ca, in special caile catabolice sunt mai diversificate, ca o consecinta directa a naturii foarte heterogene a substraturilor degradate.
Faptul ca in privinta mecanismului general de desfasurare, caile metabolice sunt similare cu ale celorlalte organisme a permis ca numeroase desoperiri ale biologiei moleculare sa se faca pe sistemul bacterian si sa se extrapoleze la celula eucariota:
ciclul Krebs
diferitele cai metabolice
procesele de oxido-reducere
mecanismul sintezei proteinelor
Din acest punct de vedere, culturile bacteriene au constituit un model ideal de investigare. Ele au avantajul cresterii rapide pe medii sintetice (cu compozitie bine determinata). A devenit astfel posibila, determinarea precisa a modificarilor chimice ale mediului de crestere, prin evidentierea diminuarii cantitative a unor componente si acumularea produselor de catabolism sau prin incorporarea precursorilor marcati ai mediului nutritiv, in macromolecule.
Deosebirea esentiala consta in faptul ca la bacterii functioneaza cai catabolice particulare, care nu se regasesc la alte organisme si care permit celulelor bacteriene sa catabolizeze substante chimice greu degradabile: cauciuc, asfalt, diferiti compusi aromatici etc.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2401
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved