| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Evolutia si raspandirea vietuitoarelor pe Glob
1 . Evolutia plantelor si animalelor in timpul erelor geologice
Evolutia biologica este un proces universal si propriu lumii vii, care consta, in esenta, in selectia naturala a mutatiilor adaptative ce asigura supravietuirea populatiilor prin adaptari noi la conditiile schimbate de mediu. Speciile care nu se adapteaza pier, lasand uneori urme fosile; deci sensul evolutiei este autoperfectionarea adaptativa a speciilor, in confruntarea cu mediul ambiant.
Evolutia este un proces creativ deoarece selectia promoveaza caracterele noi, aparute prin adaptarea la noi conditii de mediu si cu cat aceste conditii sunt mai variabile cu atat divergenta speciilor este mai evidenta; din contra, un mediu uniform timp indelungat duce la o evolutie convergenta a organelor analoage.
Evolutia este, de asemenea, un proces autoreglabil, de tip cibernetic, prin care se pastreaza un echilibru dinamic intre integralitatea specifica a populatiilor mutante si factorii de mediu care actioneaza determinand selectia naturala. Rezultatele evolutiei pot fi interpretate numai statistic si exprimate prin arbori filogenetici ale caror ramuri au valoare de vectori ai directiilor de evolutie. Taxonomia este disciplina care reconstituie si reprezinta sintetic si sistematic, evolutia plantelor sau a animalelor, clasificarea exprimand directiile evolutiei (divergente sau convergente), gradul de inrudire (prin rangul taxonomic), nodurile filogenetice, durata (in timp), viteza evolutiei (prin numarul de taxoni produsi) etc.
Evolutia este un proces continuu, dar cu ritmuri si viteze diferite la diverse grupe de organisme. Astfel, algele (cca. 35.000 specii) s-au diversificat timp de peste 600 milioane de ani, iar muschii (cca. 26.000 specii) si ferigile (cca. 12.000 specii) in numai 400 milioane de ani; gimnospennele (cca. 10.000 specii din care astazi mai traiesc doar 800 specii), desi au necesitat numai 200 milioane de ani de evolutie, predominand in mezozoic, sunt astazi in declin, in timp ce angiospermele (cca. 250.000 specii) aparute acum 150 milioane de ani, sunt astazi in plina expansiune si diversificare (D. Mititelu, 1981).
Conceptia moderna despre filogenia plantelor se bazeaza pe teoria sintetica a evolutiei care a completat ideile centrale ale darwinismului despre evolutia prin adaptare si selectie naturala cu teoria cibernetica a sistemelor autoreglabile ce caracterizeaza viziunea moderna asupra evolutiei biologice.
1 .1 Etapele principale ale evolutiei plantelor.
Cronologia stratigrafica paleobotanica nu se suprapune exact peste cea paleozoica deoarece erele caracterizate prin anumite grupe de plante, respectiv paleofitic, mezofitic si neofitic preced si se termina mai repede decat erele animalelor: paleozoic, mezozoic si neozoic.
1. Era primara a durat cca 400 milioane de ani, dar primele bacterii au aparut acum 3 miliarde de ani in urma, cand incepe epoca primelor arhebionte celulare procariote (cu nucleu difuz) si asexuate, numita Proterozoic (Tab. I).
Epoca algelor dureaza din Precambrian pana in Devonian (peste un miliard de ani), viata desfasurandu-se aproape numai in apa (pe uscat traiau putine bacterii, alge, ciuperci). In aceasta epoca s-au realizat mai multe etape de dezvoltare: celula eucariota (cu nucleu diferentiat), stadiul cenobial si talul pluricelular, pigmenti asimilatori (clorofilele) si acumularea de amidon, reproducerea sexuata.
Epoca ferigilor dureaza din Devonian pana in Permian (cca. 130 milioane de ani) si incepe cu aparitia florei terestre, prin transformarea algelor litorale in psilofite si a acestora in celelalte grupe de ferigi terestre. In Devonian psilofitele palustre reprezentau practic intreaga vegetatie, iar la sfarsitul epocii, in Carbonifer, ferigile arborescente dominau intreg Globul. Tot in aceasta epoca apar primii muschi cu organe de inmultire specializate. Se dezvolta cormul format din radacina, tulpina si frunze, cu tesuturi conducatoare specializate, tipice plantelor terestre.
2. Era secundara ce dureaza aproximativ 250 milioane de ani este, in cronologia paleobotanica, epoca gimnospermelor care incepe in Permianul superior si tine pana in Cretacic. Climatul mai secetos determina reducerea severa a ferigilor lemnoase higrofile si expansiunea gimnospermelor mai xerofile care ocupa intreg landsaftul. Perfectionarea organelor de reproducere, respectiv aparitia florii-con in care ovulul devine samanta, determina expansiunea considerabila mai ales in emisfera nordica a gimnospermelor. Emisfera sudica dominata de un climat tropical favoriza inca persistenta ferigilor arborescente.
3. Era Tertiara incepe acum 70 milioane de ani in urma si este marcata de expansiunea angiospermelor, aparute in Cretacic, probabil din cele mai primitive gimnosperme. Angiospermele sunt cele mai evoluate plante de uscat ale caror caractere de superioritate fata de gimnosperme (o protejare mai buna a semintei in ovarul ce devine la maturitate fruct) le permit o expansiune rapida pe tot uscatul. Fiind eel mai bine adaptate la viata de uscat s-au diversificat realizind cca. 250.000 specii, cele mai multe genuri formandu-se inca din Tertiar si migrand in toate continentele datorita legaturilor temporare sau permanente dintre acestea. Angiospermele au urmat mai multe directii evolutive, constituind mai multe subclase cuprinse in doua clase:
I. Clasa Magnoliatae (dicotiledonate):
- Magnoliidae - angiosperme primitive policarpice (mai multe carpele), lemnoase si ierboase, aparute in Jurasic si cunoscute ca fosile in Cretacicul inferior din care se presupune ca ar fi derivat celelalte grupe de angiosperme; se clasifica in urmatoarele ordine: Magnoliales, Piperales, Ranunculales, Aristolochiales, Nymphaeales, Papaverales;
- Ranunculidae - plante ierboase asemanatoare magnoliidelor; consistenta ierboasa, lipsa radacinii principale si a pungilor secretoare de uleiuri eterice, aparitia zigomorfiei etc. sunt caractere de superioritate fata de magnoliide; familii mai importante: Ranunculaceae si Berberidaceae;
- Hamamelide - plante lemnoase cu multe caractere primitive; majoritatea au flori unisexuate grupate in amenti (amentifere); se clasifica in 7 ordine; familii mai importante Platanaceae, Ulmaceae, Moraceae, Canabaceae, Urticaceae, Fagaceae, Betulaceae, Juglandaceae;
- Caryophyllidae - plante ierboase, rar lemnoase raspandite pe tot Globul ce prezinta asemanari cu reprezentanti din dileniide, rozide si chiar ranunculide; ovule putine (deci seminte putine) prinse central (centrosperme); familii mai importante: Caryophyllaceae, Aizoaceae, Chenopodiaceae, Polygonaceae etc.;
- Dileniidae - plante lemnoase si ierboase din regiunile tropicale si subtropicale, cu numeroase ovule dispuse parietal; cuprinde 10 ordine; familii mai importante: Paeoniaceae, Theaceae, Hypericaeae, Violaceae, Brassicaeae, Salicaceae, Cucurbitaceae, Malvaceae, Tiliaceae, Bombacaceae, Sterculiaceae, Ericaceae, Primulaceae etc.;
- Rosidae - dicotiledonatele cele mai tipice cu flori bisexuate dialipetale; se clasificii in 20 de ordine; familii mai importante: Crassulaceae, Saxifragaceae, Droseraceae, Rosaceae, Fabaceae, Mimosaceae, Rizophoraceae, Sonneratiaceae, Onagraceae, Myrtaceae, Onagraceae, Lytraceae, Anacardiaceae, Aceraceae, Geraniaceae Araliaceae, Apiaceae, Vitaceae, Santalaceae, Rafilesiaceae, Euphorbiaceae.
- Asteridae - constituie una din cele mai evoluate linii filogenetice (gamopetale tetraciclice) din clasa Magnoliatae; sunt plante lemnoase si ierboase riispandite pe tot globul; se clasificii in 8 ordine; familii mai importante: Gentianaceae, Rubiaceae, Caprifoliaceae, Dipsacaceae, Oleaceae, Convolvulaceae, Solanaceae, Scrophulariaceae, Plantaginaceae, Lamiaceae, Asteraceae etc.;
2. Clasa Liliatae (monocotiledonate)
- Alismidae - trimere hidrogame cu carpelele libere; familii mai importante: Alismaceae, Potamogetonaceae;
- Liliidae - trimere entomogame; familii mai importante: Liliaceae, Agavaceae, Iridaceae, Orchidaceae, Juncaceae, Czperaceae, Bromeliaceae, Poaceae;
- Arecidae - plante cu inflorescenta sub formii de spadice (spadiciflore); familii: Arecaceae (Palmae), Araceae, Lemnaceae, Typhaceae, Sparganiaceae.
In Fig. 2. se prezinta o cheie simpla de determinare a lumii vegetale, angiospermele fiind clasificate pe un criteriu legat de morfologia florii (respectiv, flori tarii petale, flori cu petale1e libere, flori cu petalele sudate).
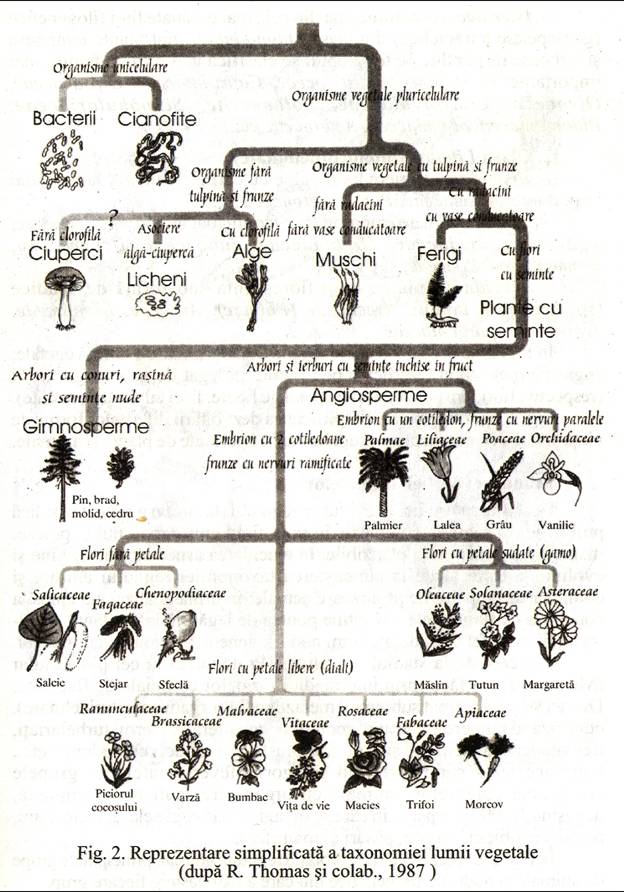
In Fig. 3. se prezinta istoria simplificata a dezvoltarii diferitelor forme de viata si releva sincronizarea grupelor mai importante de plante si animale.
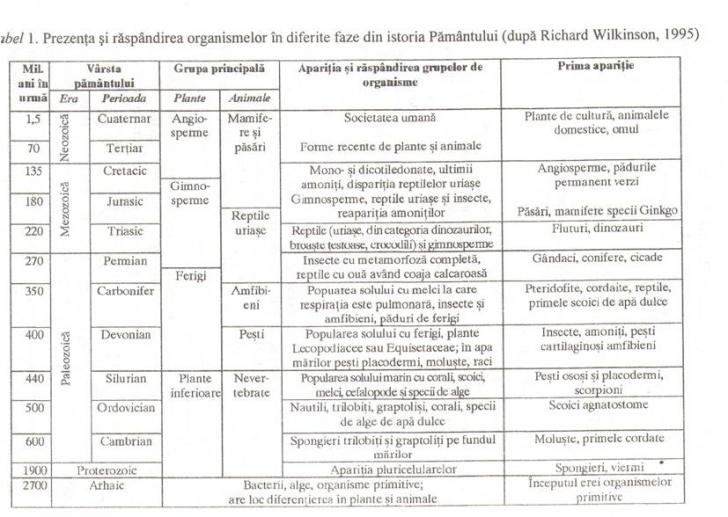 fig. 3
fig. 3
Etapele evolutiei animalelor
Descifrarea originii si evolutiei regnului Animalia a ridicat si ridica probleme deosebit de complexe, in timp fiind emise mai multe ipoteze, mai mult sau mai putin plauzibile. In elucidarea aspectelor de origine si evolutie s-a plecat de la cunoasterea taxonomiei regnului animal si compararea grupelor de protozoare actuale; in urma analizei s-a ajuns la concluzia ca flagelatele constituie puntea de legatura intre plante si animale, fiind astfel considerate prim nod filogenetic in evolutia animalelor.
Trecerea de la stadiul unicelular (Protozoare) la cel pluricelular (Metazoare) s-a facut prin intermediul formelor coloniale de flagelate. Din acestea s-a format subregnul metazoarelor (organisme pluricelulare), cuprinzand numeroase grupe: spongieri, celenterate, viermi turbelariati, trematode, nematelminti si anelizi, moluste, artropode, echinoderme etc., toate acestea facand parte din categoria nevertebratelor si grupele apartinand cordatelor, animale cu organizare mult mai complexa, depasind 50.000 de specii, in care se includ urmatoarele clase: ciclostomi, pestii, amfibieni, reptile, pasari si mamifere.
Cordatele inferioare reprezinta animale putin mobile, care se hranesc in mare masura pasiv; stramosii lor au dat nastere la forme capabile sa se deplaseze activ 'asemanatoare pestilor', cunoscute sub denumirea de Ostracodermi, considerati ca un ram inrudit cu primele vertebrate si gasite in strate siluriene si devoniene. Ostracodermii au disparut nelasand descendenti.
Stramosii directi ai pestilor sunt necunoscuti deocamdata, desi urme de pesti placodermi s-au gasit in Silurianul superior si Carbonifer, iar de pesti cartilaginosi si ososi in Devonian. Stramosii Crossopterigienilor si ai Dipnoilor au aparut in Devonian si au ajuns la dezvoltarea maxima in Carbonifer; primii prezinta un interes special prin aceea ca dintre toti pestii, ei stau eel mai aproape de grupa de origine a vertebrate lor terestre, Stegocefal
Amfibienii prezinta un interes deosebit pentru evolutia animalelor deoarece la nivelul lor vertebratele au trecut de la viata in mediul acvatic la cea pe uscat, fapt insotit de doua adaptari hotaratoare: respiratia oxigenului atmosferic si locomotia pe substrat tare. Amfibienii fosili sunt cunoscuti din straturile din Devonianul superior din Groenlanda. In strate din perioadele Carbonifer si Permian s-au descoperit resturi de labirintodonti, cuprinzand cei mai mari amfibieni dispamti (de marimea crocodililor actuali). Amfibienii disparuti sunt reuniti in subclasa Stegocefalilor, ce aveau o cuirasa osoasa continua ce le acoperea capul; acestia au trait pana la inceputul Mezozoicului, iar ordinele actuale de amfibieni au aparut abia la sfarsitul Mezozoicului, din care cauza nu a putut fi stabilita o legatura directa intre ele. Conditiile din ultima perioada a Carboniferului, cand clima era umeda, calda si uniforma pe cea mai mare parte a continentului, probabil unic, au fost favorabile Stegocefalilor.
Deplasarile continentelor fata de poli, formarea un or lanturi muntoase, insotite de schimbarea climei si a vegetatiei au dus la reducerea mlastinilor si intinderilor umede si aparitia zone lor uscate cu vegetatie xerofila, favorabile reptile lor care au evoluat foarte rapid, extinzandu-se in cele mai diferite conditii de mediu, unele readaptiindu-se vietii acvatice, iar altele devenind aeriene. Adaptarea divergenta a reptilelor a fost uimitoare. Pentru majoritatea grupelor era caracteristica o mare mobilitate, scheletul a devenit mai usor, structura membrelor, a trunchiului si a craniului au suferit modificari esentiale, adaptiindu-se pentru a folosi o hrana tot mai variata. De aceea, Mezozoicul a fost denumit 'era Reptilelor'. Dintre reptilele cele mai primitive, Rincocefalii aparuti in Triasic, Sphenodon (Hatteria) punctatum s-a pastrat, in Noua Zeelanda, pana in zilele noastre. Crocodilienii au aparut la sfarsitul Triasicului: majoritatea speciilor erau de apa dulce si mai putine specii marine. Pterozaurienii erau reptile zburatoare de cativa centimetri pana la un metru si mai bine; maximul lor de dezvoltare a fost in Jurasic. Dinozaurienii au fost grupa cea mai numeroasa si mai diversa de reptile, dimensiunile lor variind intre 1 si 30 m lungime; ei au aparut in Triasic si au ajuns la apogeu in Cretacic, spre sfarsitul perioadei disparand. Lacertilienii au avut 0 dezvoltare mai mare in Cretacic, iar ofidienii au aparut mai tarziu decat celelalte reptile (in Cretacic) din lacertilieni. Chelonienii sunt un grup foarte vechi care a aparut in Permian, in Mezozoic erau probabil animale sapatoare de uscat, care ulterior s-au adaptat la viata acvatica. Din Triasic pana in zilele noastre si-au pastrat toate trasaturile esentiale ale organizarii lor.
La sfarsitul mezozoicului marea majoritate a grupelor sistematice mari (subclase) disparusera, probabil datorita specializarii lor unilaterale la conditii de viatii specifice si strict determinate. Modificarile insemnate de clima si de peisaje geografice, datorita deplasarilor litosferei si formarii muntilor din 'epoca alpina de formare a muntilor' s-au reflectat in evolutia covorului vegetal prin inlocuirea florei mezozoice de Cicadale si Conifere cu plante de tip nou, Angiospermele; toate aceste modificari au avut repercusiuni asupra tuturor grupelor de animale, si in primul rand asupra celor strict specializate. 'Reptilele actuale reprezinta doar ramasite deplorabile ale faunei din Mezozoic' (S.P.Naumov, 1954).
Pasarile isi au originea in reptilele vechi (grupa Pseudosuchiilor), foarte specializate, adaptate la catarat, ulterior dezvoltandu-si capacitatea de a sari de la un arbore la altul. Scheletele relativ complete a doua tipuri: Archaeopterix si Archaeornix, descoperite in strate jurasice, releva un amestec de trasaturi evidente de pasari si de reptile, modul lor de viata fiind arboricol, puteau sa planeze, dar nu sa zboare. Cu timpul, zborul planat a dus la aparitia zborului propriu-zis si, de asemenea, la desavarsirea altor trasaturi: aparitia penajului a dus la realizarea homeotermiei; separarea sangelui arterial de cel venos si complicarea aparatului respirator au dus la intensificarea metabolismului; dezvoltarea sistemului nervos a dus la perfectionarea comportamentelor de procurarea hranei, de apararea de dusmani, de cresterea puilor. Zborul le-a permis ca in scurt timp sa populeze intregul Glob pamantesc, ocupand cele mai variate conditii de mediu.
Mamiferele s-au desprins in Paleozoic din Reptile primitive care nu aveau o specializare stricta, astfel incat sub actiunea factorilor externi s-au produs adaptari hotaratoare, respectiv evolutia progresiva a Mamiferelor, care consta in: temperatura ridicata a corpului si capacitatea de termoreglare (atat in conditii de temperaturi scazute, cat si in conditii de temperaturi ridicate), viviparitatea (nasterea de pui vii si hranirea lor cu lapte) si, in special, activitatea nervoasa foarte dezvoltata, care asigura dezvoltarea unui comportament complex in sensul unei mai bune adaptari la mediu.
Mamiferele au aparut probabil in Jurasic sau chiar in Triasic, materialul privitor la primele Mamifere mezozoice limitandu-se la dinti izolati, la maxilare sau la fragmente de craniu. Marsupialele apar in perioada cretacica, la inceputul Tertiarului fiind raspandite pe tot globul.
Ulterior Placentarele mai perfectionate au inlaturat sau au distrus direct Marsupialele. Separarea Australiei de celelalte continente inainte de patrunderea Placentarelor a dus la supravietuirea acestora pana in zilele noastre, pe de o parte, si la diversificare maxima a formelor de marsupiale care au ocupat toate mediile. In celelalte continente, Marsupialele au fost eliminate aproape in intregime de Placentarele in expansiune.
Grupa cea mai veche de Placentare o constituie Insectivorele (identificate in Mongolia in strate din Creatacicul superior), din care au luat nastere Chiropterele, Edentatele si Rozatoarele. Cu Insectivorele se inrudesc Carnivorele (care apar pe la inceputul Tertiarului - Creodontele) ce au dat ulterior Carnivorele actuale terestre, Pinipedele si Cetaceele. Tot din Insectivore au luat probabil nastere si Ungulatele sau Copitatele.
Maimutele fosile se cunosc din Oligocen: cele de tipul Propliopithecus au dat nastere Hilobatidelor (gibonilor) si speciei Ramapithecus din Miocenul Indiei, apropiata de Antropoide. Un mare interes prezinta Australopithecus si mai ales maimutele antropoide gasite in depozitele cuatemare din Africa de Sud.
Specia cea mai primitiva de oameni stravechi - Pithecanthropus - s-a gasit in depozitele pleistocene pe insula Java, in anul 1891; resturi ale unui alt reprezentant al oamenilor stravechi, Sinanthropus, au fost gasite in anul 1927 in apropiere de Pekin; resturi fosile ale omului de Neanderthal (Homo primigenius), s-au descoperit in mai multe locuri din Eurasia. Aceasta specie a existat in perioada cuatemara, pe la inceputul ultimei epoci glaciare. La starsitul ultimei glaciatiuni, exista specia actuala de Homo sapiens.
2. Arealul geografic
1. Definitie. Majoritatea speciilor de plante si animale traiesc intr-un spatiu geografic limitat - teritoriu sau acvatoriu, daca este vorba de organisme marine - ce poarta numele de areal. Arealul se refera numai la teritoriul normal de trai al speciei si nu la teritorii in care specia se poate gasi ocazional. Exemple: rogozul arctic (Carex lapponica), murul arctic (Rubus chamaemorus) si vulpea polara (Alopex lagopus), iepurele arctic (Lepus timidus) si foca (Phoca vitulina) - pentru mediul acvatic - ocupa arealul circumpolar boreal; garofita (Dianthus cal/izonus) ocupa un areal restrans in Piatra Craiului, singurul loc din lume unde aceasta vegeteaza.
Migratiile obligatorii la unele specii de pasari si pesti au dat nastere la discutii, deoarece unele dintre aceste animale au areale de reproducere si de iemat diferite, iar altele au areale de hranire, de reproducere, de iemat si de migratie, diferite. Raspandirea geografica a acestor specii se reprezinta prin arealul propriu-zis (de reproducere), cel de iernat, cel de hranire, caile de migratie si chiar teritoriile in care specia se poate gasi ocazional (daca nu a fost adusa de om). Sunt situatii in care arealul natural al unor specii nu poate fi stabilit decat cu greu deoarece introducerea lor in teritorii noi s-a facut cu sute de ani in urma. Aceste specii numite adventive, plante ruderale (buruieni) sau animale sinantrope, transportate voit sau accidental de om, 'incurca' biogeografia, deoarece nu intotdeauna se poate stabili arealul lor natural.
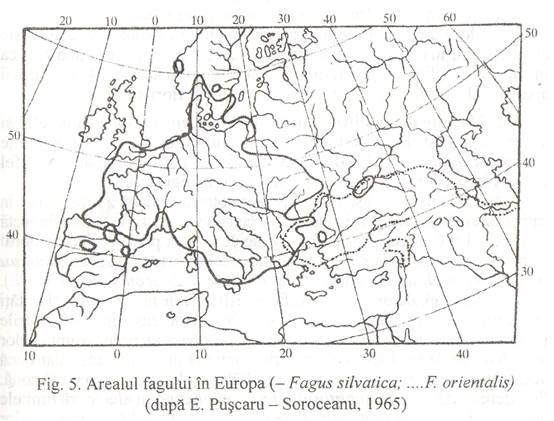
Daca se studiaza ariile de raspandire ale speciilor se constata ca ele sunt foarte deosebite (Fig. 5.) si numai rareori se suprapun ariile de la doua sau mai multe spec
Toate speciile ale caror areale se incadreaza intr-un tip de areal, apartin aceluiasi element floristic sau fitogeografic, acesta reflectand anumite conditii fizico-geografice. Numele acestuia corespunde cu denumirea arealului respectiv.
Principalele elemente floristice din Europa sunt urmatoarele:
1. elementele europene - cu raspandire in Europa Centrala si nordica;
2. elementele continentale sau estice (irano-turaniene) - cantonate in partea de sud-est a Europei si sudul Asiei;
3. elementele arctice si alpine - din extremul nordic al Europei si de pe muntii inalti;
4. elementele balcanice sau sud-estice - caracteristice Peninsulei Balcanice;
5. elementele mediteraneene sau sudice - localizate in bazinul mediteraneean;
6. elementele atlantice sau vestice - raspandite in tarile marginite de Oceanul Atlantic;
7. elementele cosmopolite - cu o raspandire larga pe tot globul;
8. elementele adventive - introduse din alte continente si care se raspandesc mult in noile regiuni;
9. elementele endemice - cu un areal foarte redus.
Flora unei anumite regiuni, ca si a unei asociatii vegetale, este alcatuita din mai multe elemente floristice. Astfel, in tara noastra, ca urmare a conditiilor pedoclimatice foarte diferite, se gasesc specii apartinand la numeroase categorii de elemente floristice.
2. Metode de stabilire a arealului. Studiul structurii arealelor si allegilor de distributie si mobilitate a populatiilor biologice constituie obiectul arealografiei, hartile de raspandire a taxonilor ramanand astfel cel mai important instrument de lucru al biogeografului.
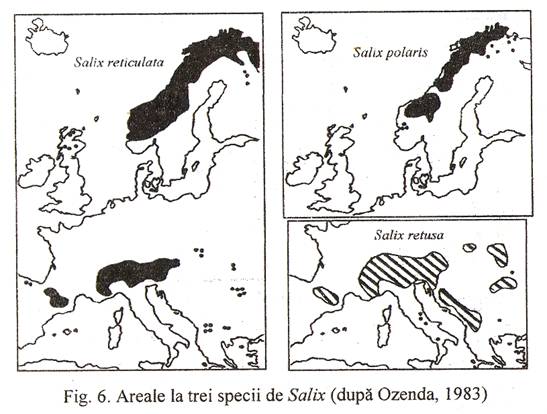
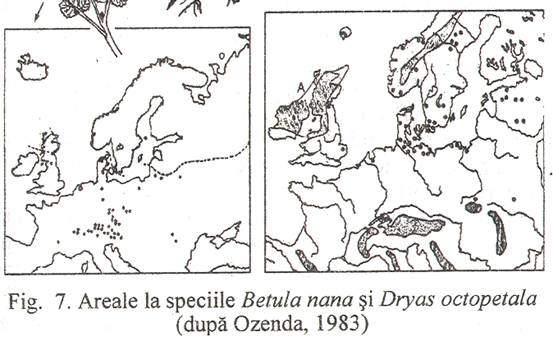
Arealul unei specii se stabileste prin notarea pe hartii a localitatilor in care aceasta a fost identificata si unirea punctelor extreme; zona astfel delimitata este arealul speciei respective. Functie de densitatea punctelor in interiorul arealului si configuratia spatiului geografic, arealul poate fi intreg sau continuu (Fig. 5) discontinuu sau disjunct (Fig. 6) si uneori chiar punctiform (Fig. 7.).
In cazul arealelor continui se face abstractie de micile discontinuitati (exemplu: in arealul iepurelui se includ si zonele intens populate, lacurile si raurile, chiar dada in acestea el nu traieste), dupa cum, in cazul animalelor de apa dulce, arealul cuprinde o parte continua de continent, chiar daca aceste animale populeaza numai raurile, baltile si lacurile din aceastii zona. Se diferentiaza astfel discontinuitati ecologice care nu afecteaza limitele arealului si discontinuitati corologice care se reflecta in delimitarea zone lor de raspandire a speciei si ridica probleme biogeografului.
3. Marimea arealelor variaza foarte mult, speciile cosmopolite sau aproape cosmopolite fiind raspandite pe tot Globul, iar cele endemice avand un areal foarte limitat (un masiv muntos, o portiune dintr-o insula, un bazin acvatic).
Marimea arealelor este determinata de mai multi factori:
- vechimea sau istoria geologica a speciei - in general, cu cat o specie are o vechime mai mare, cu atat arealul acesteia este mai extins, in timp ce speciile tinere au areale restranse; aceasta nu este o regula generala deoarece schimbarile climatice majore care au avut loc in istoria Globului au dus la restringerea severa a arealului multor specii si chiar la suprimarea multor taxoni;
- valenta ecologica a speciilor - speciile cu valenta ecologica mare, care pot trai in conditii de viatii a caror amplitudine de variatie este mare, au un areal mai extins decat cele specializate strict la anumite conditii;
- posibilitatile de raspandire ale speciei (pasive sau active);
- intamplarea.
In linii generale, speciile marine au cele mai extinse areale, dupa care urmeaza speciile terestre; cele dulcicole au de regula areale mai mici (exceptie fac speciile cu mari posibilitati de raspandire pasiva), iar cele mai mici arealele au speciile cavemicole.
In cadrul arealelor mari se diferentiaza cateva tipuri:
- areale zonale care se prezinta sub forma de benzi latitudinale, suprapuse aproximativ zonelor climatice; exemplu: areal circumpolar, areal pantropical etc.;
- areale meridiane care se prezinta sub forma de fasii dispuse pe directia nord-sud - in America de Sud numeroase specii de plante si animale au arealul aproape paralel cu coastele vestice;
- areale regionale care ocupa portiuni mai restranse sau mai intinse dintr-un continent.
Speciile cosmopolite sunt raspandite pe tot Globul. Mentionam dintre cosmopolite:
- animale: starcul cenusiu (Ardea cinerea), striga (Tyto alba), lepidoptere din genul Vanessa, unele animale marine pelagice;
- plante acvatice si palustre: lintita de balta (Lemna), broscarita (Potamogeton), trestia (Phragmites australis), papura (Typha latifolia, T angustifolia);
- plante terestre: urzica (Urtica dioica), papadia (Taraxacum officinplis), troscotul (Polygonum aviculare), patlagina (Plantago major).
In cazul unitatilor endemice, cand limitarea arealului se datoreaza factorului timp, se mentioneaza prin prefix tipul de endemism, respectiv, paleo- sau neo-endemism.
Paleoendemismele sunt unitati taxonomice de plante sau animale cu origine straveche care au reuisit sa supravietuiasca tuturor schimbarilor petrecute in timp, arealul lor restrangandu-se (in general), astfel incat muIte dintre acestea sunt pe cale de disparitie. Mentionam cateva specii de plante paleoendemice: Welwitschia mirabilis de varsta mezozoica, din desertul Namib, considerata adevarata fosila vie; arborele pagodelor (Ginkgo biloba), raspandit in Cretacic, cu un areal redus astazi in China; arborele rosu (Sequoia sempervirens) si arborele mamut (Sequoia gigantea) - gimnosperme care in Cretacic ocupau un intins areal circumpolar boreal, astazi au o raspandire redusa in vestul S.U.A.
Dintre animalele paleoendemice mentionam: pestele marin: Latimeria chalumnae din Oceanul Indian, raspandit in Paleozoic, pestii Dipnoi (Neoceratodus forsteri) din America de Sud, si, de la noi din tara, endemismele tetiare (termofile): peistele Scardininus racovitzi si melcul Melanopsis parreyssi din apele lacului Petea (Oradea). Uneori arealul unor paleoendemisme poate fi foarte intins (marsupialele-ordinul Monotrema) sunt raspandite in Australia, Tasmania si o parte din Noua Guinee), sau dimpotriva, restrans - (gimnospermul Welwitschia mirabilis vegeteaza numai in desertul Namib).
Neoendemismele sau endemismele progresive sunt specii aparute recent (Pliocen sau Cuatemar) prin izolare geografica sau specializare stricta la anumite conditii de mediu, cu tendinta de extindere.
Se apreciaza ca tara noastra este una dintre cele mai bogate tari din Europa in specii endemice (aprox. 160 taxoni) si aceasta se explica prin conditiile pedoclimatice foarte variate, prezenta unei putemice catene muntoase cu substrat geologic foarte diferit, pozitia geografica a tarii la o adevarata rascruce unde se intanesc mai multe tipuri de flora. Majoritatea endemismelor se gasesc in pajistile alpine si etajul jneapanului si au fost denumite endemisme carpatice.
Aparitia de specii noi - in cadrul aceluiasi gen! -, ca urmare a diversitatii mari a conditiilor de mediu, poarta denumirea de vicariere, iar speciile inrudite, putin diferite intre ele se numesc specii vicariante. Exemple: Fagus silvatica, este raspandit in centrul si vestul Europei, iar Fagus orientalis, in sud-estul si estul continentului.
Unitatile taxonomice relicte au supravietuit erelor geologice si ocupa areale restranse ca urmare a schimbarii in timp a conditiilor climatice. Cercetarile fitogeografice au evidentiat prezenta in flora tarii noastre a numeroase relicte tertiare si glaciare. Dintre relictele tertiare (care au trait inainte de glaciatie) amintim: Nymphaea lotus var. termalis (nufar sau drete) - Baile Felix, Oradea, Syringa josikaea (liliac ardelenesc) cu areal punctiform in Carpati, Hepatica transsilvanica (crucea voinicului) Carpatii estici si sudici. Unele specii au intregul areal fragmentat, ca urmare a schimbarilor, iar altele au un areal foarte mic, ramas din cel vechi.
Relictele glaciare sunt ramasite din timpul glaciatiunilor care traiesc si azi in unele enclave din zonele muntoase.
In zona tarii noastre populeaza mlastinile eutrofe (cca. 40 de specii): Betula humilis - Tusnadu Nou, Harghita, Viola epipsila - Bazinul Ciuc, Pedicularis sceptrum carolinum - Harman, Brasov si Zvoristea, Botosani, Carex loliacea - Dragoiasa, Suceava, Angelica palustris - Balanesti, Neamt, Calla palustris, Carex diandra, Sesleria coerulea, Ligularia sibirica si oligotrofe (cca. 20 specii: Scheuchzeria palustris - Lacul Manta, Buzau, Carex limosa, Carex paupercula, Carex pauciflora - Muntii Semenicului, Betula nana - Lucina, Suceava).
4. Raspandirea speciilor in cadrul arealului. In cadrul arealului unei specii frecventa indivizilor nu este uniforma, prezentand uneori discontinuitati mari datorita conditiilor ce pot varia foarte mult. Discontinuitati mai mici se intilnesc la speciile marine, mai mari la speciile care traiesc in mediul terestru si cele mai mari la speciile cavemicole, speciile de munte (cu cat altitudinea la care traiesc este mai mare, cu atat fragmentarea, discontinuitatea arealului este mai mare - exemplu: capra neagra are arealul fragmentat in sapte zone principale: Pirinei, Alpi, Apenini, Dinarici, Carpati, Caucaz si Taurus) si speciile acvatice dulcicole (cu cat specia este mai legata de zona superioara a raurilor, cu atat arealul este mai fragmentat - pastravul Salmo trutta fario traieste numai in apele de munte din tara noastra, fiind reprezentat prin peste 100 de populatii).
Se poate considera ca o regula generala faptul ca la majoritatea speciilor de plante si animale - exceptie endemismele strict localizate - arealul este fragmentat, chiar daca acesta apare, in reprezentarile cartografice, ca unitar.
5. Barierele sunt obstacole, intreruperi de natura geografica fiziografice si climatice - sau biologica (exemplu - dependenta unor genuri de plante de anumite specii de insecte polenizatoare) cu rol important in delimitarea arealului unei spec Barierele pot fi abrupte (un lant muntos foarte inalt, o zona de desert, un fluviu foarte lat sau un brat marin), marcand evident limita arealului, si discrete (temperatura aerului, gradul de umiditate atmosferica), care determina inrautatirea treptata a conditiilor de existenta a speciei (Fig. 8.). Astfel tantarul Aedes aegypti are o distributie tropicala si subtropicala limitata de izotermele de 10sC.
Barierele abrupte au o pozitie fixa in spatiu si permanenta in timp, iar cele discrete, in general de natura climatica, se deplaseaza de la nord la sud si invers, ca urmare a schimbarii climei.
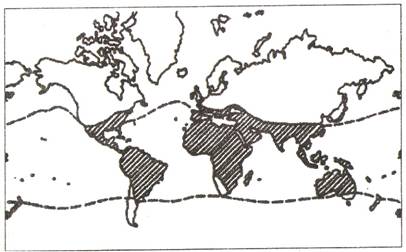
Fig. 8. Repartitia geografica a tantarului Aedes aegypti (dupa Ch.J.Krebs, 1971 din V. Simionescu, 1980)
6. Modificari ale arealelor. Desi in aparenta arealele sunt stabile, in decursul timpului geologic, acestea sunt supuse continuu unor modificari mai rapide sau mai lente, cu restrangerea sau extinderea teritoriului, ea urmare a schimbarilor climatice. In literatura sunt semnalate numeroase exemple de specii de plante si animale care intr-un timp foarte scurt si-au modificat arealul - extinderea buruienilor, restrangerea severa a arealelor unor animale de talie mare, expansiunea rapida spre nord si nord-est a speciilor mediteraneene, expansiunea spre vest si sud-vest a unor elemente siberiene. Daca unele modificari sunt o consecinta a interventiei omului - si, arealul "artificial" nu trebuie confundat cu cel ,,natural", - alte modificari insotesc de fapt schimbarile climatice actuale si evidentiaza tendinta generala a oricarei specii de a-si extinde arealul.
7. Migratiile pestilor si pasarilor ridica probleme deosebite biogeografiei, pe de o parte, in stabilirea arealelor speciilor in eauza si, pe de alta parte, in stabilirea originii anumitor grope (in cazul pestilor) sau a originii istorice a migratiilor (in cazul pasarilor).
Migratiile pestilor. Unele specii de pesti migreaza pe distante mai scurte sau mai lungi in mediul in care acestea traiesc in mod obisnuit, altele migreaza din mare in apele dulci. Astfel, unii pesti de origine mediteraneeana (scrumbia albastra pontica, palamida) ierneaza si se reproduc in Marea Marmara sau Marea Egee, iar vara traiesc in Marea Neagra unde gasesc suficienta hrana; aceste specii au nevoie in perioada de reproducere de salinitatea ridicata din arealul lor de origine dar nu suporta temperarile scazute de peste iarna din Marea Neagra.
Migratiile pestilor dulcicoli pe cursul aceluiasi fluviu nu au semnificatie zoogeografica deosebim. Cele mai importante migratii sunt cele ale pestilor care trec din mediul marin in cel dulcicol sau invers. Dintre pestii care trec din mare pentru a se reproduce in apele dulci (pesti anadromi) mentionam: somonul (urca din Marea Nordului pe fluviile din nordul si vestul Europei), scrumbia de Dunare urca, o populatie pe Dunare; alta pe Nistru si o alta pe Don, nisetrul, pastruga si morunul care urca pe Dunare. O alta categorie de migratori (pesti catadromi) traiesc si se hranesc in apele dulci si coboara pentru a se reproduce in mare. Exemplu: anghila (Anguilla anguilla) din fluviile Europei migreaza in Marea Sargasselor pentru a se reproduce; alte specii ale aceluisi gen din America de Nord, Japonia, sudul Australiei migreaza pe distante lungi pentru a ajunge in marile tropicale unde se reproduc; s-a tras concluzia ca originea lor este in marile tropicale.
Migratiile pasarilor au aparut ca o consecinta a schimbarilor severe ale conditiilor de viata (lipsa hranei, in primul rand) intr-un anumit sezon. Dovada este faptul ca pasarile din padurea ecuatoriala nu migreaza si ca, pe masura ce ne departam de ecuator creste proportia pasarilor migratoare si amploarea migratiilor. Pasarile migratoare au o zona (areal) de cuibarit si alta de iernat, situate la distante uneori foarte mari, pe care le strabat de doua ori pe an. Caile de migratie sunt constante pentru fiecare specie si chiar majoritatea speciilor aceleiasi regiuni au aceleasi cai de migratie. Astfel, pentru pasarile din vestul Europei, principala cale duce prin peninsula Iberica in Africa nord-vestica; pentru cele din est, in lungul coastelor Marii Negre, prin Asia Mica, Siria, apoi pe valea Nilului spre centrul Africii, evitand Sahara. Alta ruta duce prin Italia in Sicilia si Tunisia. Toate cele trei rute evita zonele largi ale Mediteranei.
In legatura cu originea istorica a migratiilor se presupune ca au aparut ca urmare a schimbarii regimului de hranire (cum este cel insectivor); se admite de asemenea, ca migratiile au fost determinate de modificarile climatice din trecut si odata cu deplasarea zonelor climatice s-au deplasat, de la nord spre sud si invers, atat zona de cuibarit, cat si cea de iernat si prin aceasta si caile de migratie (P. Banarescu, N. Boscaiu, 1973).
3. Modalitati de raspandire a vietuitoarelor
Plantele si animalele se raspandesc in mediul in care traiesc, la distante mai mici sau mai mari, prin mijloace active (diseminarea activa) sau pasiv, prin intermediul factorilor naturali extemi (vant, apa, animale) sau prin interventia omului (diseminarea pasiva).
Dupa modul in care isi raspandesc fructele si semintele, plantele se c1asifica in doua mari categorii: autochore, care isi raspandesc semintele prin mijloace proprii, si allochore care isi raspandesc fructele si semintele cu ajutorul unor agenti straini, functie de care plantele se numesc anemochore, zoochore si hidrochore.
Daca aceste posibilitati de raspandire sunt reduse, perpetuarea organismelor este asigurata printr-o capacitate de inmultire foarte mare, sau prin rezistenta sporita la conditii foarte severe de viata, sau longevitate. Aceste capacitati sau adaptari sunt specifice fiecarei specii in parte si difera foarte mult, uneori la grupe inrudite.
Diseminarea pasiva este caracteristica plantelor si mai putin animalelor. Sub actiunea vantului, apei, a altor animale sau omului, sporii, semintele sau fructele si, in unele cazuri plantele intregi sunt transportate la distante foarte mari, unde germineaza si dau nastere altor plante asemanatoare celor din care au provenit. Aceasta forma de raspandire poarta denumirea de anemocorie, iar speciile anemocore prezinta numeroase adaptari in acest sens:
- greutatea semintelor este foarte redusa: 0,006 mg la poroinic (Orchis fusca) si Parnassia palustris, 0,025 mg semintele de smirdar (Rhododendron), la Ericaceae, etc.;
- semintele sau fructele sunt sferice la unele specii, fiind usor rostogolite de vant pe solul neted al stepelor (Fabaceae, Brassicaceae, Caryophyllaceae) sau prezinta o morfologie aparte, fiind prevazute cu prelungiri sub forma de puf sau peri dispusi ca o umbreluta, sau pe toata suprafata; exemple:
- seminte cu o coroana de peri la un capat: la Epilobium (pufulita), salcie (Salix), plop (Populus), sau pe toata suprafata tegumentului la bumbac (Gossypium);
- seminte aripate la Catalpa si Bignonia;
- fructe aripate la Acer, Ulmus, Fraxinus, Betula;
- fructe cu o umbrela de peri numit papus, la papadie (Taraxacum), palamida (Cirsium), bumbacarita (Eriophorum), sau cu o arista penata la colilie (Stipa);
-intreaga inflorescenta cu fructe este purtata de vant datorita prezentei unei bractei ca la tei (Tilia);
- in regiunile deschise de stepa sau semidesertice, plante uscate pline de seminte (roza Ierichonului: Anastatica hierochuntica din zonele semiaride ale Egiptului si Israelului, chiurlanul: Salsola kali din stepa rusa, intalnit si la noi in Baragan sub denumirea de ciulini), purtate de vant pe distante foarte mari, imprastiind in trecerea lor semintele pe care le poarta.
Zoochoria este raspandirea pasiva sau activa de catre animale a plantelor. In mod activ: animalele frugivore consuma fructele carnoase de la unele specii de plante, semintele nefiind distruse in tubul digestiv pot fi transportate la mari distante (in special de catre pasari); plantele cu acest mod de raspandire se mai numesc endozoochore. In mod pasiv: animalele transporta pe lana, parul, penele si picioarele lor fructe, seminte si plante prevazute cu carlige sau tepi incovoiati la Galium (turita), Lappula (lipici), Xanthium, Arctium (brusture); unele plante de apa (Lemna) sau semintele unor plante de locuri mlastinoase se prind odata cu noroiul de picioarele animalelor si astfel sunt raspandite la distanta.
Hidrochorele sunt specii de plante ale caror fructe si seminte au adaptari speciale la transportul cu ajutorul apei: aerenchim bine dezvoltat (pentru a pluti la suprafata apei), pericarp si tegument impermeabil pentru apa, cel putin pentru o anumita perioada de timp (la Cucuta virosa cucuta de apa, specii de rogoz Carex, stuf sau trestie: Phragmites, papura: Typha, plop: Populus, salcie: Salix, Cocos nucifera - nuca de cocos poate pluti mii de km purtata de curentii marini).
Si animalele pot fi raspandite cu ajutorul vantului sau apei, uraganele tropicale purtand la distante mari animale mici (insecte, broaste, pasari), iar apele curgatoare, mai ales in timpul viiturilor pot transporta arbori desradacinati de care se prind moluste, viermi, reptile.
Diseminarea activa este intalnita mai mult la animale si mai putin la plante.
Plantele autochore isi raspandesc semintele prin mijloace proprii, avand unele adaptari in acest sens cum ar fi:
- prezenta in structura fructului a unor tesuturi mecanice care favorizeaza deschiderea brusca pe timp de uscaciune cu aruncarea semintelor la distante mai mari sau mai mici; astfel:
-la specia Impatiens noli-tangere (calatorule nu ma atinge!) capsula suculenta se desface brusc in cinci valve care se rasucesc in spirala, semintele fiind aruncate la 1-2 m;
- la specia Ecballium elaterium (plesnitoare), ajunsa la maturitate, presiunea lichidului din fruct determina in acelsi timp detasarea fructului de peduncul si tasnirea lichidului cu seminte la cativa metri;
-la specia Hura crepitans capsula se deschide brusc si cu zgomot, semintele fiind aruncate pana la 25 m de planta-mama.
Animalele avand posibilitati proprii de deplasare sunt capabile sa-si extinda arealele raspandindu-se activ pe mari distante; unele specii acvatice innotand cu usurinta, ocupa bazine marine imense; de asemenea, unele animale terestre (lupul, vulpea, iepurele, unele pasari) sunt raspandite pe intinse areale geografice.
Prin interventia antropica in raspandirea plantelor si animalelor care poate fi voluntara sau involuntara s-au diseminat numeroase specii de plante si animale, unele foarte importante din punct de vedere economic, altele dimpotriva, buruieni daunatoare sau specii de animale care s-au dezvoltat exploziv (in lipsa dusmanilor cu rol de control in inmuitirea acestora) provocand adevarate calamitati asupra ecosistemelor naturale din anumite zone geografice. Astfel, prin actiuni voluntare s-au adus numeroase specii de plante de cultura: in sec. XVI din America de Sud s-au adus cartoful (Solanum tuberosum), patlageaua rosie (Lycopersicum esculentum), cultivata in Franta, Italia, Anglia in sec. XVII, la noi fiind introdusa abia in sec. XIX, tutunul (Nicotiana tabacum) cultivat in Franta incepand cu anul 1560; in jurul Marii Mediterane s-au adus agavele si opuntia; in Europa s-au adus numeroase plante ornamentale (printre care freziile si muscatele). Alte specii au fost transportate intamplator: patlagina (Plantago major) si tortelul (Cuscuta epilinum) au fost duse din Europa in America de Nord, unde s-au raspandit pe tot continentul. Unele specii aduse pentru colectiile unor gradini botanice au scapat din cultura acestora raspandindu-se cu viteza pe continente intregi. Astfel Amaranthus retroflexus (stirul), cultivat de Linn la Upsala (Suedia) in 1750 a scapat din cultura, a trecut in restul tarilor din Europa, apoi in Asia Mica, Caucaz, Siberia, Africa de Nord. La noi a aparut in Transilvania in 1816; in doua secole aceasta planta s-a raspandit pe tot globul. O alta buruiana, Galinsoga parviflora (busuiocul dracului, raia pamantului) a scapat la 1800 din unele gradini botanice din Germania, in 1880 este semnalata in Rusia si Norvegia, in 1909 la noi in tara in judetul Prahova, astazi fiind raspandita in toate judetele.
Dintre speciile de animale, a caror raspandire a fost influentata de om, mentionam cateva exemple: aducerea in Europa si Siberia din America de Nord a bizamului (ondatra) (Ondatra sibethica); introducerea graurilor (Sturnus vulgaris) din Europa in America de Nord; introducerea iepurelui de casa in Australia (1840), care in lipsa camivorelor de prada s-a inmultit foarte mult producand o adevarata calamitate ecologica; inlocuirea sobolanilor sud-americani (Sigmadon) cu sobolanul negru (Rattus rattus) din Europa, dus cu corabiile pe la 1544; aclimatizarea unor specii de pesti originari din America in Europa si mai ales in raurile din tara noastra (pastravul american sau curcubeu: Salmo irideus, fantanelul, somnul american, bibanul soare)
4. Factorii paleogeografici ai raspandirii vietuitoarelor
Configuratia continentelor si oceanelor a suferit de-a lungul erelor geologice numeroase si ample modificari, asupra evolutiei acesteia fiind emise mai multe teorii, sustinute de argumente geologice, paleontologice si biogeografice. Desi, la momentul actual, teoria mobilitatii blocurilor continentale este aproape unanim acceptata, vom prezenta pe scurt trei dintre cele mai importante:
I. Teoria puntilor continentale. Aceasta admite existenta unor legaturi intre continente in anumite perioade din trecut. Se accepta cinci continente intermediare sau punti intercontinentale:
- puntea sau continentul Nord-Atlantic dintre America de Nord, Groenlanda, Islanda si Europa, care ar fi functionat in repetate randuri in Paleozoic, Mezozoic si la inceputul Tertiarului;
- Gondwana ar fi grupat din Carbonifer pana in Triasic sau chiar Jurasic, America de Sud, Africa, Madagascar, India peninsulara, Australia, Antarctida si eventual Noua Zeelanda; fragmentarea Gondwanei ar fi inceput in Triasic si ar fi durat pana in Jurasic;
- puntea africano-braziliana, rezultata din fragmentarea Gondwanei, reunea partile tropicale ale Americii de Sud si Africii si a functionat pana in Cretacic;
- Lemuria, fragment al Gondwanei, facea legatura intre Madagascar si India din triasic pana la sfarsitul Cretacicului; .
- puntea antarctica ar fi legat Patagonia si Antarctida cu Australia sau Noua Zeelanda (eventual cu amandoua) din Cretacic pana la inceputul Tertiarului;
- alte punti mai putin importante cum sunt: America Centrala, care a aparut in pliocen, legind cele doua Americi separate in Tertiar, lantul muntilor Urali care s-au ridicat 'in oligocen unind Europa si Asia, pana atunci separate de un brat marin, si, o punte continentala actuala, incompleta, constand dintr-un lant de insule, este formata din insulele Indoneziei, dintre care cele vestice (Sumatera, Djawa, Kalimantan) apartin Asiei, cele estice (Noua Guinee si cateva insule vecine) apartin lumii australiene, iar cele centrale (Sulawesi, Moluce, Timor etc.) formeaza adevarata punte de legatura, incompleta inca, intre doua continente care au evoluat independent unul fata de celalalt.
2. Teoria permanentei continentelor si oceanelor. Aceasta teorie se opune teoriei puntilor continentale, sustinand ideea pastrarii configuratiei continentelor si oceanelor din paleozoic pana in zilele noastre, singurele modificari pe care Ie recunosc fiind limita dintre uscat si mare si marile epicontinentale care inaintau sau se retrageau. Teoria a fost emisa mai ales pe baza unor argumente geofizice, dar a fost acceptata de numerosi biogeografi care, in imposibilitatea unei explicatii a raspandirii unor grupe au recurs la rolul vantului ori al curentilor marini. Adept al acestei teorii, zoogeograful Ph. Darlington (1965) prezinta astfel evolutia in timp si spatiu a vietuitoarelor:
formarea speciilor in zona tropicala a Lumii Vechi (in primul rand in sudul Asiei);
patrunderea in zona temperata nordica si cea tropicala: Asia si Europa, iar din Asia estica in America de Nord si de aici in cea de Sud;
patrunderea in zona sudica temperata: in Australia si Noua Zeelanda, dinspre Asia, iar in partile reci ale Americii de Sud din zona tropicala a aceluiasi continent;
disparitia treptata a multor specii in zona tropicala a Lumii Vechi, ulterior si in cea a Americii de Sud;
disparitia treptata din zona temperata nordica;
supravietuirea numai in zona temperata sudica.
3. Teoria translatiei continentelor, cunoscuta si sub numele de "teoria lui Wegener", dupa numele geofizicianului care a elaborat-o (si prezentat-o in 1928 in forma clasica) sau "a driftului continental", sustine existenta initiala a unui singur bloc continental "Pangea", care s-a fragmentat in timp (exista mai multe variante ipotetice ale fragmentarii), pastrandu-se legaturi intercontinentale periodice care au favorizat schimburi de flora si fauna; prin aceasta teorie, s-a constatat paralelismul tarmurilor (estic cu vestic al Atlanticului de sud, incadrarea Antarcticii intre sudul Americii de Sud, Africa, Madagascar, India, si Australia), iar biogeografii au explicat asemanarile floristice si faunistice intre tarmurile opuse, intre continentele legate prin punti; contestata si adusa iar in prim plan, de aceasta data sustinuta de cercetari paleomagnetice, aceasta teorie nu mai reprezinta o ipoteza, ci un fapt bine stabilit, verificat, reconstituit si completat cu ajutorul calculatoarelor moderne.
Consideratii biogeografice sintetice privind teoriile paleogeografice
Din cele trei teorii prezentate mai sus teoria puntilor continentale a fost inlocuita treptat de teoria translatiei continentelor, in schemele initiale propuse de Wagener, iar adeptii permanentei continentelor s-au redus pe masura ce s-au acumulat noi si incontestabile argumente in favoarea mobilitatii blocurilor continentale.
Daca numeroase fapte biogeografice pot fi explicate prin ambele teorii, unele se explica mai usor prin teoria translatiei: particularitatile florei si faunei australiene, prezenta unor grupe de organisme numai in continentele sudice sau asemanarile dintre ihtiofauna sud-americana si cea africana.
Ne vom referi pe scurt la primul exemplu:
Australia, desi este situata destul de aproape de sud-estul Asiei (considerat, pentru Tertiar, principalul centru de evolutie al grupelor modeme de animale), de care se leaga incomplet prin insulele Indoneziei, contine in flora si fauna sa numeroase grupe arhaice si doar cateva elemente asiatice moderne. Acceptandu-se teoria permanentei continentelor se pune intrebarea - de ce elementele asiatice nu au patruns de timpuriu si au patruns numai cele moderne, tarzii? Acceptandu-se teoria lui Wagener, Australia (initial vecina cu Africa, India, Madagascar, Antarctida si America de Sud) a ajuns in vecinatatea Asiei abia in Miocen; se pot explica astfel afinitatile cu teritoriile foste vecine si particularitatile capatate in timpul indelungii izolari care avea sa urmeze.
Pe baza argumentelor biogeografice, dar mai ales geofizice, translatia continentelor este considerata nu o ipoteza, ci un fapt "cvasicert", ce a avut urmatoarea evolutie: in Tertiar marile blocuri continentale actuale, deja separate intre ele, au continuat sa se indeparteze unele de altele, astfel incat Australia ajunge sa se apropie mai mult de Asia; in Cretacic orice contact direct intre continentele sudice era intrerupt. Distributia geografiea a grupelor de vietuitoare care au evoluat si s-au raspandit ulterior (in era tertiara) nu mai reflecta translatia continentelor.
Raspandirea actuala a plantelor si animalelor este rezultatul evolutiei paleogeografice a globului din Paleozoic si mai ales din Mezozoic pana in zilele noastre si ale carei etape principale (dupa P.Banarescu si N.Boscaiu, 1973) sunt urmatoarele:
I. existenta, in paleozoic pana in Triasic sau Jurasic a unui singur bloc continental (Pangea) sau a doua blocuri, unul nordic (Laurasia) si altul sudic (Gondwana); grupele de plante si animale se puteau raspandi liber intre Europa si America de Nord si respectiv, intre toate continentele si regiunile sudice (inclusiv Madagascarul, India, Noua Zeelanda);
2.fragmentarea continentului sudic (Gondwana) si indepartarea diverse lor fragmente a caror flora si fauna au evoluat ulterior independent; intre unele dintre fragmentele Gondwanei au persistat legaturi pana la sfarsitul erei secundare: intre America de Sud tropicala si Africa si intre sudul Americii de Sud, Australia si Noua Zeelanda, prin intermediul Antarctidei;
3. existenta, in cea mai mare parte a Tertiarului, a unui imens continent asiatic caruia i s-a atasat India (fragment al Gondwanei) si Europa si care a fost aproape permanent legat de America de Nord, prin zona actualei stramtori Bering; sudul tropical al Asiei a fost, in cea mai mare parte a Tertiarului, principalul centru de evolutie al grupelor competitive de animale si de plante, care tindeau sa se raspandeasca pe intreg globul; au fost astfel inlocuite flora si fauna veche din India, flora si fauna de clima temperata din nordul Asiei au populat Europa si America de Nord; se considera ca genurile florei actuale au aparut aproape in totalitate la sfarsitul Cretacicului (in urma cu peste 70 milioane ani);
4. in miocen Africa s-a atasat de Asia numeroase grupe de plante si animale tropicale atat de savana, cat si de padure, originare din sudul Asiei, au inceput sa populeze Africa, inlocuind aproape complet vechea flora si fauna de origine gondwaniana; incepand cu sfarsitul Tertiarului, principalul centru de evolutie si raspandire a grupelor competitive de animale pare sa se fi deplasat din sudul Asiei in Africa; odata cu formarea, foarte recenta, a Marii Rosii, Africa s-a izolat din nou de Asia;
5. America de Sud, fragment al Gondwanei, a stabilit la inceputul Tertiarului, o legatura temporara cu America de Nord de unde a patruns o serie de grupuri de plante si animale (mai ales mamifere) caracteristice teritoriului numit Megagea (Eurasia, Africa si America de Nord); aceasta legatura a durat putin timp, cea mai mare parte a Tertiarului, America de Sud fiind izolata, timp in care grupele care patrunsesera din alte continente au evoluat intr-un mod particular; legatura care se restabileste cu America de Nord in Pliocen duce la inlocuirea partiala a florei si faunei cu grupe venite din America de Nord;
6. Australia provenind tot din Godwana, s-a deplasat spre nord-est, ajungand prin Miocen, in apropierea Asiei de sud totodata s-a stabilit o legatura incompleta intre aceste doua mase continentale, prin care au patruns in nordul tropical al Australiei vietuitoare originare din sud-estul Asiei, dar numai dintre cele capabile sa se raspandeasca peste brate marine si anume: pasari si alte animale zburatoare, anumite genuri de plante;
7. in perioadele lungi in care blocurile continentale erau separate prin mari, izolarea lor faunistica si floristica nu a fost completa, caci o serie de plante si animale au putut depasi bratele marine, gratie posibilitatilor lor de zbor sau de raspandire pasiva; aceste schimburi floristice sau faunistice au fost posibile mai ales intre blocurile continentale separate, dar situate la distante mici - asa a fost situatia celor doua Americi din eocen pana in pliocen; izolarea a fost deplina, faunele de mamifere, pesti primar-dulcicoli si amfibieni au evoluat independent, in schimb intre faunele de pasari schimbul a fost permanent;
8. puternicele fluctualii climatice din cuatemar (care s-au manifestat sub forma de perioade glaciare si interglaciare in zonele temperate, de perioade pluviale si interpluviale in cele tropicale) au intluentat foarte mult raspandirea plantelor si animalelor, au intrerupt in mod radical o evolutie care a durat lent si continuu timp de milioane de ani.
"Se poate conchide ca pentru unele grupe raspandirea actuala se explica prin translatie continentala, iar pentru altele prin raspandirea pe calea ocolita nordica", concluzioneaza autorii mai sus citati.
5. Glaciatiunile cuaternare si consecintele lor biogeografice
Spre deosebire de glaciatiunile precuatemare, puse in evidenta numai in emisfera sudica, glaciatiunile pleistocene foarte severe au acoperit in mai multe faze teritorii foarte intinse pe ambele emisfere (cea mai mare parte a Americii de Nord, vestul si centrul Europei, sudul Americii de Sud si al Australiei, Noua Zeelanda), afectand covarsitor compozitia florelor si faunelor. Aceste perioade glaciare cu durata de 10.000-50.000 ani sunt o succesiune de mai multe oscilatii climatice foarte reci, numite stadii, si temperate (de incalzire a climatului), numite interstadii, ce au influentat radical majoritatea grupelor de vietuitoare.
Pentru Europa Centrala cele patru faze de expansiune a ghetarilor s-au denumit: Gnz, Mindel, Riss si Wrm, separate de trei perioade calde denumite si interglaciare: Gnz-Mindel, Mindel-Riss, Riss-Wrm.
Pentru nordul Europei s-au separat trei glaciatiuni: Elster, Saale, Vistula cu perioadele interglaciare Holstein si Eem. in partea europeana a Fed. Ruse au fost identificate trei glaciatiuni: Oka, Nipru, Valdai, separate de interglaciarele Lichvin si Mikulino (Minsk).
Pentru America de Nord s-au identificat patru faze de extindere a ghetarilor: Nebraska, Kansas, Ilinois si Wisconsin, separate de perioadele interglaciare Aftonian, Yarmouth si Sangamon.
Sincronizarea si echivalarea denumirilor reflecta faptul ca fazele glaciare au avut caracter global, iar manifestarea lor a avut aspect regional.
Cu ajutorul analizei sporo-polinice, palinologia a descifrat evolutia climatului cuatemar, reconstituind istoricul vegetatiei si modificarile suferite de covorul vegetal ca urmare a instabilitatii climatice. S-au separat astfel diteva tipuri de perioade fitoclimatice:
perioade anaterme (de incalzire a climatului), relevate de scaderea frecventei de polen de pin si de mesteacan si de cresterea frecventei polenului unor esente cu cerinte mai termofile, in special extinderea alunului;
perioade hipsoterme (de culminare a climatului calduros), relevate de frecventa mare a esentelor temofile (stejar, tei, ulm, carpen);
perioade cataterme (de racire a climatului), cu regresul esentelor termofile si revenirea celor microterme.
S-au separat de asemenea, in cadrul perioadelor reci epoci arctice si subarctice, iar in cadrul perioadelor calde, epoci cu climat umed numite atlantice si epoci cu climat mai rece si uscat numite boreale.
6. Evolutia climatului si a vegetatiei cuatemare
Scaderea temperaturii de la inceputul Cuatemarului a determinat inlocuirea florei termofile ramasa din Pliocen cu una caracterizata de cerinte mai microterme.
Primele glaciatiuni - Gnz si Mindel - au fost mai putin riguroase, iar interglaciarul care le-a separat a fost scurt si nu a permis reinstalarea esentelor termofile. Interglaciarul Mindel-Riss (Holstein) a fost mult mai lung decat primul si cu un climat mult mai cald decat cel actual.
Glaciatiunea Riss a fost cea mai extinsa, inregistrand doua stadii cataterme separate de o oscilatie anaterma. Europa centrala era acoperita de tundre cu salcii pitice (Salix polaris, s. retusa), mesteacan pitic (Betula humilis, B. nana), argintica (Dryas octopetala). Interglaciarul care a urmat a fost mai bland decat cel actual, favorizand dezvoltarea asociatiilor termofile.
Glaciatiunea Wrm ar fi durat aprox. 65.000 ani si a constat intr-o succesiune de stadii (reci) si interstadii (temperate). In decursul stadiilor se dezvolta o flora arctica-alpina si subarctica ierboasa si arbustiva, iar in timpul intrestadiilor vegetatia era alcatuita din paduri de molid, stejar, ulm, frasin, carpen si alun.
Ultima perioada a complexului wiirmian (Tardiglaciar) a avut temperatura medie mai coborata cu 8-12C decat cea actuala si a fost denumita Dryas dupa denumirea uneia dintre plantele caracteristice. Se considera ca tardiglaciarul incepe acum 20 000 ani, odata cu retragerea ghetafilor si dureaza pana acum 8 800 ani cand Finlanda a fost eliberata de ghetari. Intervalul cuprins intre ultima glaciatiune si perioada actuala este foarte bine cunoscut sub aspectul evolutiei c1imatice din interpretarea si omologarea datelor sporo-poliniee obtinute pentru diverse zone din Europa.
Schema de referinta pentru periodizarea Postglaciarului, elaborata de Blytt si Semander, sincronizata cu zonele poliniee stabilite de Firbas, a diferentiat zece zone polinice bine individualizate, valabile si pentru evolutia vegetatiei postglaciare de la noi, reconstituita de Emil Pop si colaboratorii (1970) pe baza unei diagrame sporo-polinice de la Taul Zanogutii (Retezat):
Zona I sfarsitul perioadei tundrelor arctice (Dryas-ul vechi) climatul arctic continental era propice dezvoltiirii tundrelor cu ierburi si arbusti in Europa centrala; in Carpati se succedau episodul pinetelor aride vechi cu jneapan (Pinus mugo), un episod Pinus-Picea intr-o perioada de incalzire (interstadiul Bolling) si un episod al pinetelor aride noi in climatul cataterm ce a urmat;
Zona II interstadiul Allerod sau perioada subarcticii mijlocie oscilatie anaterma careia ii corespunde un episod de mesteacan-pin in Europa centrala si, un episod al pinetelor cu molidis bogat in Carpatii romanesti; dureaza intre 12.500 si 10.700 i.C.;
Zona III perioada subarctica recenta sau a tundrelor parcuri noi (Dryas-ul recent) - sfirsitul tardiglaciarului cu o noua expansiune a tundrelor in care vegetau si raristi de pin;
Zona IV preborelul sau faza de tranzitie pin-molid - incalzirea climatului a favorizat extinderea padurilor de pin si molid in Europa centrala; in Carpati s-au extins masiv molidisurile;
Zona V borealul sau faza extinderii molidisurilor cu stejaris amestecat si alun - climatul calduros si uscat ajunge la apogeu, molidisurile se extind spre altitudine, foioasele termofile (stejar, ulm, lei) si alunul ocupau locurile mai joase;
Zona VI si VII atlanticul sau apogeul molidisurilor cu stejaris amestecat - domina climatul hipsoterm postglaciar cald si uscat la inceput, apoi treptat umed, foarte favorabil vegetatiei forestiere care se dezvolta exploziv; spre sfarsitul fazei in Carpati se raspandesc carpenul si fagul;
Zona VIII subborealul sau faza molidisurilor cu carpen - climatul devine mai racoros si mai uscat - in competitia biocenotica speciile mai putin adaptate sunt eliminate; in Carpati, intre stejarete si molidisuri se interpune o zona de carpinise;
Zona IX si X subatlanticul sau faza fagetelor - climatul racoros si umed a favorizat extinderea masiva a fagului pe seama carpenului a carui zona o ocupa, si a celorlalte esente care sufera o reducere a frecventei lor; in Europa centrala ca si in tara noastrii devine principalul edificator climatogen al vegetatiei montane;
Zona X, pe masura ce ne apropiem de prezent, se reflecta influenta antropica din ce in ce mai mare in dinamica vegetatiei.
Consecintele biogeografice ale glaciatiunilor
1. Oscilatiile climatice pleistocene si altemanta dintre perioadele glaciare si interglaciare au determinat migratii, deplasari alternative de la nord spre sud si invers (pentru emisfera boreala - Fig. 19, si invers pentru cea australa) ale florei si faunei mai adaptabile (deoarece contactul cu factori de mediu schimbati - substrat geologic, sol, altitudine, factori biotici - a dus la eliminarea speciilor cu cerinte stricte si supravietuirea celor cu valenta ecologica mai larga, mai usor adaptabile). Prin urmare, s-a inregistrat o saracire generala si treptata a fiorei si faunei preglaciare (P.Banarescu, N.Boscaiu, 1973). Au disparut astfel din Europa genuri ea Liriodendron, Catalpa, Magnolia si Pterocarya, s-au conservat pe arcale restranse genuri ca Sequoia si Taxodium in America de Nord, Metasequoia si Glyptostrobus in Asia, sau au supravietuit ca relicte litogeografice in statiuni adapostite (refugii) in care conditiile nu s-au modificat foarte mult (lotusul tropical Nymphaea lotus var. termalis si gasteropodul de apa dulce Melanopsis pareyssi rezista din Tertiar in apele termale ale paraului Petea de langai Oradea).
2. Elementele faunistice si floristice "arcto-tertiare", cu raspandire circumpolara aproape uniforma in cele trei continente la sfarsitul Tertiarului, datorita conditiilor climatice asemanatoare, au fost cel mai afectate de sehimbarile climatice deoarece deplasarile spre sud in timpul recrudescentei frigului si revenirile spre nord in interglaciare nu s-au petrecut uniform (elementele arcto-tertiare ramase pe varful muntilor la retragerea ghetarilor in interglaciare se numesc elemente arcto-alpine).
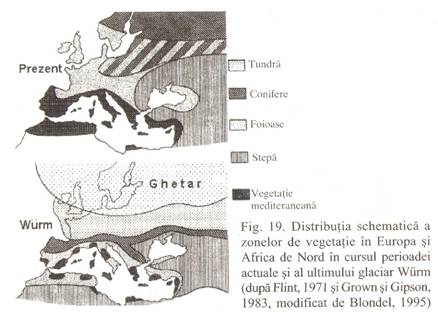
Diferentele mari s-au datorat in primul rand dispunerii diferite a lanturilor muntoase in cele trei continente. Astfel in America de Nord lanturile muntoase orientate de la nord la sud, si mai ales in Asia estica unde nu sunt munti inalti, nu au existat piedici mari in calea migratiilor. In Europa, muntii avand orientarea vest-est, au constituit adevarate bariere in calea migratiilor, ei insisi fiind acoperiti de ghetari - in acest caz au disparut multe elemente arcto-tertiare. In Siberia, desi se pare ca a lipsit calota glaciara (precipitatiile fiind foarte reduse cantitativ), excesivitatea climatului, pe de o parte, si lanturile muntoase foarte inalte din sud (Tian San, Altai), au constituit o bariera de netrecut in calea florei si faunei.
Ca urmare a deosebirilor mari de migrare nord-sud si invers, au supravietuit un numar mare de elemente arcto-tertiare in Asia estica si sud-estul Americii de Nord, un numar mic in sudul Europei (si de asemenea in Asia vestica si nordul Africii) si aproape deloc in Siberia. Arealul continuu al multor genuri a devenit discontinuu sau disjunct. Se cunosc mai multe tipuri de disjunctii:
disjunctii est asiatice-nord americane - unele elemente termofile (genuri de pesti, amphibieni, reptile si de plante), in special arhaice, supravietuiesc numai in Asia estica si sudul Americii de Nord; glaciatiunile le-au restrans arealele si le-au suprimat in Europa;
disjunctii eurasiatice-nord americane - genuri de stepa si semidesert din podisurile si stepele central-asiatice, pe de o parte, si sud-vestul Americii de Nord, pe de alta parte, care si-au restrans si mai mult arealele; si aceste disjunctii sunt mai vechi;
disjunctii europeo-nord americane - se refera la unele genuri de animale de apa dulce, elemente arcto-tertiare ramase numai in Europa si America de Nord;
disjunctii est-vest palearctice - genuri de pesti de apa dulce din Europa si Asia cstica, lipsind din Siberia datori!a glaciatiunilor si din America de Nord datorita raspandirii eurasiatice (palearctice).
unele specii si genuri cu arealul fragmentat in trei (Europa, Asia estica, America de Nord) prezinta in acelasi timp o disjunctie europeo-nord-americane si una est-vest- palearctica.
3. Tot o consecinta a schimbarilor climatice majore sunt relictele tertiare care au supravietuit in refugiile glaciare, extinzandu-si apoi arealele; aceste refugii au devenit astfel centre de raspandire postglaciara. Se considera ca foarte importante, pentru elementele de padure si campii umede, urmatoarele refugii: mediteranean, armeanopersic, est-asiatic, afganistan, Tian San si Mongolia, pentru Eurasia si cateva pentru America de Nord. Fauna dulcicola a avut mai multe refugii in spatiul mediteraneean: iberic, italian, vest-balcanic, ponto-caspic, acestea diferentiindu-se prin specii diferite.
4. Zonele tropicale au fost, de asemenea, afectate de oscilatiile climatice cuaternare, alternanta de perioade pluviale si interpluviale favorizand pe rand expansiunea florei si faunei de padure ecuatoriala, rcspectiv a savanei, in perioadele interpluviale secetoase.
5. Numarul de taxoni din flora si fauna a suferit numeroase modificari, nu toate in sensul reducerii acestuia, ci si al aparitiei de noi specii in urma proceselor de subspeciatie si speciatie favorizate de izolarea geografica. Asa se explica numarul foarte mare de specii inrudite care caracterizeaza fiecare alt refugiu din Asia Centrala sau America de Nord. Se afirma ca glaciatiunea a dus la reducerea numarului total de genuri preglaciare si, totodata la cresterea numarului total de spec
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 9210
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved