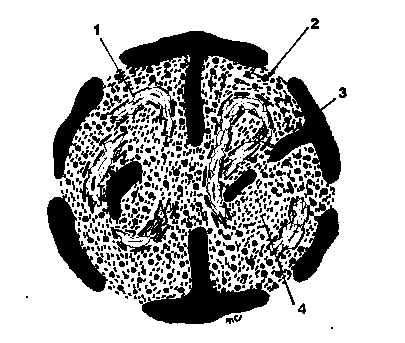| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
NUCLEUL
Nucleul (lat. nucleus si gr. karion = sambure) (nucleus) este o componenta esentiala caracteristica celulelor eucariote. Indeplineste ca functii principale: -stocarea informatiei genetice in ADN-ul nuclear , reglarea si controlul tuturor activitatilor celulare. In organismele animale, exista si celule anucleate (hematiile, trombocitele), care sunt incapabile sa sintetizeze proteine, activitatiile lor metabolice fiind foarte restranse.
Componentele nucleului
|
Denumire |
Componente |
Formatiuni ultrastructurale |
|
Membrana nucleara externa | ||
( invelisul nuclear) |
Cisterna perinucleara |
Complexul por: Anulus extern Anulus intern Granula centrala 8 granule proteice periferice 8 conuri de proteine fibrilare |
|
Membrana nucleara interna | ||
|
B. Nucleoplasma |
Scheletul nuclear |
Matricea fibrilara (granule si matrixin ) Lamina densa interna Complexul lamina por Componenta fibrilara a nucleolului |
|
Fractiune labila | ||
|
Cromatina |
Eucromatina Heterocromatina |
|
|
C. Nucleolul |
Componenta filamentara |
Filamente de 5 nm grosime/30-40 nm lungime |
|
Componenta granulara |
Precursori ribozomiali |
|
|
Componenta amorfa | ||
|
Cromozomiala |
Cromatina perinucleolara Cromatina intranucleolara |
Nucleul a fost
descoperit de
In alcatuirea nucleului intra: nucleolema sau membrana nucleara, nucleoplasma (matricea sau sucul nuclear), nucleolii si cromatina sau cromozomii.
Nucleul reprezinta centrul de comanda, control si de coordonare a celulei eucariote, functionand ca un computer chimic prevazut cu un program (ADN-ul) si o memorie (ARN-ul). In calitate de centru informational al celulei, nucleul coordoneaza toate reactiile chimice legate de desfasurarea proceselor vitale, contrland atat sinteza proteinelor enzime din celula, cat si sinteza proteinelor de structura. (Fig.1.)
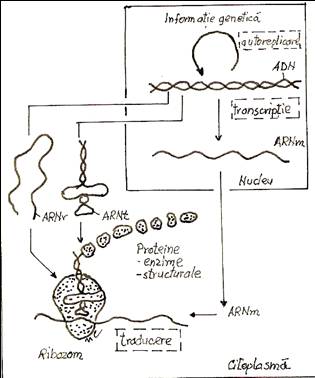
Fig.1. Rolul nucleului in circulatia informatiei genetice in celula eucariota.
Nucleul contine genomul (totalitatea genelor) celulei, format dintr-un numar precis de gene. O gena este o secventa de nucleotide (500-1500), care determina o secventa de aminoacizi, impreuna cu elementele de control (un promotor, o secventa lider si un terminator. La animalele domestice, gena ocupa un loc specific in molecula de ADN care intra in compozitia cromozomului.
Intr-o celula umana, lungimea totala a moleculelor de ADN este de cca. 1,6 m continand un potential informational imens, estimat la aproximativ 9 X 1011 biti, ce corespunde informatiilor cuprinse intr-o biblioteca cu 180.000 volume (considerandu-se ca un cuvant este evaluat in medie la 36 biti, iar in fiecare volum se gasesc 150.000 cuvinte).
|
|
Totodata nucleul contine echipamentul enzimatic necesar pentru: repararea genomului, replicarea sa, transcrierea mesajului in ARN si prelucrarea ARN-ului ( in ARN mesager, ARN de transfer si ARN ribozomal.
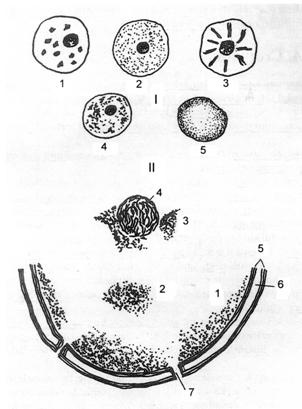
1-Veziculos (in neuron); 2-Multilobat (neutrofil); 3,11-Bilobat (eozinofil, pseudoeozinofil de pasare); 4-Aplatizat (adipocit); 5-Reniform (monocit); 6-Ovoidal (eritrocit); 7-Discoidal (celula caliciforma); 8-Alungit (leiocit); 9,10-Sferoidal sau ovoidal (limfocit)
1. Caracterele morfologice ale nucleului
Raspandire. Majorittea celulelor organismului prezinta nucleu. Exista si exceptii (hematiile,celulele cristaliniene, trombocitele), care reprezinta stadii finale, in evolutia unui tip celular, cu existenta limitata in timp, stadii specializate la functii pasive. Celulele fara nucleu nu pot creste si nu se mai divid.
Forma nucleilor este foarte variata,indicand, de regula, pe cea a celulei. Nucleul apare: - sferic-ovoidal (nucleus sphericus- ovoideus), in celulele izodiametrice (sferice,cubice,poliedrice);- alungit (bacilliformis,fusiformis), in celulele fusiforme (musculare) sau columnare (prismatice); - turtit,aplatizat (planus) in celulele pavimentoase (endoteliale),in adipocite, in celulele caliciforme; lobat (segmentalis s. moniliformis)), in celulele care trebuie sa se modeleze rapid.(Fig.2.)
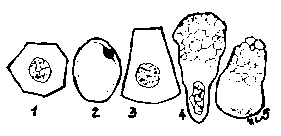
Forma nucleului este
influentata de activitatea celulei. Devine neregulata in
celulele foarte active. In acest mod se produce o crestere a
suprafetei prin care se realizeaza schimburile dintre nucleu si
citoplasma. Totodata nucleul prezinta o oarecare plasticitate
(deformabilitate) care ii permite ca, in anumite conditii spatiale intracelulare, sa isi
modifice forma. Asfel, in monocite nucleul apare reniform (nucleus reniformis), desi celula este sferica,
deformarea datorandu-se prezentei centriolului.
Fig.3. Pozitia nucleului in diferite celule.
1-Centrala; 2-Periferica; 3-Mediobazala; 4,5-Bazala.
Pozitia. In cele mai multe cazuri, nucleul ocupa o pozitie centrala, strategica pentru rolul de coordonator al activitatii celulare. Totodata exista numeroase exceptii, in care nucleul ocupa o pozitie: excentrica, in adipocite (celule care acumuleaza grasimi in citoplasma); medio-bazala, in celulele secretoare din pancreas sau parotida; bazala, in secretoare de mucus, in celulele caliciforme. In general in celulele secretoare.(Fig.3.)
Numar. Ca regula generala, o celula prezinta un nucleu. Exceptiile sunt insa numeroase. Astfel, hepatocitele sunt binucleate in proportie de 7-8%, osteoclastele au 30-40 nuclei, iar fibra musculara striata (rabdocitul) prezinta intre 20-40 nuclei pentru fiecare centimetru.
In conditii patologice, mai multi nuclei apar in:- celulele gigante Langhans (prezente in tuberculoza, au catva zeci de nuclei, dispusi la periferia celulei, in coroana sau potcoava); -celulele gigante de corp strain, care apar in cazul patrunderii in organism a unor particule straine, nedigerabile de catre celule; - celulele tumorale.
Multinuclearitatea se produce in doua moduri: 1- prin multiplicare nucleara repetata (cariokineza), fara diviziunea citoplasmei (citodiereza),rezultand un plasmodiu; 2-prin fuzionarea mai multor celule mononucleate, rezultand un sincitiu (cum este cazul osteoclastelor, rabdocitelor,celulelor Langhans,etc.)
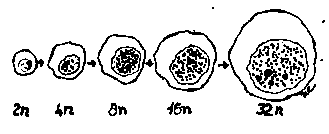
Fig. Variatia dimensiunilor nucleului in functie de gradul de poliploidie
Dimensiunile nucleului. Variaza in limite mai stranse (intre 5-15 ) decat dimensiunile celulelor, diversitatea dimensiunilor celulelor din organismul animal realizandu-se pe baza variatiei de volum a citoplasmelor.
Masurarea dimensiunilor nucleului se numeste cariometrie si reprezinta un domeniu de mare interes pentru biologia si patologia celulara. Nucleii prezinta variatii dimensionale functionale si patologice.
Variatiile functionale ale dimensiunilor nucleului sunt in legatura cu: ►Varsta, nucleul fiind mai mare in celula tanara, decat in celula imbatranita, in cazul aceluiasi tip celular;
► Bioritmul, existand diferente de volum de pana la 20%, intre zi si noapte, la hepatocite,neuroni, nefrocite,etc.;
► Gradul de poliploidie (numarul de seturi cromozomale prezente in nucleu). Cu cat gradul de poliploidie este mai mare cu atat nucleul este mai mare. Astfel, nucleii megacariocitelor sunt mult mai voluminosi decat cei a hepatocitelor. Din acest punct de vedere in hepatocite se pot intalni : 1 - nuclei mici (cu diametru de 10 m), diploizi (2n), intalniti la cca.80% din hepatocite; 2 - nuclei medii (cu diametru de 15 m); - tetraploizi (4 n), intalniti la 16% din hepatocite si uneori octaploizi. S-a constatat ca suprafata nucleara creste proportional cu gradul de poliploidie, fapt ce explica inmugurirea nucleilor poliploizi.
In patologie, variatii ale dimensiunilor nucleului se intalnesc in toxicoze, in boala de iradiere si in cancer. In celulele neoplazice, nucleii apar de talie mare, cu morfologie extrem de variata in acelasi camp microscopic, putand prezenta uneori aspect monstruos.+
Intre volumul nucleului si volumul citoplasmei se stabileste un raport nucleo-citoplasmatic (N/C), a carei valoare variaza in limite foarte largi (1/3 - 1/20), de obicei situandu-se intre 1/7 - 1/10.
Atunci cand volumul citoplasmei creste mai mult decat volumul nucleului, raportul N/C va fi refacut fie prin diviziune celulara, fie printr-o crestere a volumului nuclear. In cazul in care volumul nu poate fi refacut, functionarea normala a celulei este perturbata. In conditii patologice, raportul N/C poate fi complet modificat, ajungandu-se la inversarea raportului, in celulele tumorale.
Vascozitatea nucleoplasmei este mai mica decat a citoplasmei, exceptie facand ovocitul. Greutatea specifica a nucleului este influentata de continutul in apa si de starea fiziologica a celulei, descrescand in urmatoarea ordine: nucleol, cromatina, nucleoplasma.
Ph-ul nuclear este usor alcalin (7,4-7,5). Totodata, in nucleu se realizeaza diferente de potential intre membrana nucleara (cu sarcini electrice negative) si nucleoli sau cromozomi, ce au sarcini electrice pozitive.
Componentele nucleului sunt: membrana nucleara, nucleoplasma,cromatina si nucleolul.
2. Membrana nucleara
Membrana nucleara (nucleolema sau carioteca) (nucleolemma s. karyoteca) este o structura lipoproteica, cu o grosime de cativa zeci de nanometri, caracteristica eucariotelor care imparte celula in doua compartimente: - nucleul ce contine genomul, structurile implicate in transcriptie si in prelucrarea produsului transcriptic; - citoplasma ce cuprinde organitele celulare. In celulele vii, apare ca o pelicula fina, cu indice de refractie si densitate mai mare decat restul nucleului. In celulele fixate, se coloreaza bazofil, iar vizibilitatea cromatica se datoreste condensarii si coagularii unor componente ale carioplasmei care se depoziteaza pe fata interna a membranei.
La microscopul electronic, nucleolema apare formata din doua foite (una interna si alta externa), fiecare trilaminata, lipoproteica, groase de 60-100 , separate printr-un spatiu perinuclear sau cisterna perinucleara, de 150 - 300 , plin cu o substanta amorfa.
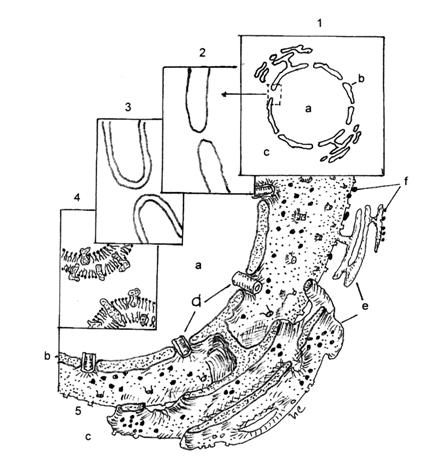
1-MN si relatiile sale cu reticulul endoplasmic (RE); 2-MN, la microscopul electronic,
cu grosisment redus; 3-Trilaminaritatea celor doua MN; 4-Organizarea moleculara a MN;
5-Ultrastructura MN .
a-Nucleul; b-Membrana nucleara; c-Citoplasma; d-Por; e-Reticul endoplasmic; f-ribozomi.
Membrana nucleara externa (membrana nuclearis externa) prezinta o fata citoplasmatica, ornata cu ribozomi, are un contur mai flexibil si zone active de formare a veziculelor.
Membrana nucleara interna (mebrana nuclearis interna) este lipsita de ribozomi si adera fie la nucleoplasma, care poate diferentia o lamina densa, fie la cromatina.
Cisterna perinucleara (cisterna nucleolemmae) apare ca un spatiu tridimensional, aflat in continuitate cu spatiul reticulului endoplasmatic. In acest spatiu s-au pus in evidenta proteine, enzime.Ca2+.(Fig.5)
Nucleolema sau invelisul nuclear este intrerupta (fenestrata) de orificii circulare, denumite pori circulari, la nivelul carora cele doua membrane se afla in raporturi de continuitate. Numarul de pori de pe o unitate de suprafata este legat de intensitatea schimburilor nucleo-citoplasmatice. Asfel, la celulele in crestere pot exista 10 pori/ m2. Se intalnesc variatii in numarul, frecventa si distributia porilor de la un tip de nucleu la altul. Porii sunt in fapt 'portile' folosite de macromolecule si de ansamblurile macromoleculare (proteine, ARN m, subunitati ribozomale,etc.) pentru a intra sau iesi din nucleu. (Fig.6)
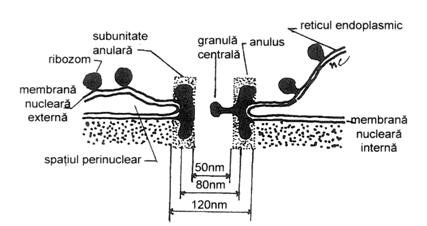
Fig.6. Structura porului nuclear.
Porul (porus nuclearis) impreuna cu structurile adiacente formeaza complexul porului.
Complexul por (complexus pori) cuprinde: a - doua inele sau anuli , asezate pe ambele fete ale porului (nucleara si citoplasmatica), fiecare inel (anulus pori) fiind compus din granule proteice sferoide cu diametrul de 10-25 nm, care sunt dispuse in forma de octagon; b - opt conuri, orientate dinspre peretele porului catre lumenul sau, considerate a fi agregate de fibrile; c - o particula centrala, in forma de granula sau bastonas, inconstant prezenta; d - pachete de filamente nucleoplasmatice de 4-8 nm, care se insera cu oextremitate de inelul intern, iar cu cealalta extremitate se prinde pe formatiuni intra nucleare (eventual pe nucleol).
Dupa unii autori, diferitele structuri care apar in centrul porului (diafragma porului, granula centrala, filamentele) par a fi mai curand materiale surprinse in timpul trecerii prin por decat structuri pemanente.
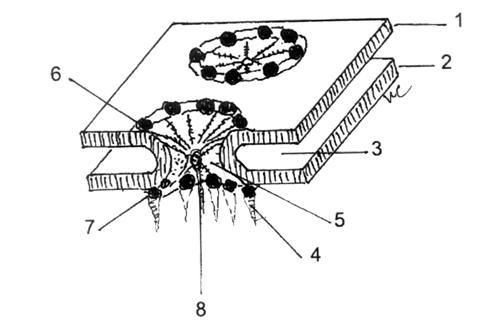
Fig.7.Schema complexului por.
1-MN externa; 2-MN-interna;3-Spatiu cisternal; 4-Filament intern; 5-Formatiune conica; 6-Granula-anulus extern; 7-Granula-anulus intern; 8-Granula centrala.
Functiile membranei
nucleare sunt: 1 -
delimiteaza continutul nucleului de cel al citoplasmei;2 -
regleaza schimburile dintre nucleu si citoplasma; 3 -
formeaza membranele reticulului endoplasmic, reprezentand rezerva de
citomembrane in celule care se divid rapid; 4 - mentine si
stabilizeaza cromatina, care se ataseaza de fata sa interna;
5 - rol mecanic in sustinerea organitelor, acestea fiind legate de
fata sa externa prin fibrile de 18 nm.(Fig.8.)
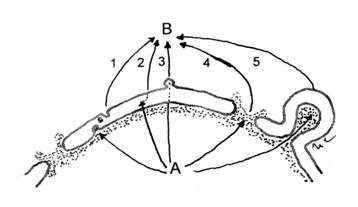
Fig.8. Diagrama cailor de translocare a matrialului din nucleu (A) in citoplasma (B).
1-Transport prin microvezicule; 2-Transport transmembranar3-Transport combinat;
4-Translocare prin
3.Nucleoplasma
Nucleoplasma, matricea nucleara sau sucul nuclear (nucleoplasma) reprezinta acea parte a nucleului, aparent lipsita de structuri (la microscopul optic si la cel electronic,de grosisment redus). La nivelul molecular apare structurata, incat denumirea de suc nuclear apare impropie, fiind mai adecvata denumirea de matrice nucleara. Nucleoplasma se prezinta ca un mediu de natura proteica care in care 'plutesc' nucleolii si cromatina. Are rol esential in organizarea nucleului, determinandu-i forma, in sinteza de ADN si ARN, in medierea semnalelor hormonale (ale steroizilor).Contine diverse enzime, ce intervin in glicoliza anaeroba si in realizarea unor legaturi macroergice.
Matricea nucleara cuprinde doua componente: matricea nucleara propiu-zisa si fractiunea labila a matricei.
Matricea nucleara propiu-zisa sau scheletul nuclear reprezinta echivalentul citoscheletului la nivel nuclear,fiind formata dintr-o retea de proteine stabile cu greutatea moleculara mare.Se poate izola dupa extragerea cromatinei si nucleolului. Este formata din trei componente: matricea fibrilara, componentele fibrilare (nemembranoase) ale invelisului nuclear si componenta fibrilara a nucleolului.
Matricea fibrilara apare ca o retea de fibrile intranucleare, intinsa in toata masa nucleului. Este formata din granule sau particule matriceale, electronodense, cu diametru de 15-20 nm si din fibrile matriceale (sau matrixin), cu diametru de 5 nm,dispuse in pachete. Matricea fibrilara contine proteine (20% din totalul proteinelor nucleare), ADN (1,2%), ARN (0,5%) si fosfolipide, in cantitate redusa. Lipsesc proteinenele histonice, in timp ce proteinele nonhistonice ocupa 18,2%, predominand cele acide.
Componentele fibrilare (nemembranoase) ale invelisului nuclear sunt o forma specializata a citoscheletului nuclear,fiind reprezentate de lamina densa interna (sau lamina fibrosa) si de complexul por.
Lamina densa interna este situata pe fata nucleara a membranei interne a invelisului nuclear. Are aspectul unei retele de fibrile legate pe de o parte la reteaua matricei nucleare si pe de alta parte la componentele fibrilare ale complexului por. Complexul por este format din structuri inelare (matricea anulara sau anulii), din granule centrale si din filamente radiare.
Lamina densa interna impreuna cu complexul por formeaza o componenta a scheletului nuclear, denumita complex lamina-por. Acest complex contine 2-3% din totalul proteinelor din nucleu (95% din ele fiind nehistonice, predominent acide). Pe fata interna a complexului lamina-por a fost identificata o nucleozid trifosfataza, implicata in transportul ARN catre citoplasma.
Componenta fibrilara nucleolara (matricea nucleolilor) este reprezentata de o retea de filamente ce ocupa aria ce corespunde partii fibroase si partii granuloase din nucleol.Este formata din polipeptide asemanatoare celor din scheletul nuclear si are rol de suport pentru depozitarea granulelor ribozomale.
Functiile matricei nucleare sunt: 1- mentine forma nucleului si stabilitatea acestuia in interfaza; 2-asigura flexibilitatea nucleului, fapt ce permite realizarea unor modificari structurale legate de organizarea cromatinei, replicarea ADN, transcriptia si transportul intranuclear de ARN; 3- contractilitatea, independenta de ATP, dar influentata de ionii bivalenti (Ca2+,Mg2+) ce permite realizarea unor variatii ale volumlui nuclear; 4- suport pentru depozitarea granulelor ribozomale.
Cromatina
Cromatina nu este o substanta chimica, ci o notiune biologica, fiind reprezentata de materialul intranuclear care se evidentiaza cu coloranti bazici. Este forma de existenta a cromozomilor in interfaza , in celula aflata intre doua diviziuni. Cromatina si cromozomii sunt doua forme de organizare ale unuia si aceluiasi material genetic (ADN-ul).
Compozitia chimica a cromatinei cuprinde ADN, proteine histonice, mici cantitati de ARN si proteine nehistonice.
Cantitatea de ADN din nucleu este constanta pentru o anumita specie si reprezinta genomul speciei respective. Este data de numarul si dimensiunile moleculelor de AND.
Histonele sunt proteine bazice, prezente numai in genomul eucariotelor. In functie de continutul in arginina si lizina exista cinci tipuri de histone, denumite H1, H2A, H2B, H3, H La randul sau fiecare tip prezinta mai multe subtipuri.
Histonele prezinta mai multe particularitati: 1- au masa moleculara mica; 2- sunt puternic bazice, datorita continutului mare (10-20%) de aminoacizi bazici (lizine si arginine); 3- sunt purtatoare de sarcini electrice pozitive, care le permit sa se lege de sarcinile negative ale gruparilor fosfat din ADN; 4-sunt proteine cu evolutie filogenetica moleculara definitiva.
Histonele indeplinesc un rol major in impachetarea ADN-ului in nucleu, in realizarea organizarii supramoleculare a ADN-ului sub forma de nucleozomi. In cursul ciclului celular, histonele sufera o serie de modificari chimice ( acetilari,fosforilari, metilari).
In spermatozoid, histonele
sunt inlocuite de protamine. Protaminele au o greutate moleculara foarte
mica (4200 daltoni), o lungime medie de numai 33 aminoacizi si sunt
foarte bazice, continand multa arginina. Inlocuirea
histonelor are loc in timpul trecerii de
la spermatida la spermatozoid, permitand realizarea unui inalt grad
de condensare si compactare a cromatinei, pentru impachetarea ADN-ului
intr-un volum foarte mic, cum este cel al capului spermatozoidului.
Fig.9. Aspecte ale cromatinei nucleare
I. La microscopul optic: 1-Cruste; 2-Granule; 3-Nevuri; 4-Nucleu hipercromatic;
5-Nucleu hipocromatic.
II. La microscopul electronic: 1- Cromatina periferica; 2-Cromatina insulara; 3- Cromatina asociata nucleolui; 4-Nucleol; 5-Membrana nucleara; 6-Spatiu perinuclear; 7-Pori.
Aspectele histologice ale cromatinei. Datorita continutului bogat in ADN,complexat cu histone, cromatina se coloreaza intens cu coloranti bazici (hematoxilina,albastru de tripan), putand prezenta aspecte variate: granule, gramezi neregulate, retele de filamente, corpusculi cromocentrici (sau cariozomi). Aspectul morfologic al cromatinei este asemanator in nucleii apartinand aceluiasi tip celular, dar variaza in functie de tipul celular si de stadiul functional,fiind un criteriu important pentru identificarea celulelor la microscopul optic.
1. Eucromatina
Eucromatina (euchromatinum) reprezinta cromatina functionala, activa, purtatoare de gene structurale, pe care se face transcriptia mesajului genetic. Poate fi de doua feluri: 1- eucromatina activa, pecare transcriptia se efectueaza continuu, asigurandu-se desfasurarea normala a vietii celulei; 2- eucromatina permisiva , potential activa, devenindactiva in momentul in care actioneaza semnale specific modulatoare (de exemplu: hormonii).
Reglarea conformatiei genetice se realizeaza pe eucromatina, prin interventia unor agenti biochimici, in principal proteine nehistonice, ce pot actiona ca represori sau derepresori, moduland transcriptibilitatea materialului genetic activ si nu structura acestuia.
Raportul dintre eucromatina si heterocromatina se exprima functional prin raportul dintre transcriptibil si netranscriptibil, permitand, in fapt, realizarea relatiei dintre genotip si fenotip. Astfel, prin genotip se intelege totalitatea materialului genetic cuprins in cromatina interfazica, deci eucromatina impreuna cu heterocromatina. Fenotipul reprezinta expresia unei parti din genotip, care a fost transcrisa din eucromatina in interfaza, totalitatea trasaturilor (insusirilor morfologice,fiziologice,etc.) exprimate din noianul de informatii inscrise in genotip.
2. Heterocromatina
Heterocromatina (heterochromatinum) este cromatina condensata, netranscriptibila, inactiva metabolic.Exista doua tipuri de heterocromatina: constitutiva si facultativa.
Heterocromatina constitutiva este genetic inactiva, lipsita de gene structurale. Pe ea nu se face transcriptie niciodata, ramanad intotdeuna condensata in interfaza. Contine ADN repetitiv sau satelit, un ADN in care anumite secvente nucleotidice sunt repetate de un numar foarte mare de ori. La mamifere, ADN-ul repetitiv reprezinta 15% din cantitatea totala de ADN. Exista doua tipuri de ADN repetitiv:1- ADN inalt repetitiv (10%), in care secventele se repeta de sute de mii de ori; 2-ADN mediu repetitiv (5 %), in care secventele se repeta de 100 ori, de 1000 ori sau de zeci de mii de ori.
Pe cromozomii omologi (care formeaza o pereche), heterocromatina constitutiva se localizeaza identic, de regula, in imediata vecinatate a centromerului, putand fi detectata prin tehnica de bandare a cromozomilor.
Rolul heterocromatinei constitutive este incomplet elucidat, atribuindu-se o semnificatie de 'protectie' sau de 'suport', sugerandu-se ipoteza ca ea determina specificitatea centromerica, respectiv pozitia centromerului in cromozomi.
Heterocromatina facultativa este o cromatina condensata, care contine gene structurale represate (inactive) pe care nu se face transcriptia. Pe aceste gene, transcriptia s-a efectuat sau nu intr-o perioada anterioara, sau se va putea efectua, daca se transforma in eucromatina, sub influenta unui agent derepresor. Astfel, transformarea unei parti din heterocromatina autozomala in eucromatina determina sinteza de imunoglobuline si transformarea limfoblastica a limfocitelor.
In functie de cromozomi in care se gaseste, heterocromatina poate fi: 1-autozomala (autosoma), prezenta in perechile de cromozomi autozomi, sub forma portiunilor condensate, heterocromatice, ce contin lanturi de gene, inactive transcriptional; 2-gonozomala (gonosoma), prezenta numai in cromozomii ce determina sexul , reprezentata de cromatina X, la femela si de corpusculul Y, la mascul.

Fig.10. Aspectele cromatinei X
1-Satelit nucleolar; 2-Drumstick; 3-Corpuscul cromocentric
Cromatina X sau corpusculul Barr a fost descrisa de M.L.BARR si de E.G.BERTRAM in 1949, in neuronii din nucleul nervului hipoglos la pisica. Este folosita ca marker genetic pentru recunoasterea sexului genetic. In mod normal este prezenta numai in nucleii celulelor somatice ale femelelor sub forma unui corpuscul cromatidian, denumit corpuscul Barr. Acesta reprezinta condensarea interfazica a unuia din cei doi gonozomi X. Prin condensare, eucromatina se transforma in heterocromatina facultativa inactiva. In situatia in care ambii gonozomi X ar fi activi (transcriptionabili), in celulele femelei ar trebui sa existe o cantitate dubla de enzime, controlate de genele situate pe cromozomul X.
Cromatina X este prezenta si detectabila la toate femelele mamiferelor, cu exceptia femelei de opussum (mamifer marsupial), unde este prezenta la ambele sexe. Lipseste la toti masculii normali.
Corpusculul Barr are un diametru de cca. 1 m si poate prezenta forme diferite: plan-convex, convex-concav, biconvex, triunghiular. Pozitia sa variaza dupa tipul celular, aparand situtat adiacent membranei nucleare interne ( in celulele epidermale si ale epiteliului mucoasei bucale) sau atasat nucleolului (in unii neuroni). In neutrofilele femelelor este prezenta o formatiune echivalenta corpusculului Barr, denumita, dupa aspectul sau 'bat de toba' (drumstick), formata dintr-un cap cu diametrul de 1,5 m, atasat printr-un filament cromatic extrem de subtire la unul din lobii nucleului. Frecventa drumstick-ului est de 7/500 neutrofile, sau chiar mai mult.
Cromatina Y sau corpusculul F se poate observa in nucleii celulelor provenite de la masculii normali, examinate in lumina ultravioleta, dupa colorare cu fluorocrom. Are aspectul unui corpuscul intens fluorescent. A fost descpoperit de PEARSON, in 1970 si reprezinta expresia citologica a cromozomului Y. La indivizii ce prezinta doi cromozomi Y se observa doi corpusculi F. In acest mod se pot diagnostica unele anomalii cromozomiale (trisomia XXY) la masculi, care predispun la un comportament dur, inrait.
3. Cromozomii
Cromozomii (Chromosoma) sunt structuri celulare cu numar, forme si marimi caracteristice pentru fiecare specie. Ei devin vizibili la microscopul optic in celulele care se pregatesc pentru diviziune sub forma unor filamente lungi, cu mare afinitate pentru coloranti bazici, de unde isi trag denumirea, croma insemnand in limba greaca culoare, iar soma - corp. (Fig.11.)
In functie de posibilitatile de a fi observati la microscopul optic, cromozomii pot fi clasificati in : - cromozomi interfazici, necondensati si neobservabili la microscop; -cromozomi mitotici, usor observabili la microscop, mai ales in cursul metafazei si de aceea denumiti cromozomi metafazici. Structura cromozomilor metafazici cuprinde: cromatidele, centromerul, kinetocorii, constrictiile secundare si satelitii.
Cromatidele (chromatidea) sunt cele doua jumatati longitudinale, genetic identice ce formeaza fiecare cromozom metafazic. Fiecare cromatida corespunde unei molecule de ADN.

Fig.11. Cromozomi interfazici si cromozomi mitotici
Extremitatile terminale ale cromatidelor se numesc telomere (telomerus) si sunt absolut necesare pentru structura si functionarea normala a cromozomilor, deoarece previn fuziunea si mentin o anumita ordine a cromozomilor interfazici in interiorul nucleului prin atasarea lor la membrana nucleara interna.
Centromerul (centromerus ) sau constrictia primara este regiunea cea mai ingusta din lungimea cromozomului, unde cele doua cromatide se unesc. Centromerul este slab colorat sau acromatic, avand un continut scazut de ADN. In vecinatatea centromerului se gaseste heterocromatina constitutiva. Cei mai multi cromozomi sunt monocentrici ( cromosoma monocentricum), prezentand un singur centromer. In cazul unor anomalii cromozomiale apar cromozomi dicentrici ( cromosoma dicentricum) sau policentrici (cromosoma polycentricum).
Kinetocorii (kinetocorus, i) sunt in numar de doi pentru un cromozom si reprezinta locul prin care fiecare cromatida se ataseaza de microtubulii fusului de diviziune. La microscopul electronic un kinetocor are forma unui oval cu dimensiunile de 5/25 nm. (fig.12.)
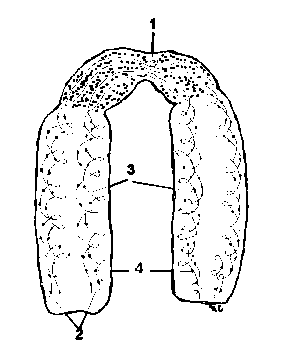
Fig.12.Ultrastructura cromozomului
1-Centromer; 2-Cromatide; 3-Cromoneme; 4-Cromomere.
Constrictiile secundare sunt zone ale cromozomilor ce nu retin colorantul bazic. Servesc drept criteriu morfologic de indiviudalizare a cromozomilor. In comparatie cu constrictiile primare, la nivelul costrictiilor secundare nuse produc deviatii angulare ale segmentelor pe care le unesc. Unele din zonele de constrictie secundara sunt legate de formarea nucleolilor si se numesc zone nucleolare sau organizatori nucleolari. iar cromozomii care participa la formarea nucleolilor sunt denumiti cromozomi nucleolari (chromosoma nucleolare).
Satelitul (satelles chromosomalis) este un corpuscul sferic, cu diametrul egal cu al cromozomului sau mai mic, atasat de restul cromozomului printr-un filament (filum satellitis) subtire de cromatina de lungime variabila. Cromozomii ce prezinta satelit se numesc SAT-cromozomi (chromosoma satellitiferum).
In functie de pozitia centromerului, cromozomii se clasifica in: 1-telocentrici (chromosoma telocentricum), la care centromerul este situat la o extremitate a cromozomului,lipsindu-i bratele scurte; 2-acrocentrici (chromosoma acrocentricum), cu brate scurte abia vizibile, centromerul fiind situat in apropierea extremitatii cromozomului;3-submetacentrici (chromosoma submetacentricum), la care centromerul este situat in apropierea mijlocului lungimii cromozomului, prezentand un brat scurt si un brat lung; 4- metacentrici (chromosoma metacentricum), cu bratele
aproximativ egale, centromerul fiind situat la mijlocul lungimii cromozomului.
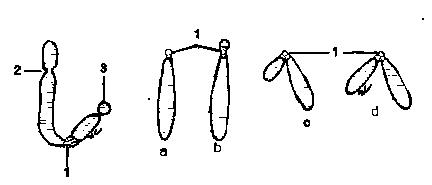
Fig.13. Schema unui cromozom.Tipuri de cromozomi.
1-Centromer: 2-Constrictie secundara; 3-Satelit;
a-Cromozom telocentric; b-Cromozom acrocentric;
c-Cromozom submetacentric;d-Cromozom metacentric.
Dimensiunile cromozomilor sunt cuprinse intre 1,5-10 m, lungime si 0,2-2 m diametru, putandu-se intalni cromozomi giganti si cromozomi pitici.
Numarul cromozomilor apare constant pentru aceiasi specie, existand: 60 cromozomi la bovine, 54 la ovine, 38 la suine, 64 la cabaline, 63 la catar, 38 la felide, 78 la canide si 46 la primate.
Celulele sexuale sau gametii sunt haploide (haplos= simplu), continand un singur set de cromozomi, notat cu 'n',fata de celulele somatice care sunt diploide (diplos=dublu) si contin doua seturi (2n) de cromozomi (un set matern si altul patern). Fiecare cromozom dintr-un set are in setul opus un cromozom complementar, identic structurat morfologic si genetic, alcatuind o pereche de cromozomi omologi. Asfel, taurinele prezinta 58 cromozomi autozomi (29 perechi) si 2 cromozomi de sex ( heterocromozomi sau gonozomi). Taurinele femele au in celulele somatice doi cromozomi X, ,iar formula cromozomiala este 60,XX. Masculii taurinelor au formula cromozomiala 60,XY si prezinta in celulele somatice un cromozom X si altul Y. In formula cromozomiala, prima cifra indica numarul total de cromozomi, dupa virgula fiind indicati cromozomii de sex (gonozomii). La taurine, fiecare din cele 29 perechi de cromozomi autosomi este formata din 2 cromozomi omologi, cu aceiasi morfologie si dispunere a genelor. Heterocromozomii ( gonozomii) nu sunt omologi. La taur, diferenta de forma intre cromozomul X si cel Y este evidenta, cromozomul Y fiind un cromozom submetacentric mic. La vaci, cei doi cromozomi X submetacentrici nu sunt omologi, deoarece numai unul este activ genetic, iar celalalt este inactiv, ramane condensat in interfaza si formeaza cromatina sexuala (corpusculul Barr). Diferentierea celor doi cromozomi X in unul activ si altul inert genetic se face intr-un stadiu precoce al dezvoltarii embrionare.
Compozitia chimica a cromozomilor include ADN, proteine (histonice si nonhistonice) si o cantitate redusa de ARN.
Histonele sunt proteine cu greutate moleculara redusa (10.000- 20.000 dal), cu continut mare (10-20%) de aminoacizi bazici (lizina si arginina), purtatori de sarcini electrice pozitive, ce le permite sa se lege de sarcinile negative ale gruparilor fosfat din ADN. La eucariote exista cinci clase de histone, ce difera dupa continutul lor in lizina si arginina: H1, H2A, H2B, H3 si H. In timp ce histonele H2A, H2B, H3 si H4 sunt cele mai constante din punct de vedere al secventei aminoacizilor la diverse specii, histonele H1 prezinta o variabilitate a secventelor.
Proteinele nonhistonice sunt de foarte multe tipuri, de la proteine ce intra in structura cromozomilor sau care se leaga de ADN, intervenind in exprimarea genelor si pana la proteine enzime ce intervin in sinteza de ADN si ARN sau in scindarea lor.
Proteinele ce intervin in reglarea exprimarii genelor sunt rare, aparand intr-un exemplar la 3000 de nucleozomi. Exista, insa si proteine nehistonice mult mai abundente, precum grupul de proteine cu mobilitate mare (HMG-higt mobility group), care fiind mici si cu multe sarcini electrice migreaza rapid in cursul electroforezei. Din acest grup, proteinele HMG14 si HMG17 se gasesc in toate celulele de la mamifere, legandu-se de nucleozomii asociati cu genele active.
ARN-ul se gaseste in cantitate redusa in cromozomi, fiind reprezentat de ARN-ul nascent, care se formeaza pe matrita de ADN in procesul de transcriere.
3.1. Cariotipul
Cariotipul este reprezentat de totalitatea caracterelor morfologice (numar, marime, forma, pozitia centromerului, constrictiile ) ale cromozomilordintr-o celula diploida. Grafic poate fi prezentat printr-o 'harta' numita cario- sau idiograma, in care perechile de cromozomi omologi sunt asezati in ordinea descrescanda a lungimii. Cromozomii sexuali sunt cromozomi X submetacentrici si cromozomul Y, submetacentric mic.
Cariotipul unei specii se obtine prin fotografierea cromozomilor metafazici, obtinuti din metafazele limfocitelor din culturi, stimulate sa se divida cu fitohemaglutinine si blocate in metafaza prin adaus de colchicina, un alcaloid ce impiedica asamblarea fusului de diviziune. Din fotografiile obtinute se decupeaza cromozomii si se aranjeaza in cariotip, putandu-se identifica grupele de cromozomi, fara a se putea preciza cu certitudine perechea.
Identificarea fiecarei perechi se face prin folosirea tehnicilor de bandare, tehnici care permit punerea in evidenta a unor benzi transversale de-a lungul cromozomilor, benzi care sunt caracteristice pentru fiecare cromozom.
Se folosesc doua categorii de tehnici de bandare: 1- tehnici cu fluorescenta prin care se obtin benzile 'Q' ( de la quinacrina) fluorescente; 2 -tehnici bazate pe tratarea cromozomilor cu diferiti agenti fizici si chimici, urmate de coloratia Giemsa prin care se evidentiaza trei feluri de benzi : benzile G (Giemsa), care apar intens colorate si corespund benzilor fluorescente Q; -benzile R (reverse band), cu o dispunere inversa benzilor G, incat benzile R intens colorate, corespund benzilor G palide;- benzile C, apar numai in regiunea centromerului sau in apropiere ei si corespund localizarii heterocromatinei constitutive.
Determinarea cariotipului are o mare importanta in practica medicala, pentru: a-diagnosticul unor boli congenitale, precum trisomia perechiii 21, care produce boala Down (idiotia mongoloida); b- sfatul genetic si dirijarea imperecherilor la animale; c-aprecierea statusului genetic la populatiile cu mare risc genetic; d-diagnosticul diferential al unor anomalii sexuale, cu determinare genetica, precum sindromul Turner (XO, in loc de XX), ce produce sterilitate, ovare vestigiale si sindromul Kleinfelter (XXX, in loc de XY), ce se manifesta prin testicule atrofiate, sterilitate, ginecomastie; e-diagnosticul unor boli ale sangelui, precum leucemia mieloida, unde apare un cromozom marker anormal (cromozomul Ph-Philadelphia); f - pentru diagnosticul de paternitate, prin masurarea lungimii cromozomului Y, care trebuie sa fie aceiasi la fat ca si la presupusul tata; g- pentru diagnosticul diferential intre unele tumori benigne si alte maligne, ce modifica cariotipul. De asemenea stabilirea locusului pe care il ocupa ogena intr-un cromozom permite realizarea hartilor cromozomiale, un domeniu de cercetare de mare actualitate.
3.2. Organizarea moleculara si supramoleculara a cromozomilor la eucariote.
La microcopul electronic, cromozomii metafazici apar ca niste gheme de cromatina, infasurate foarte neregulat. Fiecare cromatida contine o singura molecula de ADN ce se organizeaza sub forma unei fibre de cromatina, cu grosimea de 10 nm, foarte sinuoasa, impachetata compact, prezentand zone de eucromatina si heterocromatina. Impachetarea ADN-ului si a histonelor in cromozom este absolut necesara pentru ca o molecula de AND co greutate moleculara de 6 1010 daltoni si o lungime de 3 cm sa incapa intr-un cromozom lung de 5 m ( 0,0005 cm). Impachetarea se facecu ajutoul nucleozomilor, care sunt unitati fundamentale repetitive de organizare a complexului AND -histone, deci a cromatinei. (Fig.14).

Fig.1Modalitati de plicaturare a fibrei de cromatina in cromozom
1-Plicaturare transversala; 2-Plicaturare longiutdinala;
3-Plicaturare combinata; 4-Structura cuaternara.
3.3. Nucleozomii
Conceptul de nucleozom a fost elaborat de KRNBERG, in anii 1975-1977. Un nucleozom este un octamer histonic de forma unui cilindru scurt , cu un diametru de 11 nm si o inaltime de 5,5 nm, pe care duplexul de AND il inconjoara de doua ori.(Fig.15.)
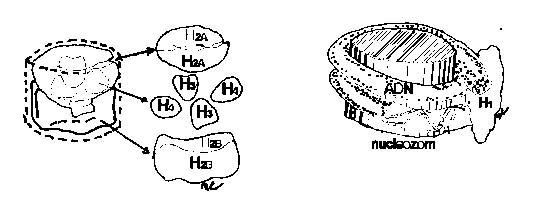
Fig.15.Alcatuirea nucleozomului Fig.16.Pozitia histonei H1
Octamerul histonic este format din opt molecule de histone, cate doua molecule de H2A, H2B, H3 si H Tetramerul de histone - (H3) 2 si (H4) 2 - bogate in arginina formeaza miezul nucleozomului si determina infasurarea duplexului de ADN. Cei doi dimeri de histone bogate in lizina, - (H2A) 2 si (H2B) 2 - sunt atasati miezului nucleozomului. Nucleozomul nu contine proteine nonhistonice.
Histona H1 nu intra in structura nucleozomului, fiind siutata lateral de acesta, in contact cu intrarea sau iesirea ADN-ului din nucleozom. Histona H1 se leaga de ADN-ul linker prin forte ionice, prezentand o portiune C (carboxil) terminala, legata direct de ADN si o portiune N (amino-), ce nu se ataseaza de ADN, dar ofera locusuri de legare pentru proteinele nehistonice HMG1 si HMG2. Histona H1 este implicata in spiralizarea Fibrei de cromatina.(Fig.16).
Molecula de ADN este continua de-a lungul cromozomului, prezetand portiuni infasurate pe nucleozomi si portiuni ce fac legatura dintre doi nucleozomi succesivi, denumite ADN-linker sau internucleozomic. De ADN-ul linker se leaga o molecula de histona H1, amplasata intre doi nucleozomi. Se realizeaza, asfel, o insiruire a nucleozomilor unul dupa altul, ca margele pe ata, formandu-se filamentul subtire de cromatina, cu diametrul de 11 nm, care se poate observa la microscopul electronic, numai daca cromatina este 'intinsa' in mod artificial. In celula vie, fibra de cromatina are un diametru de 30 nm, rasucita intr-o structura helicoidala (un solenoid sau o bobina) cu pasul elicei de 11 nm. La fiecare tur de spira se insera sase nucleozomi. Rata de imperechere a ADN-ului pe nucleozom este de circa 10/1. aproximativ 60 nm de ADN fiind infasurati pe un cilindru nucleozomic inalt de 5,5 nm. Apoi, dispunerea nucleozomilor sub forma de bobina (solenoid) cu diametru de 30 nm permite realizarea unui raport de impachetare de cca. 50/1. Eucromatina corespunde filamentului de cromatina de 10 nm, iar heterocromatina corespunde solenoidului (bobinei) de 30 nm.(Fig.17)

Fig.17.Formarea filamentului subtire de cromatina
A-Histona H1; B-Nucleozom; C-Filamentul de cromatina;
1-Portiunea globulara a histonei.
Impachetarea duplexului de ADN pe nucleozom si a nucleozomilor in fibra de cromatina de 30 nm ar permite ca intr-un cromozom ipotetic de 1 mm lungime, sa incapa un ADN lung de 3 cm. La randul ei, fibra de cromatina de 30 nm se pliaza formand bucle de marimi variabile, ce permite sa se reduca lungimea 'firului' de ADN de la 1 mm la 100 m.
Pentru a 'incape' in cei 5 m lungime ai unui cromozom metafazic sunt necesare inca doua ordine de condensare a cromatinei, care se realizeaza probabil prin impachetarea elicoidala a buclelor de cromatina, incat aglomerarea lor in anumite zone determina aparitia benzilor vizibile in cromozomul metafazic. Se pare ca superspiralizarea solenoidului se realizeaza cu ajutorul unor proteine nehistonice. Astfel, fosforilarea proteinelor nehistonice ar declansa superspiralizarea solenoidului, in momentul trecerii de la G2 la mitoza, iar defosforilarea lor ar induce despiralizarea, in momentul trecerii de la mitoza la G1. Alaturi de histone, in procesele de impachetare a nucleozomilor in fibra de cromatina, intervine ionul de Mg2+.(Fig.18.)
|
|
Fig.18.Structura fibrei de cromatina de 30 nm (A) si nucleozomului (B).
1- Miezul histonic; 2- ADN linker.
In timpul diviziunii celulare, condensa-rea cromatinei corespunde impacheta-rii compacte in spatiu a fibrei de cromatina de 30 nm, odata cu realizarea structurii cromozomului metafazic. (Fig.19.)
S-a calculat ca, ADN-ul infasurat pe un nucleozom este prea scurt pentru a corespunde unei gene structurale, incat nu exista o concordanta intre o gena, ca unitate de informatie genetica si un nucleozom, ca unitate de impachetare a ADN-ului.
Gena este formata din aproximativ 1000 perechi de nucleotide (perechi de baze), in timp ce ADN-ul infasurat pe un nucleozom are 200 perechi de baze. Unele gene de la eucariote apar discontinui, prezentand secventele din ADN care codifica aminoacizi (exoni), separate intre ele prin secvente de ADN care nu codifica aminoacizi (introni).(fig. 19)


Fig.19.Eucromatina si heterocromatina Fig.20.Etapele realizarii structurilor cromozomilor eucariotici
1-ADN linker A-Duplex de ADN;
1+2-Eucromatina; B-Filament de cromatina;
3-Solenoid de 30 nm - heterocromatina C-Fibra de cromatina;
D-Bucle de cromatina;
E-Banda condensata;
F-Cromozom in metafaza.
Exista mai multe ipoteze asupra mecanismului prin care genele structurale din ADN-ul infasurat pe nucleozom sunt activate pentru transcriptie. Cea mai larg acceptata ipoteza (emisa in 1982) sustine ca activarea genelor se face ca urmare a interventiei proteinelor nehistonice HMG14 si HMG17, care fie ca inlocuiesc proteinele HMG1 si HMG2 de pe locurile lor de legare pe histona H1,fie ca inlocuiesc insasi histona H1 de pe ADN-ul linker internucleozomic.(Fig.20.)
In acest fel, regiunile din filamentul nucleozomic in care intervin proteinele HMG14 si HMG17 isi vor modifica conformatia biochimica, devenind regiuni active transcriptional.
5. Nucleolul
Nucleolul (nucleolus) este o componenta intranucleara, cu aspect corpuscular (corpusculum nucleare), prezenta in interfaza, a carei functie principala consta in biogeneza ribozomilor (cu exceptia ribozomilor mitocondriali. Ocupa o pozitie cheie in circuitul intracelular al informatiei, fiind sediul unui considerabil trafic de molecule. In el se desfasoara principalele procese care au loc in nucleu (replicare, transcriptie, transport), jucand rolul unui intermediar intre cromozomi si citoplasma. Este prezent (vizibil sau nu ) in toate celulele eucariote. Nu exista nucleu fara nucleol, iar celulele care isi perd nucleolii nu mai sunt viabile.
In timpul diviziunii celulare se dezintegreaza si reapare dupa terminarea acesteia. In structurarea nucleolului, un rol deosebit revineorganizatorilor nucleolari, prezenti in unii cromozomi, adiacent constrictiei secundare.
Desi a fost observat inca din 1836 de catre VALENTIN, ca o granula densa in interiorul nucleului, rolul sau a fost dezlegat mult mai tarziu, abia in 1960, precizandu-se rolul esential pe care il are in biogeneza ribozomilor.
Examinat la microscopul prezinta un aspect omogen, puternic bazofil, bine delimitat. Prin impregnari argentice, au fost evidentiate doua componente in structura nucleolului: 1-nucleolonema (nucleolonema), o formatiune filamentoasa, rasucita ca un ghem; si 2- o componenta astructurata sau amorfa (pars amorpha). Nu este delimitat de o membrana.
In celulele vii, examinate in contrast de faza, nucleolul apare ca un corpuscul puternic refringent, datorita continutului redus de apa, heterogen si cu contur neregulat.
Numarul nucleolilor. In nucleii celulelor somatice exista, in mod obisnuit doi nucleoli, cate unul pentru ,fiecare set cromozomial. Numarul nucleolilor creste in raport direct cu gradul de poliploidie. Astfel, in hepatocite exista 2 nucleoli, in celulele diploide, 4 in cele tetraploide si 8 in cele octaploide.
Dimesiunile nucleolilor sunt intre 1-2 m, ocupand circa 30% din volumul nucleului. Ele variaza in functie de implicarea celulelor in sinteza de proteine. Celulele care sintetizeaza cantitati mari de proteine (neuronii, celulele embrionare, limfocitele stimulate antigenic) au nevoie de multi ribozomi,ceea ce face ca nucleolii sa aiba dimensiuni mari. In celule in care sinteza de proteine este redusa (in spermatozoizi), nucleolii au dimensiuni mai mici. Nucleolul apare hipertrofiat in anabolism, in perioada de diferentiere embrionara si se reduce in catabolism si in inanitie.
Raportul nucleolo / nuclear. Numarul si volumul nucleolilor depind de starea functionala a celulei. Cu cat celula este mai activa in sinteza proteinelor, cu atat raportul nucleolo/nuclear este mai mare. Acest raport este folosit drept criteriu pentru aprecierea varstei celulelor. Astfel celula tanara prezinta un nucleu mare eucromatic, cu un nucleol mare, cu o citoplasma redusa si bazofila, iar o celula adulta prezinta un nucleu mai mic, hipercrom, cu nucleoli mici si o citoplasma abundenta.
|
|
Fig.21. Forma si pozitia nucleolului
A-Forma: 1-Sferoidal-ovoidala; 2-Discoidala; 3-Triunghiulara; 4-Neregulata.
B-Pozitia: 1-Alipit membranei nucleolare; 2-Central; 3-Excentric.
Forma nucleolilor. Nucleolii prezinta, de cele mai multe ori, o forma sferic-ovoidala, care pe masura imbatranirii celulei devine neregulata.
Densitatea nucleolilor este mare (1,35), nucleolul fiind cea mai densa structura din celula, datorita concentratiei ridicate in substanta uscata si a cantitatii foarte mici de apa.
Dispunerea nucleolilor in nucleu este in majoritatea cazurilor centrala sau paracentrala, putand varia in raport cu mometul functional. S-au observat si descris miscari ale nucleolului in interiorul nucleului si stabilirea unor contacte cu invelisul nuclear, localizare eficienta pentru descarcarea de material biologic in citoplasma.
5.1. Ultrastructura nucleolului
Ultrastructura nucleolului evidentiaza patru componente in alcatuirea nucleolului: componenta filamentara (pars filamentosa), componenta granulara (pars granulosa), componenta cromozomiala (pars cromosoma) si componenta astructurata (pars amorpha).
Componenta filamentara cuprinde filamente de 5nm grosime si 30-40nm lungime, dispuse in pachete intretaiate, formand o retea. Contine ADN-ul pe care se sintetizeaza ARN-ul ribozomal si produsul primar al acestei sinteze (ARN de 45 S). (Fig.22.)
Componenta granulara este domi-nanta, fiindalcatuita din granule cu diametru de 15-20 nm, asemanatoare, dar nu identice cu ribozomii citoplasma-tici. Aceste granule reprezinta precursorii ribozomilor.
Componenta cromozomiala, dispusa la periferia nucleolului (cromatina perinucleolara) sau avansand spre interiorul nucleolului sub forma de benzi (cromatina intranucleolara) este alcatuita din filamente de 10 nm.
|
|
Fig.22.Ultrastructura nucleolului.
1-Componenta filamentara;
2-Componenta granulara;
3-Componenta cromozomiala;
4-Componenta astructurata.
Componenta astructurata apare omogena si cu densitate medie la fluxul de electroni. Umple spatiul dintre granule si fibre, fiind considerata, de unii autori, cariolimfa, iar de alti autori un gen de matrice pentru celelalte componente nucleolare.
Cele patru componente nucleolare se pot distinge in acelasi nucleol numai in cazuri rare. Raporturile cantitative si topografice dintre ele variaza in functie de tipul celulei si de momentul functional al acesteia. Asfel, in hepatocitul uman, componentei filamentoase ii revine 15 %, celei granulare 70%, iar componentei cromozomiale 5% din volumul nucleolului. In celulele active, componenetele nucleolare pot segrega, dand nastere unui corpuscul nucleolar (corpusculum nucleare).
Nucleolul nu contine membrane si nu este delimitat de membrane.Componenta filamentoasa si ce granulara se pot asocia formand benzi ce apar, la microscopul optic, ca nucleolema. iar portiunea periferica a componentei cromozomiale corespunde cromatinei asociate nucleolului.
In functie de criteriile ultrastructurale, exista mai multe tipuri de nucleoli: nucleoli reticulari, cei mai comuni, cu cele patru componente distincte; nucleoli compacti, in care nu se disting componentele, intilniti la cateva tipuri celulare, ca de exemplu la limfocite; nucleoli inelari, in care copmponenetele filamentoasa si granulara formeaza un inel periferic, care inconjoara o lacuna centrala.
Compozitia chimica a nucleolului variaza in functie de tipul celular si de momentul functional, principalele componente chimice fiind: ADN-ul (3 %), ARN-ul (7%) si proteinele (90% din greutatea uscata). La aceasta se mai adauga cantitati mici foarte mici de lipide si minerale (magneziu, calciu, zinc, cobalt, etc.).
ADN-ul din nucleol este reprezentat de organizatorii nucleolari ce patrund ca niste bucle de ADN in nucleol.
ARN-ul din nucleol este in principal ARN ribozomal aflat in diverse faza de maturare. Se presupune ca nucleolul este o statie intermediara obligatorie in tranzitul spre citoplasma al ARN-ului mesager si al celui de transfer.In nucleol exista diferite tipuri de ARN, ce difera dupa coeficentul lor de sedimentare (exprimat in unitati Svedberg), precum ARN 45 S, ARN 41 S, ARN 20 S, ARN 28 S, ARN 32 S. Aceste tipuri corespund diferitelor etape de maturare a ARN-lui.
Proteinele nucleolare provin din citoplasma si sunt repezentate in cea mai mare parte de enzime implicate in sinteza si maturarea ARN r (ribozomal), precum ARN-polimeraza-ADN dependenta (sau ARN-polimeraza I), convertaza, metilaza,etc.
Bazofilia nucleolului reprezinta principala sa caracteristica, vizibila la microscopul optic si se datoreste continutului relativ mare de ARN si AND.
5.2.Biogeneza nucleolilor
Nucleolul este vizibil numai in interfaza,disparand in profaza si reaparand in interfaza. Un rol esential in biogeneza nucleolului ii revine organizatorului nucleolar,care este o zona cromozomiala distincta, slab colarabila (heteropicnoza negativa), situata in vecinatatea constrictiei secundare, avand un grad mai redus de infasurare a fibrei de cromatina. Numarul cromozomilor care prezinta organizatori nucleolari este limitat si variabil in functie de specie. La hominide exista cinci perechi de cromozomi autuzomi, purtatori de organizatori nucleolari (perechile 13, 14, 15,21 si 22 ). (Fig.23.)
La microscopul optic, organizatorii nucleolari corespund cromatinei asociata nucleolului, ce poate fi evidentiata prin reactia Feulgen, aparand colorata rosu-violaceu. La microscopul electronic, organizatorii nucleolari corespund componentei cromozomiale, iar functional cu ADN r (genele care codifica ARN r) ce cuprinde cistronii responsabili de transcriptia ARN r.
|
|
Fig.23. Organizatorii nucleolari.
1-Invelisul nuclear; 2-Nucleol; 3- Cromozomi cu oranizatori nucleolari; 4-Bucle de cromatina cu gene pentru ARN r.
Ipotezele clasice privind biogeneza nucleolului sustin: a- continuitatea sau persistenta nucleolului in cursul diviziunii celulare sub forma de granule sau filamente fine (denumite corpusculi nucleolari), atasate de cromozomi, impreuna cu care se repartizeaza in celulele fiice, dupa care sunt asamblati de organizatorii nucleolari; b-formarea de novo, deoarece materialul nucleolar se dezintegreaza complet in cursul mitozei, fara a fi incorporat in noii nucleoli.
Conceptia actuala a realizat un compromis rational din punct de vedere molecular, intre ipotezele clasice (a continuitatii si a formarii de novo), in sensul ca componeneta cromozomala respecta continuitatea, iar celelalte componente (filamentoasa, granulara si amorfa) sunt neoformate rapid in faza G1 a ciclului celular, cand cistronii sunt reactivati pentru transcriptie.
5.3. Functiile nucleolului
Nucleolul indeplineste functii legate de: - sinteza ARN r si biogeneza ribozomilor; -reglarea sintezei de ARN r; - transferul ARNm si ARNs in citoplasma; pregatirea mitozei.
In sinteza de ARNr, ADN-ul nucleolar joaca rolul de matrita, reprezentand organizatorul nucleolar. ADN nucleolar contine cistroni, denumiti ADNr, responsabili pentru transcriptia ARNr, separati intre ei prin segmente de spatiere si terminare, formate din ADN care nu codifica. ADNr este transcris prin interventia ARN-polimerazei I, rezultand un ARN de 45 S care reprezinta precursorul ARN-ribozomal. In nucleolii hepatocitelor se sintetizeaza 1000 de molecule de ARN-45S pe minut. O parte din molecula de ARN de 45 S este indepartata de o endonucleaza si

apare un ARN de 41 S, care prin
interventia unei convertaze este
scindat in ARN de 20 S si ARN de 32 S. (Fig.2)
Fig.2Rolul nucleolului in sinteza de ARNr si in biogeneza ribozomilor
ARN-ul de 20 S sufera interventia unei metilaze, care indeparteaza un fragment molecular, transformandu-se in forma matura de 18 S. ARN-ul de 18 S paraseste nucleolul si se combina cu proteine venite din citoplasma, dupa care este trecut in citoplasma prin porii membranei nucleare sub forma de particule ribonucleoproteice, care reprezinta subunitatea ribozomala mica de 40 S, ce apare in citoplasma la 30 minute de la inceperea sintezei.
ARN-ul de 32 S este si el metilat si se transforma in forma
matura de ARN de 28 S, dupa ce a pierdut un fragment din molecula. ARN-ul de 28 S impreuna cu ARN-ul de 5 S ( de origine nucleara, dar extranucleolara) si cu proteine formeaza o particula ribonucleoproteica, care reprezinta subunitatea ribozomala mare (de 60 S). Subunitatea ribozomala mare trece prin porii invelisului nuclear, aparand in citoplasma la o ora de la debutul sintezei. Cele doua subunitati ribozomale se ataseaza de ARNm, in momentul inceperii sintezei de proteine, formand ribozomii. Fibrilele din componenta filamentoasa sunt filamente de ARN de 45 S, iar granulele din componenta granulara sunt subunitati ribozomale mari.
Biogeneza ribozomilor
|
Etape |
Intervin: |
Rezultat |
|
Copierea cistronilor ADNr |
ARN-polimeraza I |
Sinteza de ARN 45 S |
|
Prelucrarea ARN 45 S |
Endonucleaza |
ARN 41 S |
|
Prelucrarea ARN 41S |
Convertaze |
ARN 20 S ARN 32 S |
|
Prelucrare ARN 20 S |
Metilaze |
ARN 18 S |
|
Prelucrare ARN 32 S |
Metilaze |
ARN 28 S |
|
Prelucrare ARN 18 S |
Proteine ribozomale |
Subunitatea ribozomala mica 40 S |
|
Prelucrare ARN 28 S |
Proteine ribozomale |
Subunitatea ribozomala mare 60 S |
Reglarea sintezei de ARNr in nucleol se realizeaza printr-un mecanism de feed-back. Existeta unui exces de ribozomii actioneaza ca inhibitor al genelor ADN r, care determina sinteza de ARNr. Invers, distrugerea ribozomilor va declansa incetarea inhibitiei si cresterea sintezei de ARNr, cu formarea de noi ribozomi.
Transferul de ARN mesager (ARN m), deci a mesajului genetic si de ARN de transport (ARN t) din nucleu in citoplasma nu poate avea loc decat in prezenta unui nucleol functional, fapt demonstrat experimental rpin distrugerea selectiva anucleolului cu un fascicul laser. Se crede ca nucleolul joaca rolul de statie intermediara in tranzitul ARN m-ului si ARN t-ului spre citoplasma.
Pregatirea si desfasurarea mitozei nu pote avea loc fara prezenta nucleolului in interfaza. Distrugerea acestuia produce blocarea celulei in faza G2, care precede mitoza.
5. Implicarea nucleolului in citopatologie.
Aspectul nucleolului poate fi folosit pentru recunoasterea celulelor canceroase. In celulele canceroase, nucleolul prezinta modificari de volum,numar si forma, care sunt o consecinta a malignizarii si nu cauza acesteia.
Hipertrofia nucleolara apare ca o caracteristica aproape constanta in celulele neoplazice (canceroase),iar cresterea numarului de nucleoli si neregularitatile de forma sunt relativ frcvente. Hipertrofia nucleolara nu este acompaniata si de o hipertrofie proportionala a nucleului incat valoarea raportului nucleolo-nuclear este mai ridicat in celulele neoplazice fata de cele normale (depasind valoarea de 1/3). Gradul de hipertrofie a nucleolului reflecta capacitatea proliferativa a celulei fiind proportional cu gradul de malignitate a tumorii.
Numarul nucleolilor este mai mare in celulele maligne decat in celulele normale, fiind uneori proportional cu gradul de malignitate a tumorii. Asfel, in unele adenocarcinoame din ovar, au putut fi observati 24 nucleoli intr-o celula.
Forma nucleolilor este neregulata in celulele maligne, ea putand varia de la o celula la alta in cadrul aceleiasi tumori.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4155
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved