| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
I.Organizarea organismului viu
Conform conceptiei sistemice elaborata de Ludwig von Bertalanfy (1901-1972), organismul viu este un sistem deschis adica un ansamblu de elemente in interactiune, care efectueaza, in permanenta, schimburi metabolice si energetice cu mediul inconjurator. Intreaga materie vie este organizata ierarhic, adica orice sistem este constituit din subsisteme si, la randul lui, participa ca subsistem la alcatuirea unui sistem de ordin superior (mai complex).
Sistemele echivalente din punct de vedere al modului lor de organizare formeaza un nivel de organizare a materiei. In ceea ce priveste materia vie, aceasta are patru niveluri de organizare: individual, populational (al speciei), biocenotic si al biosferei. "Forma elementara si totodata universala de existenta si deci de organizare a materiei, este organismul individual".
Un organism pluricelular este alcatuit dintr-o serie de sisteme mai simple (subsisteme), de complexitate crescanda (ierarhizate), dintre care unele sunt nebiologice (atomi, molecule, macromolecule), iar altele biologice (organite celulare, celule, tesuturi, organe si sisteme de organe). Nici unul dintre aceste sisteme nu poate exista de sine statator.
Toate organismele vii au o compozitie chimica relativ unitara, fiind constituite din:
macroelemente - in numar de 11, ce reprezinta 99,9% din greutatea corpului: C, O, H, N, S, P, Cl, Na, K, Ca, Mg
microelemente - in numar de circa 25, ce reprezinta numai 0,1% din greutatea corpului: Fe, Cu, Mn, I, Zn, Co etc.; unele dintre ele sunt de importanta vitala, altele par (cel putin pana in prezent) lipsite de importanta si ajung in organism mai mult sau mai putin intamplator.
Componenta principala a tuturor organismelor este apa. Continutul in apa al organismelor variaza: la om 60%, la soarece 67%, rac de apa dulce 74% la meduza 96%- dupa Penzlin, 1980). Ea serveste ca solvent si ca mijloc de transport. La vertebrate circa 65-75% din cantitatea de apa se gaseste in interiorul celulelor (restul de 25-35% se afla in spatiul extracelular). Impreuna cu substantele minerale (care se gasesc, in cea mai mare parte, sub forma ionizata in solutie apoasa), apa formeaza partea anorganica a structurii organismului. In organismul animal se gaseste un numar foarte mare de compusi organici, formati in special din primele patru macroelemente (C, O, H, N), majoritatea covarsitoare a acestora apartinand proteinelor, lipidelor, glucidelor si acizilor nucleici.
Fiind un sistem deschis, organismul viu intretine permanent un schimb de substante si de energie cu mediul inconjurator. Marele fiziolog german Johannes Mller (1801-1858) afirma: "atata timp cat un organism traieste, el se gaseste in permanenta descompunere, iar materia consumata este mereu inlocuita cu una noua". Totalitatea transformarilor de substante si energie care au loc in organismul viu se numeste metabolism.
Metabolismul are doua laturi opuse si, totodata, indisolubil legate intre ele: asimilatia (anabolismul) si dezasimilatia (catabolismul).
Asimilatia (lat. assimilare - a face asemanator) cuprinde ansamblul transformarilor din organismul viu care duc la sinteza de molecule si macromolecule organice specifice organismului respectiv din substante straine, nespecifice. Prin procesele de asimilatie, in care se consuma energie (procese endergonice), se realizeaza autorefacerea protoplasmei vii (sinteza ei), care este extrem de specifica (exista numeroase tipuri de protoplasma). De exemplu, protoplasma unui muschi, a unui nerv sau a unei glande se deosebesc radical una de alta, deoarece prima indeplineste functia de contractie, a doua de propagare si a treia de elaborare a unor secretii.
Dezasimilatia (lat. dissimilare - a face diferit) cuprinde totalitatea proceselor de descompunere a substantelor mai complexe (in special a proteinelor) in substante mai simple, care sunt eliminate in mediu (produsi de descompunere sau deseuri metabolice). In celulele majoritatii animalelor substantele organice sunt descompuse prin oxidare de catre oxigenul molecular, care patrunde in organism din mediul extern. In procesele de dezasimilatie se elibereaza energie (procese exergonice) - eliberarea energiei chimice potentiale din glucide, lipide si proteine (existente nu numai in hrana ingerata ci si in protoplasma insasi), energie care este utilizata de protoplasma pentru indeplinirea functiilor sale.
Energia potentiala a substantelor organice complexe, eliberata prin descompunere, este transformata in diferite forme de energie: calorica, mecanica, electrica si chiar luminoasa (la unele organisme). Forma dominanta este energia calorica (cantitatea acesteia la mamifere reprezinta 80% din energia eliberata si este utilizata pentru mentinerea temperaturii corporale adecvate). In protoplasma musculara, o parte a energiei eliberate apare sub forma de energie mecanica, reclamata de efectuarea diferitelor activitati motoare - miscarea unei parti a corpului sau a intregului corp, transportul hranei, al sangelui de la o regiune a corpului la alta etc. Energia electrica, desi eliberata in cantitate mica, este de importanta esentiala pentru functionarea sistemului nervos.
Cele doua laturi metabolice nu numai ca sunt inseparabil legate intre ele, dar, adesea este dificil sa se decida daca anumite procese biochimice (transfosforilarea, transaminarea etc.) trebuie sa fie considerate ca procese de asimilatie sau de dezasimilatie. Aceasta legatura foarte stransa se bazeaza pe faptul ca cele doua procese se intretin reciproc. Astfel, dezasimilatia intretine asimilatia prin energia rezultata din descompunerea oxidativa a moleculelor organice din structura protoplasmei sau din compusii micromoleculari reutilizabili (produsi ai metabolismului intermediar). La randul ei, asimilatia intretine dezasimilatia prin refacerea continua a structurilor protoplasmatice. In general, intensificarea proceselor anabolice este insotita de intensificarea proceselor catabolice, asa cum se intampla in timpul cresterii organismului si inmultirii celulelor, cand are loc o intensa sinteza proteica. De asemenea, procesele de dezasimilatie se intensifica foarte mult in timpul cresterii rapide a unei tumori maligne (cand se formeaza numeroase celule noi).
Procesele de asimilatie si cele de dezasimilatie nu sunt totdeauna reciproc echilibrate. Astfel, in timpul cresterii unui organism, desi se intensifica ambele procese, exista o preponderenta relativa a asimilatiei. Cresterea organismului poate sa rezulte fie din cresterea numarului de celule, fie din cresterea cantitatii de citoplasma in celulele deja existente (neinsotita de cresterea numarului lor).
Chiar si la un organism adult, in anumite imprejurari normale, anabolismul unui organ sau al unui tesut poate fi superior catabolismului sau, in decursul unei perioade limitate. Ca urmare, creste cantitatea de protoplasma din organul sau tesutul respectiv, producandu-se asa numita hipertrofie (este bine cunoscuta hipertrofia muschilor unui atlet, precum si hipertrofia inimii, induse de un travaliu suplimentar). Atunci cand predomina catabolismul se produce atrofia organului sau a tesutului.
Nu numai cresterea, ci si toate celelalte functii fiziologice sunt asociate cu metabolismul: dezvoltarea, reproducerea, digestia, respiratia, excretia, reactiile la modificarile mediului etc. Procesele metabolice stau la baza tuturor functiilor celulelor, tesuturilor, organelor si organismului ca intreg.
Deoarece multe functii importante ale organismului sunt asociate cu procese metabolice cantitativ neinsemnate, evaluarea acestora reclama utilizarea unor metode de investigatie extrem de sensibile si precise. S-a reusit marirea sensibilitatii metodelor electrice de masurare a temperaturii astfel incat a devenit posibila determinarea productiei de caldura intr-o fibra nervoasa, in timpul transmiterii unui singur impuls nervos (aceasta productie este extrem de mica, fiind capabila sa ridice temperatura cu numai 2 x 10-6sC - doua milionimi de grad). De asemenea, tehnicile chimice permit stabilirea structurii unor substante care se formeaza in organism in cantitati extrem de mici si sunt active in concentratii neinsemnate (1 x x10-8).
Dupa cum se stie, orice sistem tinde spre un echilibru termodinamic cu mediul sau. In cazul organismelor vii aceasta tendinta este anulata de procesele metabolice. Datorita fluxului permanent de substante si energie prin organismul viu, intre acesta si mediu se stabileste un alt fel de echilibru, care a fost numit "echilibru curgator" sau "echilibru dinamic" In absenta aportului de substante nutritive aceasta stare inceteaza si organismul moare. Cadavrul ajunge in stare de echilibru termodinamic cu mediul.
Spre deosebire de sistemele nebiologice, care evolueaza in sensul cresterii entropiei (masura a gradului de dezorganizare dintr-un sistem), sistemele biologice, datorita echilibrului dinamic, au un comportament antientropic, adica tind spre micsorarea entropiei. Cu toate acestea, cel de-al doilea principiu al termodinamicii (principiul entropiei) nu-si pierde valabilitatea, deoarece entropia sistemului "organism - mediu" creste.
La temperaturi compatibile cu viata, structurile organismului viu, desi in conditii termolabile, se conserva pana la moarte. Dimpotriva, partile componente ale unei masini trebuie sa fie termostabile pentru a nu fi distruse (prin topire, evaporare, indoire).
O data cu schimburile de substante dintre organism si mediul ambiant (metabolismul material) au loc si schimburi de energie, a caror totalitate reprezinta metabolismul energetic.
La organismele animale trebuie sa se ia in considerare patru feluri de energie: chimica, electrica, mecanica si termica. Pentru activitatea lor, celulele animale utilizeaza energia rezultata din reactiile chimice, realizand trei tipuri de travaliu: mecanic, electric si chimic. O parte din energie apare insa totdeauna sub forma de caldura. Intrucat cantitatea totala de energie din sistem nu se modifica, cu cat productia totala de caldura este mai mare, cu atat travaliul este mai mic (deoarece caldura este pierduta pentru celula, organismul nedispunand de un mecanism capabil sa utilizeze energia termica, organismul nefiind o masina termica). Pentru mentinerea functiilor vitale el trebuie sa recurga la energia chimica din substantele nutritive.
Sursele de energie ale organismelor animale sunt:
Compusii macroergici (in special ATP) care sunt in prealabil sintetizati si depozitati in celule; acestia reprezinta sursa imediata de energie pentru orice reactie celulara.
Substantele alimentare (introduse din afara) furnizeaza energie care inlocuieste energia din depozite, dupa ce aceasta s-s epuizat. Energia din alimente si cea din substantele depozitate sunt utilizate prin aceleasi reactii chimice.
Radiatiile solare sunt prima sursa a oricarei energii la organismele vii (biochimistul francez Polonovski, 1948: "asimilatia clorofiliana este baza intregii energii potentiale, pe care regnul vegetal o ofera lumii animale"). Pentru a sintetiza 1 g de amidon plecand de la apa si bioxid de carbon, planta verde imprumuta de la razele solare o cantitate de energie calorica de circa 4 Kcal. Atunci cand aceeasi cantitate de amidon este degradata pana la apa si bioxid de carbon, in cursul proceselor metabolice din organismul animal, se elibereaza aceeasi cantitate de caldura.
Cea mai mare parte din energia eliberata in cursul proceselor metabolice este transformata in caldura. In repaus si pe nemancate schimburile energetice se manifesta sub forma producerii de caldura (ce rezulta din oxidarea substantelor nutritive). Prin oxidare, glucidele si lipidele sunt descompuse pana la bioxid de carbon si apa. Din descompunerea oxidativa a proteinelor rezulta si produsi finali care contin azot.
Oxidarea biologica nu este identica cu arderea asa cum s-a considerat in trecut, pe baza cercetarilor lui Lavoisier (1789). Desi in ambele cazuri termenii ecuatiei brute sunt aceiasi si se elibereaza aceeasi cantitate de caldura, cele doua fenomene sunt esential diferite. Astfel, in timp ce arderea este caracterizata prin degajarea unei mari cantitati de caldura si a unei temperaturi ridicate, oxidarea biologica se desfasoara la temperaturi mult mai scazute (la om, de exemplu la 37sC) iar energia rezultata apare numai partial sub forma de caldura, restul ramanand sub forma de energie chimica. De asemenea, spre deosebire de ardere, oxidarea biologica se realizeaza progresiv si in etape, prin transferul de electroni si ioni de hidrogen de la un compus intermediar la altul.
Pentru a putea evalua schimburile energetice trebuie cunoscute atat aportul cat si cheltuiala de energie ale organismului considerat.
Aceasta determinare poate fi facuta prin trei metode: metoda bilanturilor nutritive, metoda bazata pe caldura de combustie a alimentelor simple si metoda bazata pe analiza excretei.
1.Metoda bilanturilor nutritive - la un subiect aflat in echilibru alimentar (care nici nu pierde si nici nu castiga in greutate) se masoara cantitatea de energie calorica cuprinsa in ingesta (in alimentele ingerate) si in excreta (in substantele eliminate in mediu). Diferenta dintre cele doua valori reprezinta cantitatea de energie calorica degajata de organism pe durata experientei.
Deoarece determinarea energiei din excreta necesita operatii dificile, in practica curenta se determina numai valoarea calorica a ratiei alimentare care asigura starea normala a subiectului, fara sa se mai masoare pierderea prin excreta, care este considerata ca reprezentand, in medie, 10% din valoarea calorica a alimentelor ingerate.
2. Metoda bazata pe caldura de combustie a alimentelor simple - consta in calcularea cantitatii de energie furnizata organismului, cunoscandu-se caldura de combustie in organism a acestora, precum si proportiile in care acestea se gasesc in ratie.
Determinarea caldurii de combustie (a valorii calorigene) a alimentelor se face prin procedeul calorimetriei, utilizandu-se aparate numite bombe calorimetrice. Cea mai cunoscuta bomba calorimetrica este cea a lui Berthelot, care consta intr-un container de otel captusit cu platina, umplut cu oxigen prin intermediul unui tub. In interiorul containerului se gaseste un filament electric, cu ajutorul caruia este arsa o cantitate cunoscuta dintr-un aliment sau dintr-o substanta organica oarecare. Caldura dezvoltata este absorbita de volumul de apa in care este scufundat containerul. Cunoscand volumul apei si numarul de grade cu care aceasta si-a modificat temperatura, se poate calcula cantitatea de caldura rezultata din arderea substantei respective.
Pe cand in bomba calorimetrica sunt arse complet toate cele trei categorii de substante nutritive, in organism numai glucidele si lipidele sunt degradate complet, pana la CO2 si H2O, in timp ce proteinele sufera o degradare incompleta (pana la uree, amoniac, creatinina si acid uric), rezultand o caldura de combustie mai mica decat in bomba calorimetrica.
Valorile caldurii de combustie in organism a substantelor nutritive sunt: glucide - 4,1 kcal/g, lipide - 9,3 kcal/g, proteine - 4,2 kcal/g. Prin arderea proteinelor in bomba calorimetrica rezulta, in medie, 5,66 kcal/g. Diferenta dintre aceasta valoare si 4,2 kcal/g reprezinta valoarea calorigena a produsilor finali ai metabolismului proteic, in special a ureei si amoniacului.
Daca se ia in considerare nu cantitatea de alimente ingerate, ci cantitatea de alimente care este absorbita si deci efectiv furnizata organismului (digestibilitatea medie a fiecarei categorii de alimente), valorile medii ale caldurii de combustie sunt mai mici si anume: glucide - 3,88 kcal/g, lipide - 8,65 kcal/g, proteine - 3,68 kcal/g.
3. Metoda bazata pe analiza excretei - se determina cantitatile de azot si de carbon eliminate si se admite ca fiecare gram de azot corespunde la 6,25 g de proteine consumate si ca fiecare gram de carbon suplimentar (defalcare facuta din carbonul protidic) corespunde la 1,307 g de lipide consumate. Aceasta metoda prezinta doua inconveniente:
nu ia in considerare degradarea glucidelor in organism
nu poate fi utilizata decat in cazul experientelor de durata, deoarece eliminarea deseurilor se produce cu o oarecare intarziere fata de degradarea alimentelor in organism.
Cheltuiala de energie a unui organism animal poate fi masurata prin doua metode: metoda calorimetriei directe si metoda calorimetriei indirecte.
1. Metoda calorimetriei directe reclama utilizarea unor aparate numite calorimetre (pot avea constructii foarte variate), care, in esenta, constau dintr-o camera speciala, ermetic inchisa, in care se introduce animalul de experienta, si din diferite dispozitive care permit transmiterea caldurii degajata de acesta unui mediu material (gheata, aer si mai ales apa). Primii cercetatori care au masurat caldura animala au fost Crawford (1770), Lavoisier si Laplace (1780).
Dintre numeroasele tipuri de calorimetre vom descrie calorimetrul Atwater - Benedict, destinat cercetarilor pe om. Acesta consta dintr-o incinta de marimea unei camere mici, cu pereti dubli si capitonati cu pluta (asigura izolarea termica). In interiorul camerei se gaseste un radiator prin care circula apa rece, a carei temperatura creste datorita caldurii degajate de subiect. Cunoscandu-se masa de apa care a circulat prin radiator(m), temperatura apei la intrare (t) si la iesire (t'), se poate calcula caldura degajata de subiect (Q) dupa formula: Qkcal = mkg(t'-t).
Oxigenul consumat de subiect este inlocuit cu ajutorul unui dispozitiv anexa, iar CO2 si vaporii de apa eliminati sunt retinuti intr-un flacon cu KOH (sau calce sodata) si, respectiv, intr-unul cu acid sulfuric. Prin cantarirea acestuia din urma la sfarsitul experientei, se determina cantitatea de apa eliminata de subiect prin plamani si prin piele. Aceasta cantitate (exprimata in grame) se inmulteste cu 0,586 cal (cantitatea de caldura necesara pentru evaporarea unui gram de apa la 37sC), obtinandu-se astfel caldura pierduta sub forma latenta, care se adauga la caldura pierduta prin iradiere, determinata anterior. Acest aparat permite deci si determinarea schimburilor respiratorii, motiv pentru care se mai numeste si calorimetru respirator.
Metoda calorimetriei directe, desi asigura o mare precizie a determinarilor, este numai rareori utilizata, deoarece reclama instalatii complicate si un timp indelungat de experienta.
2. Metoda calorimetriei indirecte este mai simpla si mai rapida, ceea ce explica si larga ei utilizare. Prin calorimetrie indirecta se intelege determinarea schimburilor respiratorii ale unui organism, adica a oxigenului consumat si a bioxidului de carbon eliminat de acesta intr-o unitate de timp. Pornind de la valorile ambelor gaze respiratorii sau numai de la valoarea unuia dintre ele (de regula a oxigenului consumat) se poate calcula cantitatea de caldura produsa si implicit degajata de catre organismul respectiv prin inmultirea volumului gazului respirator luat in considerare cu coeficientul termic corespunzator (cantitatea de caldura degajata in timpul utilizarii a 1 g sau 1 l de oxigen si, respectiv, al producerii aceleiasi cantitati de bioxide de carbon). Atat coeficientul termic al oxigenului cat si cel al bioxidului de carbon variaza in functie de natura substantelor nutritive (folie).
Deoarece, pe de o parte, in mod practic, organismul utilizeaza toate cele trei categorii de substante nutritive, iar pe de alta parte, coeficientii termici ai oxigenului corespunzatori acestora sunt foarte apropiati, se utilizeaza, de regula, coeficientul termic mediu al oxigenului (4,83 kcal/l).
In ceea ce priveste coeficientii termici ai CO2, acestia nu se recomanda a fi folositi deoarece mai multe cauze (printre care retinerea in organism a unei parti a CO2 prin fixare sub forma de bicarbonati) determina aparitia unor importante incongruente intre producerea de CO2 si degajarea de caldura.
Pentru a obtine indicatii despre
substantele nutritive care au fost oxidate preferential in organism,
este necesar ca, pe langa oxigenul consumat, sa determinam
si bioxidul de carbon degajat, ceea ce ne permite sa calculam
catul respirator (QR), care reprezinta raportul dintre CO2
si O2: 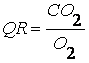
Fiecarei categorii de substante nutritive ii corespunde o anumita valoare a QR si anume: 0,8 pentru proteine, 0,7 pentru lipide si 1 pentru glucide. La om, in cazul unui regim alimentar mixt, catul respirator variaza, de regula, intre 0,85 si 0,9. Valoarea calorica a oxigenului este diferita pentru diferite valori ale catului respirator.
Daca la un organism, in afara de consumul de oxigen si producerea de bioxid de carbon, determinam si excreta de azot, putem stabili, printr-o serie de calcule, compozitia amestecului metabolic.
Pentru determinarea schimburilor respiratorii se utilizeaza diferite aparate si instalatii, adecvate dimensiunilor animalului, tipului sau de respiratie (acvatica sau aeriana) iar in cazul omului, adecvate scopului in care se face determinarea (clinic sau experimental).
III.3 Intensitatea metabolismului energetic
Intensitatea metabolismului energetic global variaza de la o specie la alta, de la un individ la altul, si la acelasi individ, in functie de starea fiziologica sau alti factori.
Speciile de animale cu mod de viata sedentar sau semisedentar au un metabolism energetic relativ scazut (folie). In cazul speciilor foarte inrudite, metabolismul energetic este mai ridicat la cele vioaie si rapide decat la cele mai putin active. De exemplu, la Arenicola (30 mm3O2/kg/h), Pecten (70 mm3O2/kg/h) si pastrav (230 mm3O2/kg/h), consumul de oxigen este mai crescut decat la speciile inrudite - Chaetopterus (8 mm3O2/kg/h), Stridie (6 mm3O2/kg/h) si, respectiv, tiparul (130 mm3O2/kg/h). Orice activitate biologica determina cresterea intensitatii metabolismului. La unele animale exista o diferenta foarte mare intre metabolismul de repaus si cel de activitate (astfel, la unele insecte - Melolontha - 360 mm3O2/kg/h in repaus si 39700 mm3O2/kg/h in timpul zborului, diferite lepidoptere, metabolismul din timpul zborului este cu peste 100 de ori mai crescut decat acela din timpul starii de repaus). La alte insecte (Coccinella, diferite diptere) metabolismul creste in timpul zborului numai de 20 de ori. Metabolismul de activitate al colibrilor este de circa 8 ori mai ridicat decat metabolismul de repaus.
Cercetarile comparative au aratat o crestere liniara a consumului de oxigen cu viteza de deplasare a animalului, aceasta crestere fiind mai rapida la animalele mici decat la cele mari. Cantitatea de energie necesara pentru transportul unei unitati de masa corporala pe o unitate de distanta scade cu cresterea greutatii corporale a animalului.
In legatura cu intensitatea metabolismului energetic au fost stabilite doua legi: legea taliilor si legea suprafetelor.
Legea taliilor arata ca exista o relatie invers proportionala intre talia animalelor (exprimata prin greutatea lor) si productia de caldura raportata la unitatea de greutate corporala. Deci, cu cat un animal este mai mic, cu atat productia lui de caldura este mai mare. De exemplu, la soarece, intensitatea metabolismului raportata la 1 Kg greutate corporala si la 24 ore este de circa 17 ori mai ridicata decat la elefant. Exista insa si exceptii de la aceasta regula. Astfel, la unele larve si stadii imago de insecte (ortoptere, diptere), precum si la unele moluste (Dreissena, Anodonta, Unio), intensitatea relativa a metabolismului (raportata, de exemplu, la 1 kg greutate corporala si 1 ora) este independenta de greutatea corporala. Cercetarile realizate de Picos si Cucerzan (1967) pe Anodonta piscinalis au dus la aceeasi concluzie: la indivizii mari din grupele mentionate, consumul de oxigen pe unitatea de greutate corporala este acelasi ca la indivizii mai mici.
Legea suprafetelor conceputa initial ca aplicandu-se numai la homeoterme, arata ca productia de caldura raportata la unitatea de suprafata corporala (m2), se exprima prin valori apropiate la cele mai diferite specii homeoterme (folie). Cercetarile ulterioare au aratat valabilitatea cestei legi si la vertebratele poikiloterme si chiar la unele nevertebrate.
Legea suprafetelor a fost criticata de numerosi autori, aducandu-i-se numeroase obiectii, dintre care mentionam:
nu poate fi comparata intensitatea pierderii de caldura a pielii animalelor cu pilozitate redusa cu cea a celor cu pilozitate abundenta (iepure)
marimea suprafetei tegumentare prin care se pierde caldura depinde de postura
Pentru determinarea suprafetei corporale au fost propuse diferite formule matematice, dintre care, foarte cunoscuta este cea a lui Meeh:
![]() sau
sau ![]() sau
sau ![]()
in care S este suprafata corpului (in cm2), P - greutatea (in grame) ,iar K este o constanta care depinde de forma generala a corpului (valorile lui K la diferite specii de vertebrate - folie). In ceea ce priveste exponentul 2/3, aceasta a facut obiectul unor serioase controverse (diferiti cercetatori propunand alti exponenti). In 1935, cu prilejul unui congres in legatura cu metabolismul energetic, s-s stabilit un exponent de 0,73.
Intensitatea metabolismului energetic este influentata nu numai de talia animalului, ci si de alti factori individuali (sex, varsta), precum si de o serie de factori externi, dintre care cel mai important este temperatura mediului ambiant.
Metabolismul bazal reprezinta cheltuiala minima de energie, strict necesara mentinerii vietii, adica, asa cum a numit-o Magnus Levy "cheltuiala de fond". Cea mai mare parte a energiei metabolice bazale este utilizata pentru activitatea muschilor respiratori, inimii, rinichilor si ficatului.
La un individ de 20 ani, 60 kg si 1,70 cm, valoarea metabolismului bazal este de aproximativ 1600 kcal/zi; la un individ de aceeasi greutate si inaltime, in varsta de 60 de ani, metabolismul bazal este de 1425 kcal/zi.
In timpul somnului, cheltuiala de energie scade cu circa 10% fata de cea bazala, din cauza scaderii functiilor organismului (scaderea respiratiei, circulatiei, tonusului muscular si secretiilor glandulare). In hipnoza se produce, de asemenea, o scadere a metabolismului bazal (daca, bineinteles nu s-s sugerat cresterea lui).
Termoreglarea
Termoreglarea este o functie a organismelor animale, de o complexitate corespunzatoare nivelului lor de evolutie si care asigura un anumit grad de constanta a temperaturii lor corporale (homeostazie termica) - termenul de "constanta" a mediului intern a fost introdus de Claude Bernard in 1864, iar cel de homeostazie de Cannon in 1928.
1. Temperatura corporala a animalelor
Din punct de vedere al dependentei temperaturii corporale (interne) de temperatura mediului ambiant, animalele se impart in doua categorii: poikiloterme si homeoterme.
Animalele poikiloterme (ectoterme) au o temperatura corporala care, in limite si grade diferite la diferite specii, variaza in functie de temperatura mediului extern. Din aceasta categorie fac parte toate nevertebratele si vertebratele inferioare (ciclostomi, pesti, amfibieni si reptile). Adesea li se atribuie si denumirea de animale "cu sange rece", insa, aceasta denumire este improprie, ea nefiind valabila in toate cazurile (de exemplu, pestele Leuciscus ceylonensis, care traieste in ape termale, la peste 50sC, are temperatura corporala mult superioara aceleia a homeotermelor - denumite si animale "cu sange cald", ea fiind insa variabila).
Dependenta temperaturii corporale a poikilotermelor de temperatura mediului extern se explica prin urmatoarele:
intensitatea redusa a proceselor lor metabolice
imperfectiunea mecanismelor lor de termoreglare
Aceasta dependenta nu este insa totala, deoarece temperatura corporala a animalelor poikiloterme nu urmeaza in mod fidel variatiile temperaturii mediului si, in general, nu este identica cu aceasta, ci mai crescuta cu de la cateva zecimi de grad la cateva grade (dupa Chanton si Paniel, 1966, excesul temperaturii interne fata de cea a mediului ambiant este de: 0,2-0,3sC la pesti, 0,5-0,9sC la broaste testoase, 1-2sC la broasca, 5,5sC la vipera si 7sC la soparla). Acest decalaj termic se datoreaza interventiei sistemului nervos, iar, in unele cazuri, prezentei depozitelor de grasime.
Animalele homeoterme (endoterme) au temperatura corporala relativ constanta, mult superioara mediei temperaturii ambiante si putin dependenta de ea. In aceasta categorie se includ pasarile si mamiferele, care dispun de mecanisme perfectionate de termoreglare (capabile sa asigure constanta temperaturii lor corporale). Faptul ca temperatura lor corporala este superioara temperaturii mediului se explica prin nivelul ridicat al termogenezei.
La pasari temperatura corporala este mai ridicata decat cea a mamiferelor (vrabie - 44,5sC, gaina - 42,5sC, rata - 42sC, porumbel - 42sC; la mamifere: lup - 40,5sC, bovine, porcine - 39,5sC,la maimuta - 38,5sC, la om - 37sC, la tatu - 34sC, la ornitorinc - 32sC ).
Mamiferele hibernante sunt denumite de unii autori heteroterme, deoarece, in timpul repausului hibernal, temperatura lor corporala scade si urmeaza fluctuatiile temperaturii ambiante.
Constanta temperaturii corporale la homeoterme nu este generala si nici absoluta, asa cum demonstreaza urmatoarele exemple:
la camila, in timpul zilei, temperatura interna se modifica cu 6sC si anume creste de la 34-35sC dimineata, dupa o noapte racoroasa, la 41sC, cand temperatura externa atinge valoarea maxima
nou-nascutii unor specii de homeoterme trec printr-o perioada de poikilotermie, a carei durata depinde de mai multi factori (in special modul de viata al speciilor respective). Astfel, in timp ce puii pasarilor nidifuge devin homeotermi aproape imediat dupa eclozare, aceia ai pasarilor nidicole sunt poikilotermi o perioada indelungata, in care au nevoie de caldura corpului matern. In ceea ce priveste mamiferele, puii de cobai, manjii si viteii sunt homeotermi chiar de la nastere, pe cand puii altor specii (iepure, caine, maimuta, om), in primele 8-15 zile ale vietii lor sunt poikilotermi.
chiar si la homeotermele adulte temperatura corporala sufera variatii, uneori destul de importante, sub influenta mai multor factori: varsta, sex, activitate musculara, digestie, temperatura aerului, perioada zilei si altii.
Un interes deosebit prezinta variatiile suferite de temperatura corporala a homeotermelor in decursul perioadei de 24 ore (variatiile nictemerale sau circadiene). La speciile diurne temperatura cea mai ridicata se inregistreaza in timpul zilei si cea mai scazuta in timpul noptii, iar la speciile nocturne, situatia este inversa. La pasari, diferenta dintre temperatura corporala in perioada activa si cea din perioada inactiva este de 1-4sC.
La om, cea mai mica temperatura corporala (36,5sC) se inregistreaza spre orele 4 dimineata, iar cea mai mare (37,7sC), spre orele 16 dupa amiaza. Aceasta hipertermie vesperala ser datoreaza, in parte, activitatilor diurne (musculara, digestiva si intelectuala). In timp ce la om variatiile circadiene ale temperaturii interne nu depasesc 1sC, la mamiferele mari aceste variatii sunt mai importante (de exemplu, la hipopotam, temperatura interna este de circa 34sC dimineata si de circa 39sC dupa amiaza).
Un alt factor care modifica temperatura umana este stressul psihic, asa cum au aratat si cercetarile lui Gotev si colaboratorii (1950, 1971), realizate pe un esantion mare de studenti la medicina, farmacie si biologie in zilele de examen la unele discipline grele, cand, la 92% din studentii cercetati, temperatura corporala a crescut pana la 38,4sC.
2. Homeostazia termica la animalele poikiloterme
Termoreglarea nu este apanajul exclusiv al homeotermelor, ea intalnindu-se, intr-un grad mai redus, si la poikiloterme. Existenta unor mecanisme simple de termoreglare la poikiloterme a fost admisa de Dajos (1971), care a observat ca, la asa numitele cicloterme, datorita unor astfel de mecanisme, temperatura corporala inregistreaza o usoara scadere cand temperatura externa depaseste 30sC si o usoara crestere cand ea scade sub 10sC.
Faptul ca animalele poikiloterme dispun de unele posibilitati de termoreglare a fost demonstrat experimental inca din 1898 de catre Vernon care, cercetand dependenta de temperatura a producerii de dioxid de carbon la diferite specii (rama, melc, broasca etc.) a constatat existenta unor intervale termice (de marime diferita la diferite specii), in limitele carora, desi temperatura creste, producerea de CO2 ramane relativ constanta.
Acelasi autor a demonstrat, de asemenea, ca aceasta relativa homeostazie termica se datoreaza interventiei sistemului nervos. Astfel, la broaste, el a constatat ca, dupa sectionarea transversala a axului cerebro-spinal, la marginea inferioara a bulbului, se produce diminuarea degajarii de CO2 si disparitia intervalelor termice in care acest indice fiziologic ramane constant. La broastele la care sectiunea era facuta la nivelul marginii superioare a bulbului, intervalele termice mentionate s-au pastrat, iar controlul nervos al degajarii de CO2 nu a fost abolit. Din rezultatele acestor experiente, Vernon a dedus ca, la broaste, metabolismul tisular este controlat de un centru nervos situat in bulb.
Cercetarile lui Remorov (1963) au aratat ca, la broasca (Rana ridibunda), rolul principal in termoreglare este indeplinit de creierul mijlociu, deoarece, dupa indepartarea acestuia, temperatura corpului urmeaza exact temperatura mediului.
Existenta unui interval termic de metabolism constant a fost, de asemenea, demonstrata si la pesti de catre Stroganov (1939), care l-a denumit zona termica de adaptare, iar mai tarziu, de catre Schmeing-Engberding (1953), care l-a denumit interval constant. Ultimul autor a gasit ca marimea acestui interval difera de la o specie la alta, fiind cuprinsa, de exemplu, intre 20 si 24sC la puietii de linsi intre 21 si 25sC la puietii de crap. Acelasi autor a constatat ca, daca pesti erau narcotizati, intervalele constante dispareau, ceea ce constituie o dovada indirecta in sprijinul faptului ca sistemul nervos este responsabil de existenta acestor intervale.
S-a stabilit marimea intervalului termic de metabolism constant si la alte grupe de animale poikiloterme: la serpi (Natrix natrix), acest interval este cuprins intre 9 si 19sC (Schmidt, 1971), la melci (Helix pomatia) intre 15 si 20sC (Grossu-Moisa, 1971) la miriapode (Strongylosoma pallipes), intre 22 si 28sC (Gromysz-Kalkowska, 1973).
3. Homeostazia termica la homeoterme
Dispunand de mecanisme perfectionate de autoreglare, homeotermele sunt capabile sa-si mentina temperatura corporala la un nivel constant, intr-un domeniu foarte larg de variatii ale temperaturii mediului extern (intre + 50sC si - 50sC).
Dupa efectele diametral opuse pe care le produc, se deosebesc doua mecanisme de autoreglare: mecanisme chimice (metabolice), prin care se realizeaza producerea de caldura (termogeneza) si mecanisme fizice, prin care se realizeaza pierderea de caldura (termoliza).Atunci cand, datorita unor cauze externe sau interne, organismul tinde sa se raceasca, intra in actiune mai ales mecanismele chimice, iar atunci cand el tinde sa se supraincalzeasca, intra in actiune mai ales mecanismele fizice. Cu alte cuvinte, desi in masura inegala, atat intr-un caz cat si in celalalt, participa ambele categorii de mecanisme.
Intr-adevar, organismul lupta impotriva racirii nu numai prin intensificarea producerii de caldura, ci si prin reducerea pierderii caldurii. Pe de alta parte, in lupta impotriva supraincalzirii, intensificarea pierderii de caldura este insotita de o usoara crestere a metabolismului, reclamata de activitatea crescuta a organelor circulatorii, respiratorii si glandelor sudoripare. In legatura cu aceasta Frdricq (1944) afirma: "lupta pentru homeostazie reclama o cheltuiala energetica reala, si aceasta oricare ar fi sensul - scadere sau crestere - in care temperatura exterioara se indeparteaza de neutralitatea termica: mecanismele termoregulatoare nu functioneaza in mod gratuit".
Neutralitatea termica este o stare de confort termic al organismului homeoterm in repaus, determinata de un anumit nivel al temperaturii mediului, in conditiile caruia termogeneza si termoliza sunt exact echilibrate (deci mecanismele termoregulatoare nu intervin), iar valoarea metabolismului este minima. Dupa cum afirma Lefevre "un mediu asigura neutralitatea termica atunci cand el nu provoaca nici racirea, nici incalzirea corpului si permite scurgerea caldurii sale pe masura ce ea se produce".
Neutralitatea termica corespunde unui domeniu de temperaturi ambiante variabil in functie de specie, denumit zona de neutralitate termica. La omul dezbracat, aceasta este cuprinsa intre 28 si 30sC. In ceea ce priveste animalele, unele au o zona de neutralitate termica mai ingusta (de exemplu 28-30sC la sobolan, soarece, cobai, porumbel, 25-29sC la caine), iar altele o zona mai larga (0-28sC la oaia netunsa, 15-28sC la cocos, gaina , 10-21sC la iepurele de casa - Ryback, 1962).
3.1. Producerea de caldura (termogeneza)
Avand un metabolism ridicat, organismul homeoterm este capabil sa produca o cantitate mare de caldura (lucru demonstrat de faptul ca temperatura lui este superioara celei a mediului ambiant).
Intrucat cantitatea de caldura produsa de un organism este egala cu aceea pe care el o pierde in exterior, prin masurarea acesteia din urma (mai ales prin metoda calorimetriei indirecte) s-a putut stabili valoarea celei dintai, la cele mai diferite specii de homeoterme:
Cantitatea de caldura produsa de unele homeoterme
(dupa Chanton si Paniel, 1966)
|
Productia de caldura |
|||
|
pe kg/24 ore (kcal) |
pe m2/ora (kcal) |
||
|
Soarece | |||
|
Sobolan | |||
|
Porumbel | |||
|
Cobai | |||
|
Iepure | |||
|
Caine | |||
|
om | |||
Analiza datelor din tabel permite sa se constate, pe de o parte, ca productia de caldura pe unitatea de greutate corporala este cu atat mai mare cu cat animalul este mai mic (in conformitate cu legea taliilor), iar, pe de alta parte, ca productia de caldura pe unitatea de suprafata corporala are valori apropiate la cele mai diferite specii de homeoterme (in conformitate cu legea suprafetelor).
Organele care participa cel mai mult la termogeneza totala sunt muschii si ficatul, care au si cel mai intens metabolism. Scheletul, tesutul conjunctiv si pielea, care au un metabolism scazut, participa la termogeneza totala numai intr-o masura neinsemnata.
Atunci cand homeostazia termica este periclitata din cauza frigului, se produce cresterea termogenezei, mai ales prin intensificarea metabolismului in muschii scheletici si in ficat. In functie de intensitatea frigului si de specia animalului, cresterea termogenezei se realizeaza, fie prin tremuraturi musculare (termogeneza cu frison), fie prin intensificarea oxidarilor celulare, mai ales in ficat (termogeneza fara frison).
Tremuraturile sunt contractii musculare rapide, care nu se soldeaza cu efectuarea de lucru mecanic, ci numai cu producerea de caldura, care este integral utilizata pentru incalzirea corpului. Ele se declanseaza in mod reflex, prin excitarea termoreceptorilor tegumentari.
La pasari, Steen si Enger (1957) au constatat ca, intre +20 si - 24sC, intensitatea tremuratului creste paralele cu scaderea temperaturii. La pasarile adulte, termoreglarea chimica se realizeaza numai prin termogeneza cu frison, in timp ce la pui este prezenta si termogeneza fara frison.
In ceea ce priveste mamiferele, cele mai multe dintre ele, atunci cand sunt expuse la frig, isi intensifica termogeneza prin frison muscular. La om aceasta poate sa mareasca productia de caldura de 3-4 ori, iar temperatura corporala, cu mai mult de 0,5sC.
Atat la pasari cat si la mamifere, cresterea termogenezei se realizeaza si prin ingerarea hranei, care este urmata de asa-numita actiune dinamica specifica (ADS), adica de producerea de caldura suplimentara. Alimentele de natura proteica au cea mai mare ADS, aceasta reprezentand 25-30% din valoarea energetica a proteinelor ingerate. S-a stabilit ca, dupa ingerarea de alimente, mai ales cand aceste contin proteine, se reduce necesitatea producerii de caldura prin tremurat.
3.2. Pierderea de caldura (termoliza)
Intrucat, de regula, temperatura mediului este mai scazuta decat cea a organismului homeoterm, acesta pierde continuu caldura in mediu, cu atat mai mult cu cat decalajul termic este mai mare. In cea mai mare parte, pierderea de caldura se face prin piele si prin suprafetele respiratorii, datorita urmatoarelor fenomene fizice: conductia, convectia, iradierea si evaporarea apei.
Pierderea de caldura prin primele trei fenomene este mai importanta la nivelul pielii si depinde de marimea suprafetei corpului, diferenta de temperatura intre piele si mediul ambiant, natura si culoarea suprafetei corpului.
Pierderea caldurii prin conductie se realizeaza prin contactul fizic al animalului cu substratul, cu diferite obiecte si cu aerul linistit.
Pierderea caldurii prin convectie are loc atunci cand aerul este in miscare, fapt care permite ca aerul incalzit la contactul cu pielea, sa fie reinnoit cu aer mai rece.
Izolarea termica, realizata prin penaj si blana la animale si prin imbracaminte la om, reduce la minim pierderea de caldura prin conductie si convectie, care sunt totdeauna asociate.
Pierderea de caldura prin iradiere se realizeaza prin intermediul undelor electromagnetice infrarosii, care, in cazul pielii mamiferelor, au lungimea de unda de 5-20 μm. Puterea de emisie a radiatiilor calorice depinde de natura suprafetei corporale. Intr-adevar, la pasarile cu penaj bogat sau la mamiferele cu blana, pierderea de caldura este mai mica decat la acelea cu invelisuri corporale reduse. De asemenea, pierderea de caldura prin iradiere este influentata de culoarea suprafetei corporale. O suprafata corporala de culoare inchisa emite o cantitate mai mare de radiatii calorice decat o suprafata corporala de culoare deschisa. Asa se explica faptul ca blana animalelor este colorata in regiunile calde si alba in regiunile reci.
Pierderea caldurii prin evaporarea apei
La temperaturi ambiante foarte ridicate, evaporarea apei la suprafata pielii, sub forma de sudoare, are rolul principal in lupta organismului impotriva supraincalzirii. La aceasta modalitate de pierdere a caldurii recurge indeosebi organismul uman, deoarece la pasari si la majoritatea mamiferelor, glandele sudoripare lipsesc sau sunt putin numeroase.
Prin evaporarea sudorii la suprafata pielii se pierde foarte multa caldura, si anume 586 kcal pentru un litru de apa la 37sC. Transpiratia este puternica chiar la 30sC si se intensifica paralel cu cresterea temperaturii. Atunci cand se efectueaza o munca grea, ea se produce si in mediu rece.
Daca umiditatea aerului este prea crescuta, sudoarea nu se mai poate evapora si deci nu se mai produce nici o pierdere de caldura prin intermediul ei. De asemenea, cand aerul este uscat, sudoarea care se scurge in picaturi (de exemplu, pe frunte) nu participa la termoreglare.
Atunci cand, din cauza transpiratiilor abundente, se produc pierderi foarte mari de apa si saruri, se poate produce moartea organismului.
In ceea ce priveste pierderea de caldura prin suprafetele respiratorii, aceasta este mai putin importanta la om (8% din totalul caldurii pierdute), dar foarte importanta la pasari si la majoritatea mamiferelor, la care glandele sudoripare sunt absente sau putin numeroase. La aceste animale, evaporarea respiratorie este intensificata prin accelerarea miscarilor respiratorii (polipnee termica).
La pasari, polipneea este o reactie obisnuita la cresterea temperaturii mediului, frecventa miscarilor cutiei toracice fiind diferita la diferite specii (de la 40 la 700/minut). La numeroase pasari (gaina, rata, porumbel), polipneea termica este asociata cu pulsatii rapide ale regiunii gatului (Flutterul gatului), care reclama o cheltuiala de energie mult mai mica decat miscarile cutiei toracice.
In ceea ce priveste mamiferele, polipneea termica este bine cunoscuta la caini, caracterizandu-se prin miscari respiratorii rapide (250-260/minut), de amplitudine mica si in timpul carora gura este deschisa, iar limba atarna in afara.
La om, la temperatura normala a camerei, pierderile de caldura pe diferite cai se exprima prin urmatoarele valori procentuale aproximative:
Iradiere, convectie si conductie........65%
Evaporarea apei prin piele si plamani.....30%
Incalzirea aerului inspirat..........3%
Pierderi de caldura prin urina si fecale.....2%
In lupta impotriva supraincalzirii corpului, homeotermele recurg si la o serie de acte comportamentale, prin care evita caldura excesiva a mediului sau faciliteaza pierderea de caldura prin mecanismele fizice prezentate anterior. Un comportament frecvent intalnit este retragerea in adaposturi mai racoroase. Prin comportamente adecvate, animalele evita nu numai supraincalzirea ci si racirea corpului. Totalitatea acestor comportamente constituie asa-numita termoreglare comportamentala, care are o importanta primordiala pentru animalele poikiloterme.
4. Controlul neuro-endocrin al termoreglarii
4.1. Rolul sistemului nervos in termoreglare
Asa cum am aratata anterior, chiar si la animalele poikiloterme, sistemul nervos este responsabil de homeostazia termica limitata intalnita la acestea. La homeoterme, pentru care homeostazia termica este o caracteristica esentiala, rolul sistemului nervos in termoreglare a facut obiectul multor cercetari, in special in cazul mamiferelor.
Controlul nervos al termoreglarii se realizeaza cu participarea termoreceptorilor, cailor aferente, centrilor nervosi si cailor eferente.
Termoreceptorii sunt structuri specializate pentru receptionarea variatiilor de temperatura; dupa topografie, se impart in trei categorii: periferici (cutanati), interni (viscerali) si centrali.
a) termoreceptorii cutanati sunt de doua feluri: pentru cald (corpusculii Ruffini) si pentru rece (corpusculii Krause). Cercetarile din ultimul timp au aratat insa ca acesti receptori nu sunt riguros specializati. Receptorii pentru rece sunt situati mai superficial si sunt mai numerosi, iar receptorii pentru cald sunt situati mai in profunzimea dermei si sunt in numar mai mic. La mamifere, activitatea maxima a receptorilor pentru rece se manifesta la temperatura cutanata de 25-30sC, iar a celor pentru cald, la circa 40sC. Termoreceptorii cutanati au un prag redus de excitabilitate. Intr-adevar s-a constatat ca, daca pe piele sau pe limba se produc variatii de temperatura de 1/10sC sau chiar mai mici, acestea sunt capabile sa provoace modificarea senzatiei de rece sau cald.
b) termoreceptorii interni (viscerali) sunt situati in mucoasele organelor interne, existenta lor fiind multa vreme contestata. Prin metoda reflexelor conditionate s-a demonstrat prezenta acestora in mucoasa gastrica.
c) termoreceptorii centrali - aceasta denumire se aplica neuronilor termosensibili, existenti in diferite segmente ale SNC si care sunt excitati de modificarile temperaturii sangelui. In afara de diencefal, unde au fost constatati mai demult, termoreceptorii centrali se gasesc si in alte segmente nervoase (bulb, maduva spinarii) asa cum au aratat numeroase cercetari. Astfel, cercetarile lui Chai si Lin (1972) pe maimute (Macaca cyclopis) au demonstrat ca atat in maduva spinarii cat si in bulb, se gasesc elemente termosensibile, care participa la termoreglare (prin incalzirea maduvei sau a bulbului de la 38 la 42-43sC a produs vasodilatatie cutanata, polipnee, bradicardie si o usoara scadere a temperaturii corporale; dimpotriva, racirea maduvei sau a bulbului de la 38 la 32-33sC a produs vasoconstrictie, incetinirea respiratiei, tahicardie, hipertensiune si o usoara crestere a temperaturii corporale).In timp ce racirea maduvei a provocat tremuratul celor patru membre, racirea bulbului a provocat numai tremuratul falcilor.
Atat la pasari cat si la mamifere, responsabilitatea principala a controlului mecanismelor de termoreglare revine diencefalului.
La pasari se admite, desi cu unele rezerve, ca centrul termoreglarii se gaseste in regiunea talamica. In acest sens, pledeaza cercetarile lui Rogers (1928) care au aratat ca, daca talamusul porumbeilor decerebrati este excitat direct cu aer rece sau cu apa rece, precum si cu curent electric, se produc tremuraturi musculare si cresterea temperaturii corporale. Pe de alta parte, Frdricq (1944) relateaza faptul ca, dupa distrugerea talamusului, porumbelul devine poikiloterm.
In ceea ce priveste mamiferele, este bine demonstrat faptul ca, la acestea, mecanismele reglarii sunt controlate de centri hipotalamici. In hipotalamusul anterior exista un centru de racire sau de pierdere a caldurii, iar in hipotalamusul posterior, un centru de incalzire sau de producere si pastrare a caldurii. Experientele au aratat ca, dupa lezarea hipotalamusului anterior, animalul devine incapabil de a-si regla temperatura intr-un mediu cald si adesea moare, din cauza hipertermiei, chiar daca este tinut la temperatura camerei. Lezarea hipotalamusului posterior determina diminuarea capacitatii animalului de a-si mentine temperatura normala, atat intr-un mediu cald, cat si intr-un mediu rece, ceea ce inseamna ca el devine poikiloterm.
La animalul intact, centrii hipotalamici ai termoreglarii sunt excitati de modificarile temperaturii sangelui, provocate de modificarile temperaturii mediului. Cresterea acesteia din urma excita centrul de pierdere a caldurii, care declanseaza reactiile de lupta contra supraincalzirii corpului (vasodilatatie periferica, transpiratie etc.). Scaderea temperaturii mediului excita centrul de producere a caldurii, care declanseaza reactiile de aparare impotriva racirii corpului: tremuraturi si hipersecretie de adrenalina (care maresc termogeneza), vasoconstrictie periferica si piloerectie (care maresc izolarea superficiala, contribuind astfel la pastrarea temperaturii corpului). La om piloerectia nu participa la termoreglare, dar se pastreaza sub forma unui vestigiu cunoscut sub numele de "piele de gaina".
Activitatea centrilor hipotalamici ai termoreglarii este influentata si de impulsurile care vin de la termoreceptorii periferici. De la receptorii pentru rece pleaca impulsuri care excita centrul de producere si pastrare a caldurii si inhiba centrul de pierdere a caldurii. Efecte diametral opuse produc impulsurile de la receptorii pentru cald.
Centrii hipotalamici ai termoreglarii pot fi activati in trei moduri: direct, reflex si prin asociatie intercentrala.
Activarea directa - este produsa in mod obisnuit de modificarile temperaturii sangelui (acest fapt a fost demonstrat experimental), dar se poate realiza si de numeroase substante biologic active, existente in sange: adrenalina, noradrenalina, 5-hidroxitriptamina (serotonina), acetilcolina, histamina. S-a stabilit ca, la una si aceeasi substanta, diferite specii pot reactiona in mod diferit: astfel, in timp ce la pisica, caine si maimuta, adrenalina si noradrenalina provoaca hipotermie, iar serotonina hipertermie, la iepurele de casa si la oaie efectul este invers.
Activarea reflexa - este realizata de impulsurile care isi au originea mai ales in termoreceptorii periferici, care ajung la centrii nervosi pe caile sensibilitatii termice (fasciculele spinotalamice laterale, care conduc si excitatiile dureroase).
Activarea prin asociatie intercentrala - este demonstrata de faptul, constatat inca de la sfarsitul secolului XIX, ca inteparea diferitelor segmente encefalice (punte, mezencefal, talamus, corpi striati) determina o puternica hipertermie, motiv pentru care procedeul mentionat a fost denumit injectie termica. Alte dovezi ale existentei acestei modalitati de activare:
la om, diferite afectiuni cerebrale (tumori, encefalite, ramolismente etc.) produc tulburari de termoreglare
emotiile determina modificari ale temperaturii corporale.
Centrii hipotalamici ai termoreglarii transmit comenzile corespunzatoare la efectori, prin intermediul nervilor (somatici si vegetativi) si al hormonilor.
Inervatia somatica este reprezentata de nervii motori ai muschilor scheletici, a caror activitate mareste termogeneza. Daca temperatura mediului extern scade sub o anumita limita, creste tonusul muscular si, prin aceasta, se declanseaza frisonul termic. Acesta cuprinde mai intai muschii masticatori (fapt pentru care individul caruia ii este frig "clantane din dinti"). Persistenta frigului determina intensificarea si generalizarea frisonului (tremuratul intregii musculaturi scheletice).
Inervatia vegetativa cuprinde totalitatea fibrelor nervoase motoare, simpatice si parasimpatice, prin intermediul carora sunt declansate diferite reactii, care mentin temperatura corporala la un nivel constant. De exemplu, impulsurile transmise prin fibrele simpatice produc vasoconstrictie, iar cele transmise prin fibrele parasimpatice produc vasodilatatie. Homeostazia termica se realizeaza mai ales cu participarea sistemului nervos simpatic, deoarece majoritatea organelor implicate in termoreglare (glandele sudoripare, muschii pilosi, glanda tiroida etc.) poseda o inervatie simpatica.
4.2. Rolul hormonilor in termoreglare
Hormonii intervin mai ales in termogeneza, determinand cresterea acesteia prin intensificarea metabolismului tesuturilor.
Dintre hormonii implicati in termoreglare mentionam mai intai hormonii medulosuprarenali: adrenalina si noradrenalina (catecolamine). Noradrenalina este considerata responsabila pentru termogeneza fara frison. Cercetarile facute pe mamifere expuse la temperaturi scazute au aratat ca acestea prezentau o crestere a cantitatii de noradrenalina in sange si urina. Experimente realizate pe sobolani, la care frisonul muscular a fost blocat prin curara, au aratat ca administrarea noradrenalinei provoaca cresterea consumului de oxigen a musculaturii membrelor si al tesutului gras si totodata stimuleaza eliberarea in acesta in sange a acizilor grasi neesterificati (folositi, probabil, drept combustibil de catre tesutul muscular). Adrenalina, ca si noradrenalina, provoaca o crestere pronuntata a metabolismului energetic (efect calorigen), adrenalina participand la termoreglare si prin efectul ei vasoconstrictor.
Hormonii tiroidieni (tiroxina si trioidtironina) determina, de asemenea, o pronuntata crestere a termogenezei. Efectul hipermetabolic al tiroxinei nu se manifesta imediat, ci dupa aproximativ doua zile, creste treptata pana la un maxim, dupa care scade treptat. Trioidtironina determina cresterea metabolismului dupa un timp mai scurt (4-6 ore). In general, se considera ca tiroxina mareste termogeneza atat pe cale directa, intensificand procesele de oxido-reducere tisulare, cat si pe cale indirecta, sensibilizand celulele la actiunea adrenalinei.
Hormonii corticosuprarenali participa, de asemenea, la termoreglare, asa cum au demonstrat cercetarile realizate de diferiti autori. Astfel , cercetarile lui Maickel si colab. (1967) au aratat ca, daca cu patru ore inainte de expunerea sobolanilor suprarenalectomizati la temperatura de 4sC, acestia erau tratati cu o singura doza de cortizon, ei ramaneau capabili sa mobilizeze glucoza si acizii grasi liberi si sa-si mentina temperatura corporala normala. La sobolanii aclimatizati la rece (5sC), Bertin (1971) a constata ca, paralel cu hipersecretia de catecolamine, s-a produs si cresterea durabila a cantitatilor de corticosteron in suprarenale si plasma. Buolouard (1973) a gasit o crestere foarte mare a cantitatii de corticosteroizi la sobolani supusi 48 ore la post si frig (el presupune ca acesti hormoni intervin in gluconeogeneza).
In stadiul actual al cunostintelor noastre este dificil sa se precizeze exact rolul fiecarei glande endocrine in termoreglare, din cauza interactiunii complexe dintre ele. Pe de alta parte, rolul aceleiasi glande poate fi diferit nu numai la diferite specii, ci si la aceeasi specie, in diferite stadii ontogenetice. Activitatea glandelor endocrine care intervin in termoreglare este dirijata de hipotalamus, atat prin semnale nervoase , cat si prin semnale chimice.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3050
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved