| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Protozoarele au fost descrise in l674 de
Limitele celor trei grupe de microorganisme eucariote nu sunt clare. Exista organisme cu proprietati comune algelor si protozoarelor sau fungilor si protozoarelor. De exemplu, liniile mutante de Chlamydomonas si Volvox, faraclorofilasunt clasificate printre protozoare, iar cele cu clorofilasunt considerate caapartin algelor. Euglenofitele sunt un grup de microorganisme unicelulare, flagelate, care de cele mai multe ori, au clorofilasi sunt fotosintetizante. De aceea sunt clasificate printre alge. Dar speciile de euglenofite pot sacreascape medii complexe, la intuneric si de aceea unii autori aseazaintregul grup printre protozoare.
Mixomicetele celulare si plasmodiale sunt considerate ca fungi sau ca protozoare.
Majoritatea protozoarelor sunt organisme care traiesc liber si sunt mobile. Unele sunt microscopice (de cativa m), iar altele se vad chiar cu ochiul liber (1 mm). Unele sunt parazite pe gazde foarte diverse: alge, nevertebrate, vertebrate si chiar la om.
Se cunosc peste 45 000 de specii de protozoare. Toate necesitamedii umede, indiferent de habitat: in apa dulce si marina, in sol sau parazite la alte organisme.
Protozoarele se dezvoltain mediile care contin substante organice, pe seama carora se dezvoltabacteriile. La randul lor, bacteriile constituie o sursafoarte importantade hranaa protozoarelor.
Apele bogate in vegetatie aquatica (ape putin adanci, lacuri artificiale, canale, mlastini), sunt favorabile dezvoltarii protozoarelor. In mediul marin, macroalgele si vegetatia mlastinilor sarate, sunt habitate favorabile protozoarelor. Ele sunt abundente in instalatiile de tratare a apelor menajere, in special in namolul activ.
Multe protozoare traiesc comensale in tractul digestiv al animalelor.
Unele protozoare sunt parte componentaa zooplanctonului si se hranesc cu algele fitoplanctonului: altele tot acvatice, sunt fixate pe diferite suporturi. Putine protozoare populeazahabitate foarte specializate: intestinul termitelor, compartimentul ruminal. Ele au raporturi mutual-benefice cu gazdele.
Protozoarele care au capacitatea de descompunatori ai materiei organice, contribuie la fertilizarea solului. Cele pradatoare au un rol deosebit de important in controlul numeric al diferitelor populatii de microorganisme, deoarece se hranesc pe seama acestora.
Formele parazite produc boli la om si animale: de exemplu, boala somnului, dizenteria amoebiana, malaria, toxoplasmoza etc.
Organizare celularasi fiziologie
Complexitatea functiilor esentiale ale celulei protozoarelor necesita specializarea diferitelor componente. Celula protozoarelor este una dintre cele mai complexe din lumea vie: ea poseda organite implicate in obtinerea si digestia substantelor nutritive, in excretie si osmoreglare, in reproducere, protectie, receptionarea stimulilor, locomotie etc.
Deplasarea. Locomotia protozoarelor se realizeazaprin trei tipuri de organite: pseudopode, flageli si cili.
Pseodopodele sunt prelungiri celulare temporare, prezente la membrii grupului Sarcodina. Ele pot fi lobopodii (rotunjite), alcatuite din ectoplasma si endoplasma, filopodii (prelungiri lungi si ascutite), alcatuite numai din ectoplasmasau rizopodii (prelungiri citoplasmatice ramificate).
Pentru deplasare, ectoplasma (pelicula superficiala a citoplasmei) mai vascoasa, se fluidizeaza. Din cauza presiunii interne, in acest punct, citoplasma formeaza un pseudopod (fig. 156). Pe masura ce endoplasma fluida inainteaza, ea se schimba in ectoplasma mai vascoasa si ancoreaza pseudopodul de suport. Intreaga celula este deplasata in directia punctului de fixare a pseudopodului. Pseudopodele au rol esential in procesul fagocitarii materialului nutritiv particulat.
Flagelii protozoarelor sunt organite de locomotie, lungi si delicate. Structura si organizarea flagelilor este aproximativ aceiasi la toate categoriile de celule eucariote: doi microtubuli centrali, inconjurati de 9 dublete tubulare periferice. Aranjamentul celor 9 x 2 + 2 tubuli centrali, este caracteristic pentru majoritatea cililor si flagelilor celulelor eucariote, de la Euglena pana la om.
Protozoarele nu au perete celular. Invelisul extern sau teaca unui flagel este o continuare a membranei celulare. La baza fiecarui flagel, se gaseste un granul denumit kinetoplast, care contine ADN.
La cele mai multe flagelate, celula este prevazutacu 1-2 flageli. La un grup de flagelate parazite in intestinul termitelor, celula are zeci de flageli. Existaflagelate libere cu 4, 8 sau mai multi flageli.

Fig. 156. Ilustrarea schematica a mecanismului deplasarii amoebiene. Sagetile indica directia deplasarii.
Cilii sunt flageli miniaturali. Structura interna este aceiasi ca si a flagelului: doi microtubuli centrali si 9 dublete periferice, acoperiti de un invelis, care este o continuare a membranei celulare. Membrana citoplasmaticaare o consistentaspecialasi se numeste pelicula. Fiecare cil are la origine, un granul bazal.
Cilii pot sa acopere intreaga suprafata a celulei sau au o distributie limitata, in special la polul oral. La unele specii, cilii fuzioneazain manunchiuri compacte denumite cirri. Lungimea cililor este variata. Cei de la extremitatile celulei sunt mai lungi.
Pentru miscarea perfect coordonata, cilii sunt conectati printr-o retea de fibre situata sub membrana peliculara.
Nutritia. In general, protozoarele se hranesc prin ingestia materialului particulat sau macromolecular. Macromoleculele in mediul apos sunt inglobate prin pinocitoza. Majoritatea protozoarelor ingera materialul particulat prin fagocitoza. Pinocitoza este relativ nespecificasi continua, iar fagocitoza este un proces discontinuu si de asemenea, nespecific.
Protozoarele sunt, in general, organisme aerobe, heterotrofe, rareori anaerobe (de exemplu, cele ce traiesc in intestinul uman, in rumen sau in intestinul termitelor).
La protozoare se intalnesc doua modalitati majore de nutritie:
unele sunt autotrofe, adica sintetizeazacompusi organici din substante anorganice, pe cale fotosintetica. Sunt organisme producatoare de materie organica: glucide, proteine, lipide. Pot fi cultivate in solutii de saruri anorganice, in conditii de iluminare;
majoritatea protozoarelor sunt heterotrofe, adica organisme consumatoare si utilizeaza substantele organice preformate.
Unele protozoare heterotrofe sunt saprobionte, adicase hranesc prin absorbtia substantelor nutritive. Altele sunt holozoice, adicaingerahranaparticulatadin mediu. Ele poseda mecanisme pentru captarea si ingestia hranei. Celula inglobeazahrana particulata, intr-o vacuolade fagocitoza (fagosom), in care se elibereaza continutul enzimatic lizosomal. In timpul digestiei, dimensiunile vacuolei cresc. Produsele de digestie trec in citoplasma, iar materialele indigeste sunt excretate.
La grupul Ciliophora, hrana este ingerataprintr-o regiune diferentiata a celulei, o deschidere denumita citostom. Hrana particulatadin mediul lichid, este orientataspre citostom, prin miscarea ordonataa cililor.
Cultivare. Exigentele nutritive ale protozoarelor sunt diferite. Cele libere se cultivape medii artificiale care contin infuzie de fan sau in apa de lac, la care se adauga boabe de orez, de grau, lapte degresat. Culturile pure (axenice) se obtin mai greu, deoarece multe protozoare necesita hrana particulata.
Mediile de crestere contin hidrolizat de caseina, extract de levuri, extract de carne, glucoza, minerale. Unele specii de Amoeba si Didinium (flagelat), necesitaadaugarea in mediul de crestere, a altor microorganisme (bacterii, protozoare) ca sursa de hrana.
Unele protozoare parazite au fost cultivate in tesuturi crescute in vitro, iar altele in medii complexe, cu adaus de sange integral sau de ser. Culturile sunt deosebit de importante pentru obtinerea vaccinului si pentru testarea efectului agentilor chimioterapeutici.
Cele mai multe protozoare parazite s-au cultivat in organismul animal: sobolan, soarece, hamster. Anumite specii de Plasmodium se cultivain vivo, prin inocularea puilor de gaina.
Pentru protozoare nu existacolectii de culturi, asa cum existapentru bacterii si fungi.
Excretia. Produsele de catabolism (CO2, NH3, uree si alte substante difuzibile) sunt eliminate prin difuzie libera, la nivelul intregii membrane citoplasmatice sau prin vacuole contractile.
La ciliate, materialele indigeste sunt eliminate printr-o zonaspecializata, denumitacitopige, localizat in pozitie opusacitostomului.
Majoritatea protozoarelor din apele dulci, posedavacuole contractile. Ele actioneazaca niste pompe pentru indepartarea excesului de apacare patrunde in celula, avand astfel rol in reglarea osmolaritatii. Intr-o celula, vacuola contractilapoate fi unicasau sunt multiple. Vacuola primeste apa, fie direct din citoplasma, ori, la Paramecium, este inconjuratade mai multe canale radiare, cu rolul de a colecta apa din celula.
Ciliatele de apa dulce sau sarata, au totdeauna vacuole contractile, deoarece, chiar in mediul salin, invelisul celular este permeabil pentru apasi impermeabil pentru saruri. Frecventa pulsatiilor vacuolei, denumite sistole, depinde de cantitatea de apacare intrain celula. Speciile dulcicole, dupatransferul in apasalina, isi diminuarata pulsatiilor. Speciile mari au o frecventamai micaa pulsatiilor.
Sistemul vacuolei contractile este o membranaseparatoare intre continutul diluat (solutie hipotonica) si mediul celular cu presiune osmotica superioara.
Vacuola se deschide la nivelul unei zone specializate - un por - la suprafata celulei. La amoeba, porul este o structuratemporara, ce se formeazala inceputul fiecarei pulsatii, iar la parameci porul este o deschidere permanenta.
Structuri speciale. Unele protozoare sunt sesile, fixate pe substrat, prin intermediul unei structuri specializate denumitacrampon. Unele protozoare sesile au existentacoloniala. Celulele fiice rezultate din diviziunea celulei mama, raman asociate formand colonii bi- sau tridimensionale, cu aspect si dimensiuni caracteristice.
Majoritatea protozoarelor sunt organisme nude. Unii reprezentanti ai grupului Sarcodina formeazacochilii. Acestea sunt structuri externe rigide, secretate de celulasau formate din materiale preluate din mediul extern (granule fine de nisip, frustule de diatomee) si incorporate intr-un material mucilaginos secretat de celula. Sarcodinele din grupul Foraminifera produc, printr-un proces de secretie, cochilii de carbonat de calciu, iar Radiolaria au cochilii de siliciu sau sulfat de strontiu. Cochilia nu este totdeauna limita externaa celulei protozoarului, pentru cala multe foraminifere, protoplasma se extinde peste cochilie si chiar o acoperacomplet.
Dupa epuizarea sursei de hrana, iar experimental si sub actiunea altor factori (de exemplu, diferite noxe), multe protozoare formeazastructuri de rezistentadenumite chisti.
Reproducerea
Protozoarele se reproduc pe cale sexuata si asexuata. Unele se reproduc numai pe cale asexuata. La unele protozoare, cele douamodalitati de reproducere pot saalterneze in timpul ciclului de viata.
In procesul reproducerii asexuate, celula parentala se divide simetric sau asimetric si rezulta doua sau mai multe celule. Fisiunea binara este cel mai comun tip de reproducere asexuatasi implica diviziunea mitoticaa nucleului. In reproducerea prin fisiune binaranu se produce meioza si nici fuziunea de gameti (fig 157).
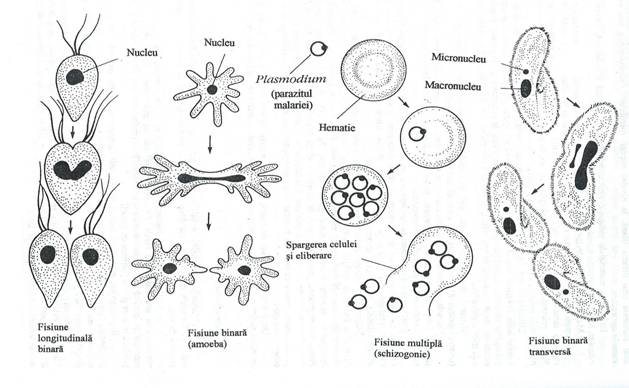
Fig. 157. Modalitatile de reproducere asexuata la protozoare.
Reproducerea asexuata se face si pe alte cai: inmugurire, fisiune multipla, urmatade citochineza. Prin inmugurire, la suprafata celulei parentale, se formeazao nouacelula, in care migreazaunul dintre nucleii rezultati din diviziunea mitotica. Mugurele creste progresiv, panase desprinde de celula parentala.
Fisiunea multipla, denumitaschizogonie, implicaformarea unui organism multinucleat. Nucleul si celelalte organite esentiale se divid repetat, faracitochineza. Ulterior, se produce diviziunea, prin care, simultan rezultaun mare numar de celule uninucleate. Fisiunea multiplaeste caracteristicasporozoarelor parazite.
Reproducerea sexuata este comunala protozoare. Unele sunt diploide in cea mai mare parte a existentei lor. Formarea gametilor este consecutivadiviziunii meiotice. Celulele sexuale (gametii) se unesc, fenomen denumit singamie si formeazazigotul diploid. Acest tip de reproducere se gaseste la sporozoare (Plasmodium).
La ciliate are loc un schimb reciproc de macronuclei haploizi, intr-un proces de sexualitate denumit conjugare, care implicastabilirea unei punti citoplasmatice intre cele douacelule conjugante. Micronucleul transferat de la donor, impreunacu micronucleul propriu, prin fuziune, formeazaun nucleu zigot (fertilizat).
Autogamia este o variantasimplificataa conjugarii, deoarece procesele nucleare se petrec in interiorul unei celule. Micronucleul se divide meiotic. Se formeazadoi micronuclei, care se reunesc pentru a forma un nucleu zigot. Ulterior, celula se divide prin fisiune simplasi rezultadouacelule, fiecare avand setul complet al structurilor nucleare.
O proprietate importantaa protozoarelor este capacitatea lor de a regenera o parte pierdutaprin excizie sau lezare. De remarcat este faptul caregenereazanumai portiunea care contine nucleul. Unele ciliate pot regenera intreaga celula, din circa l0% din volumul celulei originale, dacanucleul ramane intact. Citostomul parameciului nu se regenereazadupaamputare.
Clasificare
Principalele grupe de protozoare se disting usor pe baza criteriilor morfologice, a modului de obtinere a nutrientilor, a organitelor de locomotie, a organizarii celulare, la care se adaugaanaliza biochimicaa acizilor nucleici si a proteinelor, pentru a stabili gradul de asemanare sau de deosebire a unor specii foarte inrudite. Pe baza acestor criterii se disting urmatoarele grupe:
grupul Sarcodina, la care locomotia si captarea hranei se fac prin intermediul pseudopodelor;
grupul Mastigophora sau Flagellata posedacel putin un flagel, intr-un anumit stadiu al ciclului de viata, cu rol in locomotie;
grupul Sporozoa cuprinde protozoare parazite, care se hranesc prin absorbtia hranei din organismul gazda. Nu au organite de locomotie.
Grupul Ciliophora este caracterizat prin prezenta cililor, cu rol atat pentru deplasare, cat si pentru capturarea hranei.
Mastigophora (Flagelate)
Flagelatele se caracterizeazaprin prezenta unuia sau mai multor flageli, intr-un stadiu al ciclului de viata. Flagelii au rol in locomotie, pentru obtinerea hranei si sunt receptori pentru excitanti. Flagelatele sunt cele mai apropiate, filogenetic, de alge. Unele alge (Chlamydomonas, Volvox) sunt adeseori clasificate ca flagelate.
Unii autori clasifica flagelatele in douagrupe:
fitoflagelate (asemanatoare cu plantele) contin clorofilasi sunt fotosintetizante. Depoziteazaamidon si au perete celular celulozic. Reproducerea sexuataeste comuna;
zooflagelate (asemanatoare organismelor animale) nu au pigment fotosintetizant si sunt heterotrofe. Depoziteazaglicogen sau uleiuri. Nu au stadiul de reproducere sexuata.
Cele doua subgrupe se aseamanaprin prezenta flagelului ca organit de locomotie, dar deosebirile sunt majore si plasarea lor in aceiasi unitate taxonomicaeste artificiala.
Speciile g. Euglena au un statut aparte, datoritacapacitatii lor de a se hrani fotosintetizant sau heterotrof. Sub actiunea unor agenti chimici sau a radiatiilor se poate induce pierderea cloroplastelor. Mutantele acloroplastice se hranesc heterotrof.
Majoritatea flagelatelor sunt libere in apa dulce si sarata. Altele traiesc in sol sau in tractul intestinal al unor animale. Unele flagelate sunt parazite pentru om si animale, fiind foarte importante din punct de vedere medical. Cele mai importante mastigofore patogene sunt tripanosomele, agentii bolii somnului (T. brucei, T. gambiense). Lungimea lor este de 20 m, subtiri, cu formade semiluna, uniflagelate, cu originea intr-un corp bazal. Flagelul se orienteazaspre polul posterior, pe marginea celulei, fiind inclus intr-o cutaa membranei. Se formeazaun pliu membranar ondulator, ale carui miscari sunt coordonate de flagel. Membrana ondulantamareste eficienta deplasarii parazitului in sange, a carui vascozitate este mai mare.
T. brucei si T. gambiense sunt transmise prin intepatura mustei tete (Glossina). Netratata, victima devine somnolenta. Agentul patogen invadeaza sistemul nervos central si produce inflamatia tesutului nervos, determinantaa manifestarilor neurologice ale bolii. Parazitul se multiplicaprin fisiune binarain tractul intestinal al artropodului. Parazitul invadeaza glandele salivare, de unde este transferat la o noua gazda umana (fig. 158).
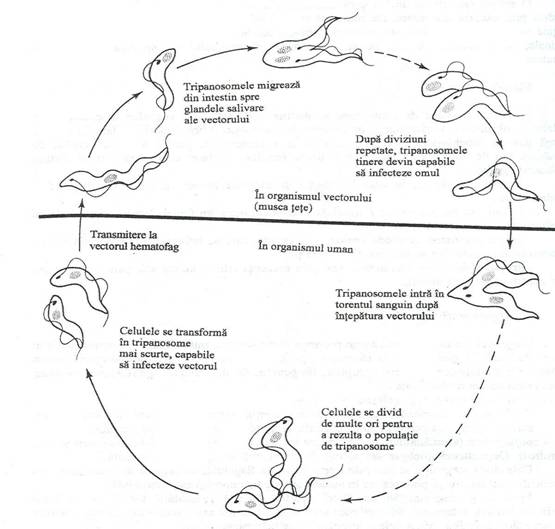
Fig. 158. Diagrama ciclului de viata la Trypanosoma. Dezvoltarea in organismul mustei tete este necesara pentru a produce forma infectioasa pentru organismul uman.
Leishmania sp. produce infectia cutanatala om, denumitaleishmanioza.
Giardia lamblia este unul dintre cele mai raspandite protozoare intestinale. Infecteazaomul si animalele. Parazitul se ataseazade celulele mucoasei intestinale. In numar mare, celulele parazitului, interferacu absorbtia substantelor nutritive. Produce tulburari digestive diareice, dar si manifestari nespecifice. Infectia este diagnosticataprin identificarea chistilor in materialele fecale. Sursa de infestare este apa sau alimentele vegetale contaminate de om sau de animalele bolnave. In comunitatile de copii, parazitul se transmite direct.
Trichomonas vaginalis este un flagelat mai raspandit decat G. lamblia si produce o infectie usoaraa vaginului.
Flagelatele din intestinul termitelor, constituie asociatii reciproc benefice. Termitele ingeralemnul, dar nu pot degrada celuloza. Flagelatele digeraceluloza si furnizeazaglucide simple, atat pentru ele insele cat si pentru gazda.
Sarcodina
Protozoarele cuprinse in grupul Sarcodina sunt mai simple ca structuradecat flagelatele si ciliatele, deoarece au mai putine diferentieri celulare. Locomotia si capturarea hranei se realizeazaprin intermediul pseudopodelor. La amoebe apare si miscarea flagelara, pentru caunele specii formeazasi celule flagelate in anumite conditii de mediu nefavorabil, astfel caorganismele se pot dispersa mai rapid.
Reprezentantii grupului Sarcodina traiesc in apa. Foarte interesante sunt subgrupele Foraminifera si Radiolaria. Ambele formeazacochilii. Cochiliile foraminiferelor sunt formate din calcar secretat de celula, iar la radiolari, cochilia se formeazadin materiale exogene. Celula nu este ferm atasatade cochilie si de aceea isi poate extinde pseudopodele prin unica sau numeroasele deschideri ale cochiliei.
La formele cochiliere, diviziunea celularaeste o reminiscentaa inmuguririi levurilor: se produce mitoza si unul dintre nuclei se deplaseazaintr-o portiune a citoplasmei, care se extruzeazaprin apertura cochiliei. In jurul citoplasmei extruzate se secretao nouacochilie si celula progenase separade celula parentala. Celula progenaare o cochilie noua, iar celula parentalao pastreazape cea veche.
Foaminiferele traiesc in regiunile de coasta, iar radiolarii, in larg. La multe foraminifere apare un ciclu sexual si in unele cazuri are loc o alternantade generatii: celulele haploide si diploide se divid si formeazalinii celulare care se reproduc independent. Unele celule diploide se divid meiotic si rezulta celule haploide care cresc vegetativ, inainte de a forma gameti. Acestia se imperecheaza si refac organismul diploid.
Datorita
greutatii cochiliei, celulele cad pe fundul apei si se
hranesc cu bacterii si detritus. Cochiliile acumulate pe fundul
oceanului formeaza roci sedimentare. Fiind foarte rezistente, se
fosilizeaza. In anumite conditii geologice, depozitele de cochilii
ale foraminiferelor se pot transforma in creta. Foraminiferele fosile sunt
foarte utile pentru geologi, in vederea evaluarii varstei rocilor de calcar.
Depozitele de foaminifere sunt utile pentru prospectiunile geologice
petroliere. Piramidele de langa
Amoeba proteus este un protozoar de apadulce, cu lungimea de 300 m. Are un nucleu mare, fapt care a usurat studiile de enucleare si inlocuire cu nucleul altei amoebe.
O larga varietate de amoebe este parazitala om si la alte vertebrate, cu localizare in cavitatea orala sau in tractul intestinal.
Entamoeba histolytica este parazita in tractul intestinal uman. In multe cazuri, infectia nu produce simptome evidente, dar la unii indivizi produce ulceratii ale tractului intestinal, insotite de diaree. Starea patologica provocata de infectie se numeste dizenterie amoebiana. Maladia este raspandita in toata lumea, dar prevaleaza in regiunile geografice cu conditii sanitare precare.
Parazitul este transmis de la un organism la altul, sub formade chist, prin contaminarea fecala a apei si a alimentelor. Odataajunsi in intestin, din chisti se elibereaza trofozoiti. Acestia se hranesc cu secretia mucoasasi cu bacterii. De aceea se dezvolta numai la organismele conventionale si nu la cele germ free. Trofozoitii, prin crestere, devin amoebe, care cresc si se divid prin fisiune binara. Amoebele produc enzime proteolitice, care lezeaza mucoasa intestinala. Astfel, amoebele patrund in grosimea peretelui intestinal, unde invadeaza vasele sanguine. Pe cale portala ajung in ficat, dar si in alte organe. Amoebele se multiplica si produc abcese tisulare.
Amoebele produc un efect iritant asupra celulelor mucoasei intestinale. Se intensifica peristaltismul si secretia de fluid intestinal. Diareea, uneori saguinolenta, datorata leziunilor mucoasei, este simptomul dizenteriei amoebiene. In fluidul diareic, pe o lamade microscop incalzita la 35-37o se observa trofozoitii mobili, cu diametrul de 20-40 m. La examenul fecal, trofozoitii nu se observa, dar chistii sunt prezenti. La trecerea prin intestinul gros, prin doua diviziuni succesive, chistii formeaza 4 nuclei si sunt infectiosi pentru gazda urmatoare. Chistii se identifica prin examenul direct al fecalelor persoanelor infectate. Daca sunt in numar prea mic, se pot detecta dupaconcentrare, prin diferite metode.
E. histolytica creste anaerob in culturi pure sau aerob in medii lichide care contin bacterii. Amestecul mai multor specii de bacterii favorizeaza cresterea amoebei. E. histolytica produce putine leziuni, dar acestea lipsesc la animalele germ free.
Antibioticele care inhiba dezvoltarea bacteriilor intestinale sunt eficiente fata de infectia amoebiana, chiar daca nu au efect direct asupra amoebelor. Aceasta denota dependenta amoebei de populatia bacteriana intestinala, pe seama careia se hraneste si chiar existenta unui sinergism al amoebei si bacteriilor pentru producerea dizenteriei amoebiene.
Sporozoa
Sporozoarele sunt un grup mare de protozoare, toate parazite obligate, unele pentru o singura gazda, altele pentru doua, in ciclul lor infectios in care se succed tipuri morfologice si fiziologice distincte.
Sporozoarele nu au organite de locomotie, desi unele formeaza pseudopode, iar uneori, gametii au mobilitate flagelara.
Modul de nutritie este particular: hrana nu este ingerata, ci este absorbitain formasolubila, prin invelisul extern, ca la bacterii si fungi.
Denumirea de "sporozoare" presupune formarea sporilor in ciclul de viata. Totusi, nu formeaza spori adevarati de tipul celor fungici, ci numai structuri analoge denumite sporozoiti, implicati in transmiterea la un nou tip de gazda.
Gazdele sporozoarelor sunt specii de nevertebrate, dar mai ales vertebrate. Unele prezinta o alternanta obligatorie a gazdelor, deoarece unele stadii ale ciclului se desfasoara la o gazda, iar alte stadii au loc in organismul altei specii de gazda.
Cei mai importanti membri ai grupului sunt coccidiile, parazite la pasari si plasmodiile (agentii malariei), infectioase pentru pasari si mamifere, inclusiv pentru om.
Plasmodium. Omul este infectat de 4 specii de Plasmodium care produc malaria: P. vivax, P. falciparum, P. malariae, P. ovale. Toate necesita o alternanta de gazde, om-tantar. Speciile de Plasmodium difera morfologic, prin unele aspecte ale ciclului de viata si prin severitatea malariei.
Cel mai raspandit este P. vivax. Parazitul isi desfasoara o parte a ciclului de viata la om si alta la tantar, vectorul care-l transmite de la om la om (fig. 159). Sunt implicati numai tantarii din g. Anopheles. Malaria este o boala infectioasa, foarte frecventa in zonele tropicala si subtropicala. Cea mai caracteristica trasatura a malariei este reactia febrila paroxistica, la intervale de 1-3 zile, care alterneaza cu stari fiziologice normale.

Fig. 159. Ciclul de viata al parazitului malariei (Plasmodium sp.) in organismul tantarului si al mamiferului
(dupa Phillips, 2000).
Omul este infectat prin intepatura femelei de tantar, care isi insera trompa direct intr-un capilar sanguin, unde inoculeaza sporozoitii. Sporozoitii plasmodiali sunt celule alungite care se formeaza in organismul tantarului si care se localizeaza in glanda salivara a insectei.
Dupa inocularea in gazda umana, prin intepatura, parazitul malariei este transportat in ficat unde paraseste circulatia si infecteaza celulele hepatice. Sporozoitii sunt transportati cu torentul sanguin in tot organismul, dar sunt retinuti de celulele sistemului fagocitar mononuclear din ficat, splina si din ganglionii limfatici. In ficat, sporozoitul creste si se transforma in schizont. Schizontul se fragmenteaza in mii de celule fiice mici, denumite merozoiti, care sunt eliberati din ficat in sange.
Unele specii de Plasmodium reiau ciclul de multiplicare hepatica, unde produc o infectie persistenta, de lunga durata, rezistenta la tratamentul infectiei sanguine si care se reactiveaza dupa intreruperea tratamentului.
Merozoitii infecteaza hematiile, initiind stadiul de schizont eritrocitar. La P. vivax, ciclul multiplicarii in hematie si liza ei dureaza 48 de ore. Stadiul multiplicarii eritrocitare este insotit de simptomele alternante de febra si frig. Senzatia de frig apare cand din eritrocite se elibereaza o noua generatie de paraziti.
Cele mai multe plasmodii tinere, eliberate din eritrocitul lizat, reiau ciclul infectios in noi eritrocite. O parte din plasmodiile eliberate din eritrocite nu pot sa infecteze alte eritrocite. Ele sunt celule specializate denumite gametocite si sunt infectioase numai pentru tantar. Gametocitele sunt celule sexuate care se deosebesc de celulele vegetative, atat ca aspect cat si ca sensibilitate la medicamentele antimalarice. Nu se dezvoltain organismul uman si nu au importantapentru producerea simptomelor malariei.
Gametocitele sunt ingerate de Anopheles odata cu sangele, prin intepatura. Infectia tantarului incepe odatacu ingestia celulelor sexuate ale parazitului. Gametocitele se matureazasi rezulta gameti masculi si femeli. Doi gameti cu polaritate sexuala opusa fuzioneaza si formeaza un zigot, care se deplaseaza prin miscare amoeboidala in peretele intestinului mijlociu al tantarului. Aici zigotul devine chist de reproducere, cu crestere progresiva prin diviziuni celulare. Una dintre diviziuni este meiotica si se formeaza un numar mare de celule asexuate, denumite sporozoiti. Acestia sunt eliberati, ajung in glanda salivara a tantarului, de unde sunt inoculati la o noua gazda umana. Infectia plasmodiala a tantarului nu interfera cu functiile fiziologice ale insectei, ceea ce denota ca raporturile lor sunt vechi si in evolutie s-a produs o selectie a variantelor de Plasmodium, capabile sa convietuiasca cu insecta. La om insa, malaria este o boala grava, uneori mortala.
Plasmodium nu a fost cultivat pe medii artificiale. Studiile s-au facut pe stadiul infectios eritrocitar. Hematiile infectate pot fi separate din sange si pot fi incubate in vitro, in conditii care permit multiplicarea parazitului. Parazitul are propriul sistem enzimatic generator de energie. Nu sintetizeaza CoA, pe care o preia din eritrocit. Sintetizeaza acidul folic, daca este disponibil precursorul acestuia, acidul para-amino-benzoic. Pentru sinteza proteinelor proprii, parazitul foloseste aminoacizii derivati din hemoglobina, cea mai abundenta proteina eritrocitara (circa 90%). Hemoglobina este endocitata prin pinocitoza si este hidrolizata la amnoacizi. Hematia mamiferelor nu contine ADN, ci numai mici cantitati de ARN, ceea ce denota ca, probabil, parazitul sintetizeaza bazele purinice si pirimidinice, pornind de la aminoacizi. Probabil capermeabilitatea foarte mare a membranei celulare, permite preluarea unor molecule mari din eritrocit. Permeabilitatea crescuta, determina pierderea rapida a macromoleculelor in mediul extracelular, ceea ce explica parazitismul sau obligat si imposibilitatea de a creste pe medii artificiale.
Probabil exista diferente fiziologice intre stadiul infectios pentru eritrocite si cel infectios pentru ficat. Schizontul eritrocitar pare adaptat sa utilizeze numai hemoglobina ca sursa de aminoacizi.
In zonele calde ale globului, unde tantarii sunt numerosi, malaria este o infectie endemica. In aceste regiuni, indivizii umani dobandesc rezistentala infectia cu Plasmodium. In Africa de Vest, rezistenta la malaria cauzata de P. falciparum este asociata cu prezenta in hematii, a hemoglobinei S, care difera de hemoglobina A, printr-un singur aminoacid in cele doua jumatati simetrice ale moleculei. In hemoglobina S, aminoacidul neutru valina este inlocuit cu un aminoacid acid, acidul glutamic. Hemoglobina S are afinitate scazuta pentru O2, ceea ce creeaza conditii nefavorabile pentru Plasmodium, care are un metabolism strict aerob si nu se dezvolta la fel de bine in hematiile S, ca in hematiile cu hemoglobinanormalaA. Corelat cu afinitatea mai mica a hemoglobinei S fata de O2 este faptul ca indivizii suporta greu altitudinile mari, la care presiunea O2 este mai mica, dar in regiunile joase, dezavantajul creat de hemoglobina S nu este evident.
In unele regiuni mediteraneene, unde malaria este endemica, rezistenta la P. falciparum este asociata cu deficienta in hematii, a enzimei glucozo-6-fosfat-dehidrogenaza.
Pe baza corelatiilor dintre tipul de hemoglobina si gradul de sensibilitate la infectia cu Plasmodium, se considera ca parazitul malariei a fost unul dintre factorii importanti in evolutia sub raport biochimic a speciei umane.
Un alt sporozoar cu un ciclu complex de viataeste Toxoplasma gondii, agentul toxoplasmozei la pasari si mamifere, la care a produs epidemii cu pierderi economice importante.
T. gondii infecteaza celulele mucoasei intestinale ale pisicii. Procentul pisicilor infectate pare sa fie foarte mare (circa 50%). Alte animale sau omul se infecteaza prin ingestia hranei sau apei, contaminata cu fecalele animalelor infectate. Parazitul strabate peretele intestinal si pe cale sanguinaajunge in tesuturile gazdei (vaca, oaie etc.) unde se dezvolta intr-o forma chistica, in care prin diviziune, rezulta multe celule progene infectioase. Ele raman viabile pentru o perioada nedefinita si sunt transmise la om, prin ingestia carnii insuficient pregatita. Studiile epidemiologice au aratat ca persoanele care traiesc in contact cu pisica au o incidenta mai mare a toxoplasmozei.
Ciliophora
Ciliatele sunt protozoare care se caracterizeaza prin prezenta cililor pe suprafata intregii celule sau numai in zone specializate ale ei. Din punct de vedere structural si functional, celulele ciliatelor sunt printre cele mai complexe si specializate din lumea vie, deoarece poseda organite ce realizeaza functii vitale particulare, de ingestie a hranei, de eliminare a resturilor indigeste, de mentinere a echilibrului osmotic, de miscare, de receptie a stimulilor etc.
Cele circa 6 000 de specii sunt impartite in doua grupe:
cele care au cili numai pe o zona a suprafetei celulare (Stentor);
cele care au cili distribuiti pe toata suprafata celulei (Paramecium).
Ciliatele sunt foarte raspandite in apele dulci, iar parameciul este abundent in apele statatoare.
Ciliatele sunt unice printre protozoare, deoarece au doi nuclei: micronucleul, cu rol de a transmite caracterele ereditare si pentru realizarea fenomenului de sexualitate; macronucleul, care nu are rol in transmiterea caracterelor ereditare, dar codifica sinteza ARNm si coordoneaza toate functiile celulei.
Macronucleul este poliploid, avand multe copii ale informatiei genetice a celulei. Contine de circa 500 de ori mai mult ADN decat micronucleul. La unele cilitate, macronucleii sunt multipli.
Majoritatea ciliatelor se hranesc, ingerand materiale particulate printr-o regiune diferentiata, denumita citostom. In jurul regiunii orale se gasesc cili, care prin miscarea lor coordonata, creeaza un curent al apei ce deplaseaza particulele alimentare spre orificiul oral. Citostomul se inchide si se deschide, ca raspuns la absenta sau prezenta particulelor alimentare. Particula alimentara este transportata prin citofaringe, in citoplasma, unde este inchisa intr-un sistem membranar si se formeazao vacuola digestiva, in interiorul careia se elibereaza enzimele hidrolitice.
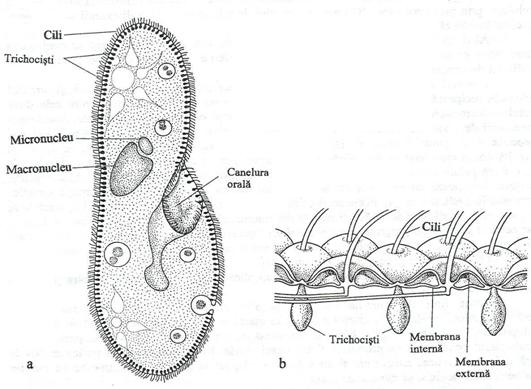
Fig. 160. a. Organizarea celulara la Paramecium. b. Numerosi cili emerg prin membrana interna si externa a peliculei. in repaus, trichocistii sunt localizati sub pelicula.
Vacuola digestiva este delimitata de o membrana externa foarte fina si este transportata intracelular, prin curenti citoplasmatici. Vacuola digestiva se opreste totdeauna un interval de timp, in apropierea nucleului. Continutul vacuolei are, initial, o reactie acida. Odata cu progresia digestiei, continutul devine alcalin, prin hidroliza componentelor proteice. Continutul vacuolei se pierde in regiunea porului anal, unde are loc excretia produselor de uzura. Produsele de catabolism (apa, CO2, uree etc.) sunt eliminate la suprafata celulei, printr-o structura specializata, denumita vacuola contractila, o structura permanenta sau temporara.
Un alt organit specializat al ciliatelor este trichocistul, o structura filamentoasa, subtire, ancorata sub invelisul celular si se formeaza dintr-o grupare de cili. El poate fi proiectat sau retractat si serveste ca mijloc de ancorare, arma de aparare sau instrument de captare a prazii. La Didinium, un ciliat pradator, trichocistul paralizeaza prada, inainte de a o ingera.
Ciliatele se deosebesc net de alte protozoare, prin prezenta celor doua tipuri de nuclei: micronucleul diploid si macronucleul poliploid. Micronucleul nu are rol direct in cresterea si diviziunea celulei. Celula fara micronucleu continua sa creasca si sa se divida. Dupa indepartarea macronucleului, celula moare repede. Dupa excizia unei parti a macronucleului, fragmentul ramas, regenereaza macronucleul intreg.
La ciliate se produc doua feluri de fenomene nucleare: autogamia si amfimixia.
In procesul de autogamie are loc reorganizarea nucleara, in absenta proceselor sexuale. Semnificatia fiziologica a procesului este prevenirea senescentei si consta in regenerarea aparatului nuclear, fara conjugare. Macronucleul degenereaza, iar micronucleul se divide repetat si se organizeaza un alt macronucleu. Ca rezultat al autogamiei, celula devine homozigota, deoarece toate genele ambilor tipuri de nuclei provin din micronucleul haploid. Datorita repetarii autogamiei pentru mai multe generatii, se produce senescenta (imbatranirea) populatiei celulare, prin pierderea functiei macronucleului. Rata diviziunii scade treptat si chiar inceteaza.
Al II-lea fenomen nuclear, care are loc la ciliate este amfimixia si implica conjugarea intre doua celule ce apartin unor "tipuri sexuale" diferite. Fecundarea nu se produce intre indivizii descendenti din aceiasi celula parentala.
Procesul conjugarii este precedat de recunoasterea celor doi parteneri si implica aderenta reciproca in zona citostomului si realizarea unei punti de legatura intre cele douacelule. Macronucleul degenereaza treptat, iar micronucleul sufera modificari ample, de-a lungul unei serii de diviziuni succesive. Prima diviziune este mitotica. Cea de a II-a este meiotica si produce 4 micronuclei haploizi. Din cei 4, trei degenereaza. Cel care ramane, se divide si rezulta doi micronuclei de fecundatie sau pronuclei. Aceste evenimente se produc simultan in cele doua celule conjugante. Are loc un schimb reciproc de pronuclei haploizi. Micronucleul exogen fuzioneaza cu cel autohton si rezulta un nucleu diploid, a carei informatie genetica provine, in egala masura, din cele doua celule conjugante. Dupa schimbul de nuclei, celulele se separa. Nucleul diploid se divide si rezulta doi micronuclei. Unul ramane micronucleu diploid, iar celalalt se mareste progresiv prin endomitoze succesive, isi mareste numarul de cromosomi si atinge un grad inalt de poliploidie. Rezultatul net al conjugarii este formarea exconjugantilor, care sunt hibrizi pentru genele doi conjuganti.
Conjugarea nu este o modalitate de multiplicare, ci este o cale de reinoire partiala a garniturii de cromosomi.
Multe linii de Paramecium (dar si alte protozoare) contin alge sau bacterii endosimbiotice, localizate in citoplasma sau in macronucleu. De exemplu, la P. bursaria, in citoplasma, se gaseste alga Chlorella. In acest raport simbiotic, alga este protejata, iar protozoarul beneficiaza de produsele fotosintezei algale. Simbiontii algali ai protozoarelor de apa dulce se numesc zooclorele si au culoare verde, iar ai protozoarelor marine au culoare galbena sau cafenie si se numesc zooxantele.
Paramecii au o varietate de endosimbionti de natura bacteriana. P. aurelia contine celule bacteriene cunoscute sub denumirea de particule kappa. Endosimbiontul secretao substanta toxica, ce produce efect litic asupra paramecilor sensibili, lipsiti de particulele endosimbionte bacteriene si de aceea au fost denumiti parameci killer.
Alt exemplu de endosimbiont bacterian, este particula lambda, care a fost cultivatasi este un bacil Gram negativ.
Rolul bacteriilor, algelor si fungilor endosimbiotici este incert, dar exista dovezi ca endosimbiontii au rol nutritiv pentru ca sintetizeaza vitamine si alti factori de crestere pe care protozoarele nu-i pot sintetiza si necesita aportul lor exogen.
Unul dintre cele mai mari ciliate este Stentor, cu lungimea de 1 mm, vizibil cu ochiul liber. Este sesil, fixat pe substrat prin intermediul unui peduncul. Deschiderea citofaringelui este tapetata cu un sir de cili, care prin miscare ordonata, orienteaza hrana din curentul de apa, spre citofaringe.
Diviziunea celulei este un proces complex. In timpul diviziunii, una din celulele fiice pastreaza citofaringele matern si formeazaun nou crampon, iar cealalta formeaza un citofaringe nou si pastreaza cramponul vechi.
Opalina ranarum este un ciliat parazit in intestinul unor broaste. Diviziunea nucleului nu este sincrona cu diviziunea celulei, ci permanenta, pe toata durata cresterii celulei. Opalinele se deosebesc de ciliatele propriu-zise, prin aceea caau doi nuclei egali. Lipseste micronucleul.
In compartimentul ruminal al rumegatoarelor, se gaseste o asociatie diversificata si caracteristica de ciliate, care au rol benefic in procesul digestiei masei vegetale si in aprovizionarea organismului animal cu proteine.
Un alt grup de ciliate, cu morfologie distincta este Suctoria. Celulele libere formeaza o parte a planctonului. Pe masura ce se matureaza, pierd cilii si se fixeazade suport printr-un peduncul sau un disc. Membrii grupului se gasesc in apa dulce si sarata, fixati pe plante si animale. Isi obtin hrana din tesutul suport, prin aspiratie, cu ajutorul unor tentacule protoplasmatice fine, care penetreaza stratul protector al tesutului gazdei. Se multiplica prin inmugurire.
Foarte putine ciliate sunt parazite la om si animale. Balantidium coli este parazit la animalele domestice, dar infecteaza tractul intestinal uman. Simptomele produse sunt similare cu cele cauzate de E. histolytica.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 14641
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved