| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Quinolonele
Quinolonele (denumite si 4-quinolone) sunt primele substante antimicrobiene obtinute pe cale sintetica si formeaza o familie de compusi care se aseamana prin existenta nucleului quinolinic. Primul compus din acest grup, folosit in terapie este acidul nalidixic (Fig. 78).
Quinolonele, alaturi de - lactamice si macrolide, reprezinta una dintre cele trei familii principale de agenti antimicrobieni folositi in terapeutica umana. Importanta lor terapeutica este in continua crestere incepand din 1968, data comercializarii primei quinolone reprezentata de acidul nalidixic.
Acidul nalidixic este un produs intermediar de sinteza a quinolonelor. Ulterior quinolonele s-au diversificat prin introducerea unui atom de fluor (F) in pozitia 6 si a unui heterociclu in pozitia 7 (piperazine, pirolidina, etc.) care au generat fluoroquinolonele. Aceste molecule poseda un spectru antibacterian foarte larg si pot fi divizate in molecule metabolizabile si nemetabolizabile (grupele III si IV).
Quinolonele se pot clasifica in doua grupe :
cele de prima generatie, ca acidul nalidixic, active asupra bacililor Gram-negativi;
fluoroquinolonele
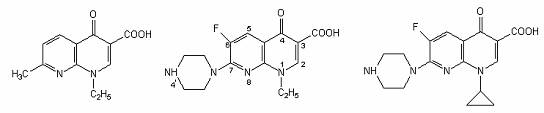
Acid nalidixic Norfloxacin Ciprofloxacin
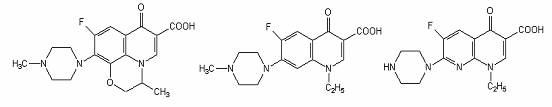
Ofloxacin Pefloxacin Enoxacin
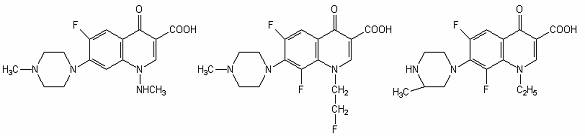
Amifloxacin Flerofloxacin Lomefloxacin
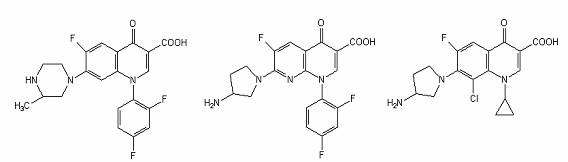
Temafloxacin Tosufloxacin PD 127, 391
Agenti antimicrobieni fluoroquinolonici.
Gama derivatilor quinolonei s-a diversificat prin modificarea nucleului de baza, 4-quinolona. La atomul C6 s-a adaugat unul de fluor, ceea ce a crescut semnificativ spectrul si potentialul lor antimicrobian.
Avand in vedere spectrul lor de activitate antibacteriana, limitat la bacterii Gram negative si in principal la enterobacterii, acidul nalidixic si derivatii sai au fost folositi pentru tratamentul infectiilor urinare. Modificarile structurii au dat nastere la quinolone, denumite noile quinolone sau fluoroquinolone (norfloxacin, pefloxacin, ofloxacin, ciprofloxacin etc.), al caror spectru de activitate antibacteriana se extinde la alte specii Gram negative (de ex. Pseudomonas aeruginosa), dar si la anumite specii Gram pozitive (S. aureus, micobacterii).
Totusi, activitatea noilor quinolone fata de alte specii, asa cum sunt cele natural-sensibile la acidul nalidixic, ramane modesta, ceea ce corespunde unui anumit grad de rezistenta intrinseca a acestor specii.
Mecanismul de actiune a quinolonelor este foarte complex. Aceste molecule patrund in celula bacteriana prin difuzie pasiva si actioneaza asupra tintelor specifice reprezentate de topoizomeraze*: ADN-giraza (topoizomeaza II) si topoizomeraza IV. Actiunea celor doua enzime este inhibata. Ambele sunt proteine heterotetramerice, alcatuite din 2 subunitati A si 2 subunitati B. Quinolonele se leaga si stabilizeaza complexele giraza-ADN (quinolona singura nu se asociaza cu ADN), dupa clivarea lantului, impiedicand actiunea catalitica a ADN-polimerazei la nivelul bifurcatiei de replicare. Complexul genereaza o rupere a moleculei de ADN, pe care celula nu o repara eficient.
Formarea suprahelicei moleculei de ADN al cromosomului bacterian si relaxarea ei este conditionata de activitatea unui set de enzime, denumite ADN-topoizomeraze. Topoizomerazele sunt enzime care modifica conformatia spatiala a ADN prin ruperea si reunirea catenelor. Bacteriile poseda patru clase de topoizomeraze (I - IV). O topoizomeraza este o nucleaza reversibila, care se leaga covalent la o grupare fosfat a ADN si rupe legatura fosfodiesterica. Deoarece legatura covalenta care uneste topoizomeraza la o grupare fosfat a ADN retine energia legaturii fosfodiesterice pe care o rupe, reactia este reversibila, adica incizia este urmata de legarea celor doua capete. Legarea este rapida si nu necesita o sursa suplimentara de energie.
Topoizomerazele de tip 1 actioneaza prin recunoasterea unui segment de ADN, partial despiralizat, prin incizia unei catene, ceea ce permite celor doua parti ale helicei de ADN, de o parte si de alta a inciziei, sa se roteasca liber una fata de alta, in sensul care reduce tensiunea de supraspiralizare. Aceasta inseamna ca replicarea ADN se face numai cu rotatia unei mici parti a helicei, adica a celei situata in aval de bifurcatie. Problema transcrierii se rezolva in acelasi mod.
Topoizomerazele de tip II se leaga covalent, simultan, de cele doua catene ale dublei helice si produc o rupere bicatenara tranzitorie. Aceste enzime se activeaza la situsurile cromosomale la nivelul carora se intrepatrund doua duble helice.
Dupa fixarea topoizomerazei la un astfel de situs, etapele actiunii sale sunt urmatoarele:
clivarea uneia dintre cele doua helice duble;
enzima determina trecerea celei de a II-a catene, prin deschiderea creata;
repara discontinuitatea inainte de a se disocia de ADN.
ADN-polimeraza de tip II poate astfel sa separe cele doua molecule de ADN catenate.
Unele topoizomeraze sunt helicaze sau giraze(produc spiralizarea moleculei de ADN), iar altele sunt derulaze (produc despiralizarea prin incizia unei catene si bucla se relaxeaza).
Fluoroquinolonele formeaza complexe stabile cu topoizomeraza II, efectul fiind moartea celulei. S-a sugerat ca quinolonele nu se leaga cu ADN-giraza insasi, ci probabil chiar la situsuri specifice pe ADN, create de ADN-giraza
Studiile comparative ale sensibilitatii la fluoroquinolone si de dezvoltare a rezistentei, au relevat ca ADN-giraza este tinta primara a fluoroquinolonelor la bacteriile Gram negative, iar topoizomeraza IV este tinta primara la bacteriile Gram pozitive. Exceptia o constituie S. pneumoniae, la care fie giraza, fie topoizomeraza pot fi tinte primare, in functie de fluoroquinolona folosita.
Activitatea antibacteriana este dependenta intr-o masura semnificativa de atomul de fluor din pozitia 6 si de nucleul piperazinic din pozitia 7. Configuratia spatiala a quinolonei determina nivelul activitatii. Astfel, enantiomerii stereochimici (care difera unul de altul numai prin pozitia in spatiu a unei grupari particulare), ce implica grupul metil atasat la inelul al III-lea de ofloxacin, au activitati antibacteriene foarte diferite, care difera in proportie de 1/10.
Celulele eucariote contin topoizomeraze care au omologie limitata a aminoacizilor cu ADN-giraza si cu topoizomeraza IV.
Cele 2 enzime au localizare citoplasmatica si quinolonele trebuie sa traverseze structurile de suprafata ale celulei bacteriene.
Quinolonele sunt agenti bactericizi. Ele stopeaza rapid sinteza replicativa a ADN si intrerup progresia bifurcatiei de replicare. Inhibitia activitatii ADN-girazei sub actiunea fluroquinolonelor induce moartea rapida a celulei bacteriene. Inhibitia rapida a sintezei ADN nu explica moartea celulei bacteriene. Pentru efectul letal sunt necesare alte evenimente suplimentare: inhibitia sintezei ARN si a proteinelor. La concentratiile de quinolone care depasesc un anumit prag, activitatea bactericida diminua, probabil pentru ca este inhibata numai sinteza ARN si a proteinelor.
Tratamentul cu quinolone, probabil induce efecte pleiotrope, ce pot fi consecinte secundare ale inhibitiei sintezei ADN-girazei: leziuni ale ADN bacterian, deoarece quinolonele sunt inductoare ale sistemului reparator SOS, dependent de Rec A.
ADN giraza sau topoizomeraza II este o proteina heterotetramerica formata din doua subunitati A(gyr A) si doua subunitati B(gyr B). (La E. coli proteinele gyr au 97 kDa).
Situsul catalitic a ADN girazei este situat la tirozina din pozitia 122 a subunitatii A.
Subunitatea B cuprinde situsul de hidroliza a ATP, hidroliza care furnizeaza energia necesara activitatii enzimatice.
Subunitatile A si B ale topoizomerazei II sunt codificate de genele gyrA si gyrB.
Dupa purificarea ADN-girazei de E. coli, structura acestei enzime a fost investigata pentru numeroase alte specii bacteriene, evidentiindu-se un grad inalt de omologie intre subunitatile A pe de o parte si a subunitatilor B pe de alta parte. Cu toate acestea, secventa situsurilor catalitice din proteinele gyr A si aceea a situsului de hidroliza a ATP in proteinele gyr B sunt foarte conservate. Genele gyr A si gyr B sunt gene structurale ale subunitatilor A si B ale ADN-girazei.
Subunitatea A a fost desemnata ca tinta preferentiala a actiunii quinolonelor. Subunitatea B este tinta altor antibiotice: cumermicina si novobiocina.
ADN-giraza purificata introduce rasuciri suprahelicale negative ale moleculei de ADN circulara inchisa si separa reversibil moleculele circulare catenate. Aceste activitati sunt dependente de energia eliberata prin hidroliza ATP si constau in clivarea ambelor catene ale moleculei de ADN, trecerea altui duplex de ADN (sau alt segment al aceluiasi duplex) si reunirea catenelor. Activitatea ADN-girazei este inhibata de quinolone.
ADN-giraza este o topoizomeraza, singura enzima care supraspiralizeaza molecula de ADN, adica modifica configuratia spatiala a moleculei, prin catalizarea suprarasucirilor negative ale ADN cromosomal si plasmidial, usurand impachetarea cromosomului bacterian in spatiul restrans al celulei. ADN-giraza este singura enzima care influenteaza gradul de spiralizare al ADN, fiind esentiala pentru mentinerea starii suprahelicale a cromosomului bacterian. Inhibitia activitatii acestei enzime de catre fluoroquinolone este asociata cu moartea rapida a celulei bacteriene. Proteina se leaga de ADN ca un tetramer, in care cele 2 subunitati A si 2 subunitati B impacheteaza ADN prin supraspiralizare negativa.
ADN-giraza utilizeaza energia rezultata prin hidroliza ATP si este esentiala pentru mai multe procese vitale: initierea si progresia bifurcatiei de replicare, terminarea replicarii ADN, transcrierea unor operoni, repararea ADN, recombinarea si transpozitia.
Aceste activitati sunt rezultatul sectionarii coordonate a ambelor catene ale ADN, trecerea celuilalt segment de ADN prin nisa si restabilirea continuitatii catenei. Mecanismul de actiune este caracteristic topoizomerazei II.
ADN giraza elimina rasucirile suprahelicale pozitive care se acumuleaza inaintea bifurcatiei de replicare.
Topoizomeraza IV a fost descrisa recent, iar functia sa principala este decatenarea, adica separarea copiilor ADN circular dublu catenar dupa replicarea cromosomului bacterian si a plasmidelor. Topoizomeaza IV este omologa structural cu ADN giraza. Este o enzima de separare a catenanilor (a moleculelor surori catenate de ADN), rezultati dintr-un rund de replicare bidirectionala si permite segregarea lor in celulele surori.
ADN-giraza si topoizomeraza IV actioneaza asupra dublei catene, dar efectele sunt diferite: giraza impacheteaza ADN prin inducerea supraspiralizarii, iar topoizomeraza IV separa moleculele reunite prin legaturi intermoleculare.
Topoizomeraza IV este formata la fel ca ADN-giraza din doua subunitati denumite Par C si doua subunitati Par E, cu aceeasi repartitie functionala ca si a subunitatilor ADN-girazei. Proteinele Par C si Par E sunt foarte asemanatoare prin structura lor primara cu proteinele Gyr A si Gyr B (40% din secventa aminoacizilor este identica) si sunt codificate de genele parC si parE.
Topoizomeraza IV modifica intr-o masura mult mai mica topologia ADN dublu catenar: rolul sau este important pentru separarea catenelor de ADN dupa terminarea replicarii.
Rolul topoizomerazei IV, ca tinta specifica a quinolonelor a fost recent demonstrat la E. coli (Koto, 1990), S. aureus (Ferrero, 1994) si N. gonorrhaeae (Belland, 1994).
Noile quinolone au reprezentat un real progres terapeutic avand in vedere caracteristicile lor antibacteriene (spectru larg, activitate bactericida si farmacocinetica). Aceasta explica spectaculoasa crestere a utilizarii lor in ultimii 10 ani. Dar utilizarea extensiva a noilor quinolone s-a tradus prin emergenta ingrijoratoare a tulpinilor rezistente a unor specii bacteriene de mare importanta medicala (enterobacteriile, P. aeruginosa, S. aureus, M. tuberculosis, Neisseria gonorrhoeae) si a tulpinilor multirezistente la alte antibiotice.
Mecanismele rezistentei bacteriene la fluoroquinolone sunt de trei categorii:
modificari ale enzimelor tinta ale medicamentelor
alterari care limiteaza accesul medicamentelor la tinta
activitatea pompelor de efux.
Rezistenta la quinolone este, predominant, consecinta modificarilor enzimei tinta, la situsurile active ale enzimei. La bacteriile Gram negative, ADN-giraza pare a fi tinta primara pentru toate quinolonele. La bacteriile Gram pozitive, quinolonele, in functie de compusul chimic, actioneaza asupra topoizomerazei IV sau ADN-girazei. Structura quinolonei determina specificitatea tintei de actiune asupra bacteriilor.
Rezistenta speciilor Gram pozitive la quinolone se datoreaza in special mutatiilor intr-o regiune specifica (quinolone resistance determining region - QRDR) a subunitatii A a ADN-girazei. QRDR este regiunea N-terminala a proteinei, omologa cu regiunile GyrA si ParC de la E. coli. Regiunea cuprinsa intre codonii 67 - 106 ai GyrA la E. coli, este determinanta pentru rezistenta la quinolone. Mutatiile genei gyrA induc schimbari ale situsului de legare/sau ale sarcinii, care conditioneaza interactiunea ADN-girazei cu quinolona. Quinolonele interactioneaza in primul rand cu subunitatea A, dar s-au identificat mutatii ale subunitatii B care confera rezistenta la quinolone.
ADN giraza si topoizomeraza IV sunt localizate in citoplasma bacteriana. Pentru a-si atinge tinta, antibioticele fluoroquinolonice trebuie sa traverseze invelisul celular. Modificarile structurale ale membranei externe a bacteriilor Gram negative asociate cu diminuarea inglobarii sunt factori importanti ai rezistentei la fluoroquinolone.
Variantele Gram negative rezistente la quinolone care se selecteaza, se datoreaza modificarii porinelor din membrana externa, asociata cu scaderea permeabilitatii. Rezistenta la quinolone nu este transferabila prin intermediul plasmidelor. La bacteriile Gram pozitive, scaderea ratei inglobarii nu s-a demonstrat a fi un mecanism al rezistentei.
Atat bacteriile Gram pozitive, cat si cele Gram negative pot dobandi un nivel scazut al rezistentei, mediat de activitatea pompelor de eflux, cu rol de transportori multipli, a caror activitate este dependenta de gradientul electrochimic (forta proton motrice).
Nu s-au identificat enzime cu efect inactivator fata de quinolone, asa cum β-lactamaza inactiveaza penicilina sau cefalosporinele.
Ciprofloxacinul are un potential antibacterian mult mai ridicat decat acidul nalidixic.
Ciprofloxacinul se foloseste in special pentru tratamentul infectiilor respiratorii, ale tractului urinar, gonoreii, pentru tratamentul infectiilor diareice cu tulpinile enterotoxice de E. coli, Campylobacter jejuni, Shigella. Este activa, in vitro, fata de M. tuberculosis. Este partial metabolizata de ficat si excretata de rinichi. Medicamentele antiacide (Maloox) blocheaza absorbtia ciprofloxacinului si nu vor fi administrate concomitent. Administrata impreuna cu teofilina, poate duce la acumularea unor nivele sanguine inalte ale teofilinei. Teofilina se foloseste ca bronhodilatator in tratamentul astmului. Nivelul crescut al teofilinei poate produce atacul cerebral si tulburari de ritm cardiac.
Ionii de Ca2+, Cu2+, Fe2+, Mn2+, Mg2+, Zn2+ pot sa lege ciprofloxacina si diminua mult absorbtia medicamentului.
Multe antibiotice, inclusiv ciprofloxacinul, pot altera microbiota normala a colonului, favorizand dezvoltarea bacteriilor care produc inflamatia mucoasei (colita pseudomembranoasa) si determina tulburari diareice. Colita pseudomembranoasa se datoreaza cresterii in exces a bacteriei Cl. difficile si poate induce stari febrile, durere abdominala, tulburari ale tranzitului intestinal si chiar starea de soc. Tratamentul cu ciprofloxacina poate sa duca la cresterea in exces a levurii C. albicans, cu localizare vaginala (vaginita) sau intestinala (disbioza).
Manifestarea efectelor secundare este prevenita prin administrarea probioticelor: Lactobacillus casei, L. acidophilus, Bifidobacterium longum, Saccharomyces cerevisiae.
Derivatii quinolonici se folosesc pentru tratamentul infectiilor cailor urinare, unde, dupa administrare orala, se acumuleaza in concentratii inhibitorii. Fluoroquinolonele patrund in tesutul prostatic la concentratii care echivaleaza sau depasesc de cateva ori pe cele din plasma.
Cele mai sensibile la actiunea quinolonelor sunt enterobacteriile, dar compusii grupului sunt activi fata de chlamidii si micobacterii. Efectul bactericid al ciprofloxacinului este rapid, cu o pierdere de 90% a viabilitatii, intr-un interval de l9 minute.
Quinolonele au efect antimicrobian prelungit dupa administrare, ceea ce se reflecta in continuarea supresiei cresterii bacteriene, dupa eliminarea agentului antimicrobian din organism. Daca un medicament are un efect persistent dupa administrare, inseamna ca poate fi eficient chiar in intervalele dintre doze, cand nivelul seric si tisular au scazut sub nivelul concentratiei minime inhibitorii.
Quinolonele isi pastreaza activitatea fata de multe bacterii rezistente la antibiotice, inclusiv fata de bacilii Gram negativi cu rezistenta multipla, fata de S. aureus rezistent la meticilina, fata de N. gonorrhoeae rezistent la penicilina, fata de H. influenzae producator de b-lactamaze.
Ciprofloxacina, ofloxacina si tosufloxacina sunt active fata de Chlamydia trachomatis si Mycoplasma hominis. Quinolonele sunt de asemenea active fata de Rickettsia spp.
Acidul nalidixic inhiba numai speciile de bacterii Gram negative aerobe. In molecula de ciprofloxacina, fluorul confera activitate fata de bacteriile Gram pozitive. Grupul piperazinic creste activitatea fata de enterobacterii, iar gruparea piperazina si ciclopropil confera activitate fata de speciile de Pseudomonas.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1508
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved