| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Reglarea activitatii celulei bacteriene
Activitatea metabolica a celulei bacteriene se adapteaza permanent conditiilor de mediu mereu schimbatoare, prin intermediul unor mecanisme fine si eficiente.
Simplitatea organizarii interne a celulei bacteriene si deosebita sa plasticitate fiziologica ii asigura supravietuirea in contextul unor variatii largi ale conditiilor de mediu, spre deosebire de celulele organismelor superioare, care tolereaza modificari in limite foarte restranse ale parametrilor homeostaziei. Celula bacteriana se gaseste intr-o permanenta stare de echilibru dinamic, in care toate componentele sale structurale sunt continuu construite si degradate.
Bacteriile sintetizeaza unele enzime necesare degradarii anumitor substraturi, numai cand acestia sau analogii lor se gasesc in mediu.
In mod obisnuit, bacteriile necesita produsele de sinteza ale unui singur operon. In celula functioneaza mecanisme reglatoare de tip on-off care permit sinteza enzimelor, numai daca acestea sunt cerute de conditiile externe.
Selectia naturala optimizeaza eficienta. Sistemele reglatoare ale celulelor procariote au o finalitate teleonomica precisa: sa favorizeze cresterea cu o rata maxima, cu exceptia cazurilor in care cresterea ar fi daunatoare. Aceiasi strategie a finalitatii se aplica si organismelor eucariote unicelulare: levuri, alge, protozoare.
Reglarea celulara la bacterii este supusa unor reguli generale:
moleculele care sunt necesare numai ocazional, se sintetizeaza numai cand prezenta lor este necesara;
o activitate enzimatica ce consuma inutil energie, sau consuma o molecula care este substratul unei alte enzime, este de obicei inhibata;
daca celula are mai multe cai metabolice disponibile, ea va alege calea care produce o cantitate mai mare de energie pe unitatea de timp.
Schimbarile chimice in lumea vie se produc pe cale metabolica, adica printr-o secventa de reactii, fiecare fiind catalizata de o enzima specifica.
La bacterii, reglarea activitatii unei astfel de caI, se face prin sinteza sau inhibitia sintezei intregului set de enzime si proteine accesorii: ori se sintetizeaza intregul set de enzime, ori nu se sintetizeaza nici una. Acest fenomen, denumit reglare coordonata deriva din controlul sintezei unei molecule de ARNm policistronic, care codifica toate produsele genice.
Un operon functioneaza cand este necesar (on) si se intrerupe (off) cand enzimele codificate sunt inutile pentru celula.
Dupa Kleinsmith (l976), activitatea mecanismelor reglatoare se poate desfasura la 4 nivele:
la nivelul transcrierii genetice (prin sinteza ARN sau prin blocarea ei);
prin controlul la nivelul traducerii genetice;
prin modificarea directa a activitatii enzimelor existente in celula;
prin modificarea ratei degradarii proteinelor.
Controlul la nivelul traducerii mesajului, este un eveniment mult mai rar, dar cele de reglare la nivelul transcrierii sunt comune.
Controlul la nivelul transcrierii prin oprirea procesului nu are semnificatia blocarii sale complete (off state). Nu se cunoaste nici un sistem cu intrerupere totala a sintezei ARNm. Totdeauna ramane un nivel foarte scazut al transcrierii, ce consta din l-2 evenimente de transcriere/generatie celulara, astfel incat sinteza produsului genic este foarte limitata. Numai in sporii bacterieni, exprimarea genelor este total oprita.
Mecanismele de reglare la nivelul transcrierii sunt dependente de rolul enzimelor reglate: daca actioneaza in caile degradative sau au rol in reactiile de anabolism. De exemplu, intr-un sistem degradativ cu mai multe trepte, disponibilitatea moleculei ce urmeaza a fi degradata, conditioneaza adeseori, sinteza enzimelor. Invers, intr-o cale biosintetica, produsul final al caii, este adeseori, molecula reglatoare.
Chiar intr-un sistem in care o singura molecula proteica este sintetizata din ARNm monocistronic, proteina poate fi autoreglata, adica ea insasi poate sa inhibe initierea transcrierii. Concentratia crescuta a proteinei, scade rata transcrierii ARNm specific (fig 135).
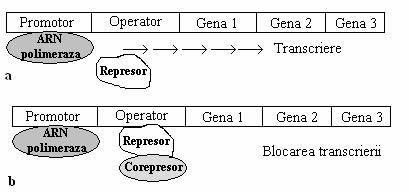
Fig. 135. Mecanismul represiei sintezei enzimelor. a. Represorul este inactiv, adica nu se poate asocia cu secventa operator, iar genele operonului sunt transcrise in ARNm. b. Dupa legarea corepresorului (o molecula mica exogena) cu represorul, acesta se activeaza, se asociaza la regiunea operatoare si transcrierea genelor operonului este blocata.
Mecanismele moleculare sunt proprii pentru fiecare sistem reglator, dar in functie de efectele asupra celulei, procesele reglatoare sunt de doua categorii:
Procese reglatoare pozitive sunt acelea in care exprimarea informatiei genetice creste cantitativ. Intr-un sistem reglat pozitiv, o molecula efectoare (o proteina, un complex molecular) activeaza rata transcrierii de la un promotor.
Procesele reglatoare negative semnifica diminuarea cantitativa a exprimarii informatiei genetice. Intr-un sistem reglat negativ, transcrierea genelor este blocata de un inhibitor prezent in celula. Inhibitorul se numeste represor. Pentru reluarea transcrierii, este necesar un antagonist al represorului denumit inductor.
O cale degradativa poate fi reglata pozitiv sau negativ.
Intr-o cale biosintetica, produsul final al caii, regleaza negativ propria sa sinteza.
Activitatea reglatoare a diferitelor activitati celulare se realizeaza la nivelul unitatilor genetice denumite operoni.
Modelul reglator al operonului
Operonul este o unitate functionala de reglare, alcatuita dintr-un grup de cistroni (A, B, C etc.), care codifica o anumita functie (de exemplu, biosinteza enzimelor unei cai metabolice) si este transcrisa unitar intr-o molecula de ARNm policistronic. Cel mai cunoscut este operonul lac, care codifica si regleaza sinteza enzimelor ce intervin in metabolismul lactozei.
Pe baza studiului operonului lac, Jacob si Monod (l967) au elaborat modelul clasic de structura a operonilor. Un operon este alcatuit din:
gene structurale;
regiunea operator (o) formata din circa 27 de perechi de baze, ce functioneaza ca situs de recunoastere si legare a represorului specific;
regiunea promotor (p), cu o lungime de circa 80 de baze, cu rolul de a lega ARN-polimeraza pentru initierea transcrierii genelor structurale;
gena reglatoare, situata adiacent sau la distanta de regiunea operatoare, ce tine sub control toate celelate gene ale unui operon.
La bacterii, activitatea genica este reglata, in primul rand, la nivelul transcrierii. Transcrierea coordonata a genelor intr-un operon este blocata de proteinele represor si este activata de proteinele stimulatoare.
Elementele genetice ale operonului bacterian sunt urmatoarele: gena reglatoare (i), situsul operator si un set de gene structurale (z, y, a) (fig. 136).
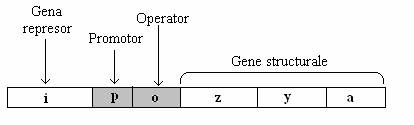
Fig. 136. Harta genetica a operonului lac. Secventele p si o sunt mult mai mici decat genele.
Gena reglatoare (i) codifica sinteza unui represor de natura proteica, ce interactioneaza cu situsul operator. Situsul operator este un segment de ADN adiacent genei structurale pe care o controleaza. Operonul lac poate fi reglat pozitiv (inductie) sau negativ (represie).
Reglarea pozitiva (Inductia sintezei enzimelor)
Majoritatea genelor din structura cromosomului bacterian sunt nefunctionale in absenta substratului specific, astfel ca enzimele de catabolism nu se sintetizeaza. Dupa adaugarea substratului in mediu, gena (genele) se activeaza, fenomen cunoscut sub denumirea de inductie sau derepresie.
Enzimele care se sintetizeaza numai in prezenta substratului corespunzator (adaugat in mediul de crestere) se numesc enzime inductibile sau adaptative (Karstrom, l938). Enzimele constitutive sunt acelea a caror sinteza este permanenta, independenta de prezenta substratului in mediu.
Numeroase enzime de catabolism apartin categoriei enzimelor inductibile. Cei mai cunoscuti inductori sunt amidonul (pentru amilaza), zaharoza (pentru invertaza), lactoza (pentru b-galactozidaza).
Mecanismul molecular al inductiei enzimatice. E. coli utilizeaza lactoza ca unica sursa de carbon. Enzimele esentiale pentru catabolismul acestui dizaharid sunt β-galactozidaza (sub actiunea careia, lactoza este hidrolizata la galactoza si glucoza), galactozid-permeaza (necesara transportului lactozei prin membrana celulei bacteriene) si thiogalactozid-transacetilaza (al carei rol fiziologic este incert).
Celulele de E. coli cultivate pe un mediu cu glucoza sau cu glicerol, contin mai putin de l0 molecule de b-galactozidaza/celula, iar numarul lor creste la cateva mii, dupa cultivarea celulelor pe mediul cu lactoza. Sinteza b-galactozidazei este inductibila.
Inductorul fiziologic al sintezei b-galactozidazei este alolactoza (izomerul glucoza-l,6-galactoza), derivata din lactoza, printr-o reactie de transglicozilare. Sinteza alolactozei este catalizata de cele cateva molecule de b-galactozidaza, existente in celula, inainte de inductie. Sinteza b-galactozidazei la nivel minim se numeste sinteza constitutiva de baza (bazala) si este posibila, deoarece legarea represorului de operator nu este niciodata atat de ferma incat sa blocheze complet transcrierea operonului.
Inductorul (lactoza) se asociaza cu represorul si il modifica, astfel incat acesta nu mai interactioneaza cu operatorul. In prezenta inductorului, operatorul ramane liber si promotorul este disponibil pentru initierea sintezei ARNm. Genele structurale z, y, a sunt transcrise intr-o molecula de ARNm, codificatoare pentru toate cele trei proteine. O molecula de ARNm ce codifica sinteza a cel putin doua proteine diferite se numeste poligenomica sau policistronica (fig 137).
Reglarea operonului necesita ca gena operatoare sa fie adiacenta genelor structurale ale operonului (lac z, lac y, lac a), dar proximitatea genei lac i nu este obligatoriu necesara, deoarece represorul lac I este o proteina difuzibila.
Genele ARNm lac sunt traduse diferential. Raportul cantitativ al celor trei proteine (b-galactozidaza, permeaza si transacetilaza) este l,0 : 0,5 : 0,2. Sinteza lor diferentiata cantitativ este rezultatul reglarii traducerii pe doua cai:
l) Gena z este tradusa prima, din ARNm policistronic. Dupa sinteza b-galactozidazei, jumatate din ribosomi se detaseaza de molecula de ARNm. Cealalta jumatate a numarului de ribosomi continua traducerea si la nivelul genei y. Astfel, numarul moleculelor de permeaza va fi numai jumatate din numarul moleculelor de beta-galactozidaza;
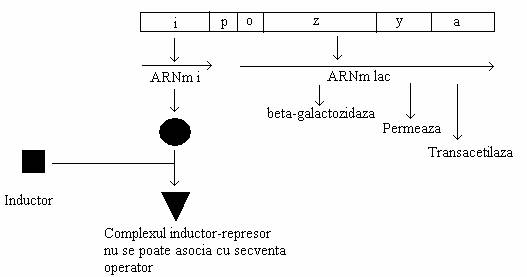
Fig. 137. Diagrama operonului lac in stare activa (indusa). Un inductor exogen se asociaza cu represorul si formeaza un complex inactiv, care nu recunoaste secventa operator. Genele operonului sunt transcrise si se sintetizeza enzimele pentru catabolismul lactozei.
2) ARNm bacterian este degradat dupa cateva cicluri de traducere. Degradarea este lenta, dar incepe cu o frecventa mai mare la nivelul genei a decat la nivelul genei y, iar degradarea la nivelul genei y este mai frecventa decat la nivelul genei z.
Reglarea negativa (Inductia sintezei enzimelor)
Proteina represor - produsul de sinteza a genei lac I, se leaga de secventa operatoare a operonului. Represorul s-a purificat din celulele bacteriene infectate cu fagi transductori ai genei lac. In absenta inductorului, represorul se leaga cu mare afinitate de regiunea operatoare (fig. 138). Represorul gaseste situsul operator prin difuzie unidimensionala de-a lungul moleculei de ADN si nu prin difuzie din mediul citoplasmatic. Intr-un mod asemanator isi gaseste situsul promotor, molecula de ARN-polimeraza. Represorul lac se leaga de situsul sau cromosomal, cu un mare grad de selectivitate.
S-au izolat mutante defective pentru sinteza uneia din cele trei proteine. De exemplu, mutanta z-y+a+ este defectiva pentru sinteza beta-galactozidazei, dar sintetizeaza permeaza si transacetilaza. Cea mai interesanta clasa de mutante este aceea a mutantelor constitutive, care sintetizeaza cantitati mari ale celor trei proteine in absenta inductorului.
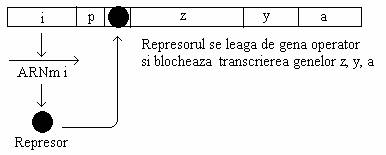
Fig. 138. Diagrama operonului lac in stare represata. Represorul activ se leaga de regiunea operator si blocheaza transcrierea genelor structurale.
Rata sintezei celor trei proteine este, in mod normal, controlata de o gena reglatoare comuna (gena i). Bacteriile de tip salbatic sunt inductibile si au genotip i+ z+ y+ a+. Efectul reglator al genei i se realizeaza prin intermediul represorului pe care-l codifica.
Represia sintezei enzimelor intr-un sistem biosintetic. Operonul triptofanului
Represia enzimatica reprezinta fenomenul scaderii ratei de sinteza a unui grup de enzime metabolic inrudite, ori chiar de oprire a sintezei lor, ca urmare a prezentei in celula, a unui metabolit cu rol de represor. Enzimele din aceasta categorie se numesc represibile.
Represia sintezei enzimelor s-a studiat la E. coli. Pe un mediu minimal, cu glucoza si azot combinat anorganic, celulele sintetizeaza toti monomerii necesari in proportii optime. In medii complexe (cu aminoacizi), prezenta produsului final al unei cai de biosinteza, reprezinta un semnal pentru celula, care, printr-un fenomen de feed-back negativ sau de represie prin produsul final al caii, stopeaza biosinteza enzimelor devenite inutile, intrucat celula foloseste cu precadere metabolitul existent ca atare in mediu. Invers, cand aprovizionarea cu triptofan diminua, se sintetizeaza enzimele sale de biosinteza.
Operonul triptofanului (trp) la E. coli codifica sinteza acestui aminoacid (fig. 139). Reglarea operonului se face astfel incat, cand triptofanul se gaseste in mediul de crestere, operonul trp nu este transcris. Cand aprovizionarea celulelor cu triptofan este insuficienta, transcrierea operonului este initiata.
Pentru reglarea unei cai biosintetice, o reglare de tipul on-off nu raspunde necesitatilor celulei. Este necesara o reglare cantitativa, deoarece in natura apar situatii in care, o cantitate suboptimala de triptofan este disponibila, dar aceasta este insuficienta pentru cresterea normala a celulei. Deficitul de triptofan in celula, este evitat printr-un sistem reglator modulator, in care, nivelul transcrierii operonului in starea derepresata este determinat de concentratia triptofanului disponibil. Acest mecanism reglator este activ pentru multi operoni care codifica biosinteza aminoacizilor.
Triptofanul se sintetizeaza in 5 trepte, fiecare catalizata de o enzima specifica. La E. coli, genele codificatoare (A, B, C, D, E) ale enzimelor sunt traduse dintr-o molecula de ARNm policistronic. Prima gena tradusa este trp E.
Gena pentru sinteza represorului (trp R) este localizata departe de acest policistron. Proteina codificata de gena trp R este aporepresorul trp, inactiv ca atare, datorita lipsei totale de afinitate pentru regiunea operator. In prezenta produsului final al caii de biosinteza (triptofanul), de origine exogena sau acumulat prin sinteza endogena, aporepresorul se activeaza: moleculele de triptofan, cu gr. mol. mica, se comporta ca un corepresor. Complexul format din aporepresor si corepresor se leaga de secventa operator si transcrierea genelor trp este blocata.
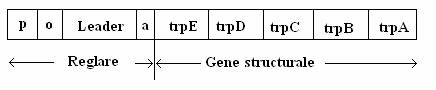
Fig. 139. Diagrama operonului triptofanului la E. coli.
Cand aprovizionarea celulei cu triptofan exogen este diminuata, operatorul este liber si transcrierea incepe. Acesta este principiul mecanismului reglator on-off.
Caile biosintetice sunt controlate insa, prin mecanisme mai fne, in care concentratia enzimelor unei cai este variabila in functie de concentratia produsului final al caii. Mecanismul consta in terminarea prematura a transcrierii, inainte ca prima gena structurala sa fie transcrisa.
Pe masura ce triptofanul disponibil scade, incepe transcrierea, dar aceasta este modulata cantitativ. Intre promotor si operator sunt doua regiuni, denumite leader si atenuator (trp L si trp a). Secventa L, necodificatoare, se gaseste la capatul 5' al moleculei de ARNm. La bacteriile de tip salbatic, in absenta triptofanului in mediu, transcrierea majoritatii moleculelor de ARNm se termina la secventa de 28 de baze, denumita atenuator. Rezultatul este transcrierea unui ARNm de l40 de nucleotide (al secventei L), fara ca genele structurale ale operonului sa fie transcrise. Secventa Leader codifica un polipeptid de l4 aminoacizi, care contine triptofan in pozitiile l0 si 11. Cei doi codoni ai triptofanului din secventa Leader sunt sensibili la concentratia ARNt trp, adica daca aprovizionarea cu triptofan este suficienta, traducerea ARNm Leader diminua.
Moleculele de ARNm Leader au rolul de a receptiona concentratia triptofanului sintetizat de enzimele operonului. Daca nivelul triptofanului este scazut, ARN-polimeraza depaseste situsul atenuator si transcrie intregul operon. Invers, cresterea concentratiei aminoacidului in celula, blocheaza transcrierea la nivelul secventei atenuator.
Alti cativa operoni care codifica sinteza aminoacizilor la E. coli, poseda situsuri atenuatoare si realizeaza o reglare cantitativa asemanatoare. Secventa atenuatoare este senzor al concentratiei celulare a aminoacidului a carui sinteza o regleaza. Peptidul leader al fiecarui operon contine cativa aminoacizi de tipul celor controlati de operon. De exemplu, operonul treoninei codifica enzime ce sintetizeaza treonina si izoleucina. Peptidul leader contine 8 resturi de treonina si 4 resturi de izoleucina. In secventa leader de l5 aminoacizi a fenil-alaninei se gasesc 7 resturi de fenil alanina, iar peptidul leader al operonului histidinei contine 7 resturi consecutive de histidina.
Din punct de vedere molecular, mecanismul reglator este conditionat de faptul ca la bacterii, transcrierea este cvasi-simultana cu traducerea mesajului.
Fenomenul represiei sintezei enzimelor prin feed-back, permite celulei sa mentina in limite adecvate, concentratiile relative ale enzimelor necesare sintezei aminoacizilor, proteinelor, acizilor nucleici etc.
Controlul activitatii enzimelor. Inhibitia prin produsul final
Reglarea prompta a activitatii enzimatice pentru evitarea supraproductiei de intermediari metabolici (aminoacizi, baze purinice si pirimidnice), se realizeaza prin inhibitia activitatii enzimelor, sub actiunea produsului final de biosinteza. Acest proces reglator se numeste inhibitie prin produs final, retroinhibitie sau inhibitie prin feed-back.
Daca celula ar controla numai biosinteza enzimelor, reglarea ar fi lipsita de promptitudine, deoarece enzimele deja existente in celula, si-ar exercita actiunea catalitica si rezultatul ar fi acumularea in exces a produsului caii respective. Celulele dispun de mecanisme de reglare fina, instantanee, ce constau in inhibarea sau blocarea functionarii primei enzime a caii biosintetice, sub actiunea produsului final aflat in exces.
Existenta reglarii prin inhibitia activitatii enzimelor s-a demonstrat pentru mai multe sisteme enzimatice, in care biosinteza unor metaboliti sau degradarea unui substrat se realizeaza printr-o serie de trepte succesive, fiecare fiind catalizata de o enzima diferita. Grupul de enzime care actioneaza cooperant pentru sinteza sau pentru degradarea unui metabolit se numeste zimom. Prezenta in mediu, a produsului final al ultimei trepte metabolice de sinteza, inhiba activitatea enzimei care catalizeaza prima treapta a biosintezei si astfel, intreaga serie de reactii succesive este blocata, dar se reactiveaza in absenta metabolitului.
E. coli sintetizeaza L-izoleucina din treonina. Daca mediul minimal, cu glucoza si NH4+ este suplimentat cu L-izoleucina, are loc blocarea prompta a activitatii enzimei L-treonin-deaminaza, implicata in prima treapta a biosintezei. Astfel, enzimele care catalizeaza reactiile urmatoare ale caii sintezei L-izoleucinei sunt lipsite de produsii intermediari, care reprezinta substratul lor specific.
De obicei este blocata numai prima treapta a unui lant metabolic.
Mecanismul molecular al inhibitiei prin produs final. Sitemele de retroinhibitie sunt active, in esenta, in reglarea cailor de biosinteza (anabolism). Produsul final al caii, inhiba actiunea catalazei ce catalizeaza prima treapta a sintezei produsului final. Produsul final al unei cai de biosinteza, formata din mai multe trepte, se deosebeste net de substratul primei enzime. Atat produsul final, cat si substratul sau nespecific, interactioneaza cu aceiasi enzima. Aceste enzime au caracter de proteine alosterice. Interactiunea cu substratul specific si cu produsul final al caii, are loc la situsuri moleculare diferite. Cand se combina cu produsul final al caii (efectorul alosteric), enzimele alosterice sunt incapabile sa functioneze catalitic, deoarece situsul lor activ sufera o tranzitie alosterica, la forma inactiva. Astfel de proteine ale caror configuratii spatiale se modifica dupa legarea moleculelor mici specifice, la alte situsuri decat situsul activ, se numesc proteine alosterice, iar moleculele efectoare ale acestor transformari se numesc efectori alosterici.
Exista mai multe tipuri de inhibitie prin produs final:
inhibitia cailor multifunctionale (multivalente), corespunde reglarii cailor metabolice ramificate, in care se sintetizeaza mai multi produsi finali. Pentru a asigura inhibitia reglatoare, este indispensabila prezenta diferitilor produsi finali. Actiunea izolata a fiecarui produs este ineficienta;
inhibitia cumulativa se caracterizeaza prin faptul ca, fiecare din produsele finale ale unei cai ramifcate, chiar la concentratia de saturare, inhiba partial activitatea primei enzime. Diferitii produsi finali actioneaza sinergic;
inhibitia asupra izoenzimelor: enzima ce catalizeaza prima treapta se gaseste sub mai multe forme (izoenzime), fiecare fiind sensibila la actiunea unui anumit produs. Cel mai cunoscut este cazul asparto-kinazei la E. coli, constituita din trei izoenzime.
Inhibitia prin produs final este de ordin cantitativ. Produsul final inhiba partial sau total, activitatea primei enzime, in functie de concentratia sa in mediu. Inhibitia totala a activitatii enzimei, se produce numai in cazul in care produsul final s-a acumulat in exces fata de necesitatile celulei.
Aplicatii ale represiei sintezei enzimelor in biotehnologie. Represia sintezei enzimelor ofera posibilitatea dirijarii proceselor de biosinteza industriala, pe urmatoarele cai:
limitarea prezentei si acumularii produsilor finali in mediu. Astfel, producerea glutamat-dehidrogenazei poate fi marita de circa 20 de ori la B. liqueniformis, prin utilizarea glucozei sau malatului ca sursa de C, in loc de glutamat sau de hidrolizat de cazeina;
limitarea producerii interne prin biosinteza, a produsilor finali, prin adaugarea in mediu, a unui inhibitor al caii metabolice;
"infometarea" partiala a unei mutante auxotrofe, fata de produsul necesar cresterii;
utilizarea unor mutante de reglare (mutante constitutive): fie a unor mutante care sintetizeaza un aporepresor ce nu se combina cu corepresorul, fie a unor mutante la nivelul regiunii operatoare, ce nu pot lega represorul. Ele sintetizeaza enzime constitutive, chiar in prezenta unor concentratii mari de produsi finali, care in mod normal sunt represoare.
Controlul sintezei proteice la nivelul traducerii
Genele codificatoare ale celor peste 50 de proteine ribosomale sunt localizate in peste 20 de operoni si sinteza lor este controlata riguros. Controlul este esential, deoarece componentele ribosomale reprezinta peste 40% din greutatea uscata a celulei bacteriene.
Controlul sintezei proteinelor ribosomale se exercita in primul rand la nivelul traducerii, spre deosebire de alte proteine care sunt reglate la nivelul transcrierii.
Cel putin o proteina, codificata de fiecare din cei peste 20 de operoni, are rolul de represor al traducerii. Represorul traducerii se leaga de ARNm langa situsul de initiere a propriei sale sinteze si blocheaza sinteza catorva proteine codificate de mesajul poligenic.
Legarea proteinelor reglatoare, de ARNm, nu interfera cu asamblarea ribosomilor, deoarece proteinele ribosomale se leaga mai ferm de ARNr decat de ARNm.
Represia traducerii are loc numai cand sinteza proteinelor ribosomale o devanseaza pe aceea a ARNr, sau cand aceste proteine nu se formeaza in cantitati echimolare.
Represia prin catabolit
Represia prin catabolit se exercita asupra sistemelor enzimatice inductibile ale catabolismului si a fost descrisa sub denumirea de "efectul glucozei", deoarece, la unele microorganisme, glucoza represeaza catabolizarea altor glucide. Acest tip de represie este evident in conditiile cultivarii unor bacterii, intr-un mediu ce contine un amestec de zaharuri, care sunt catabolizate succesiv, astfel incat curba de crestere este de tip diauxic.
Fenomenul de diauxie este datorat represiei catabolice pe care o exercita un compus usor si rapid metabolizabil (glucoza), asupra utilizarii unui alt glucid. Exemplul clasic al represiei prin catabolit, este al celulelor de E. coli, cultivate in prezenta unui amestec de glucoza-lactoza. Cultura are o curba de crestere diauxica. Daca un microorganism se cultiva in prezenta a doua substraturi glucidice, din care unul este represor, iar sistemul enzimatic de catabolizare a celui de al II-lea este sensibil la represie, metabolizarea primului glucid blocheaza sinteza enzimelor de metabolizare a celui de al II-lea. E. coli, cultivata pe un mediu cu glucoza si lactoza, creste initial, exclusiv pe seama catabolizarii glucozei, iar cand concentratia ei scade foarte mult, dupa o scurta perioada de lag, cresterea este reluata pe seama catabolizarii lactozei.
Glucoza este metabolizata cu o rata superioara altor glucide, astfel incat, in celula se acumuleaza o concentratie suficienta de metaboliti necesari diferitelor cai de sinteza.
Represia prin catabolit survine atunci cand un compus este metabolizat atat de rapid, incat produsii intermediari acopera intregul necesar de biosinteze ale celulei si face inutila si chiar costisitoare, functionarea concomitenta a caii de degradare a altui compus. Catabolitii intermediari in exces, represeaza nu numai enzimele caii metabolice prin care au fost generati, ci chiar si enzimele oricarei alte cai care are potentialitatea de a-i produce.
La nivel molecular, represia prin catabolit, este rezultatul scaderii concentratiei de AMP ciclic. Legarea ARN-polimerazei pentru transcrierea operonului lac, are loc numai daca o alta proteina, denumita proteina activatoare a catabolismului s-a asociat in imediata vecinatate a situsului promotor. Aceasta proteina se asociaza numai daca, in prealabil, s-a activat in prezenta AMP ciclic (fig 140).
In prezenta glucozei in mediu, nivelul AMP ciclic scade si proteina activatoare a catabolismului nu se asociaza promotorului operonului lac. In absenta acesteia, ARN-polimeraza nu initiaza transcrierea operonului. Astfel, represia prin catabolit pare a fi rezultatul unui deficit al AMP ciclic.

Fig. 140. Rolul AMP ciclic in sinteza ARNm si mecanismul represiei catabolice. Cand AMPc este disponbil, proteina activatoare a catabolismului (CAP) se asociaza de un situs adiacent promotorului, care la randul sau stimuleaza legarea ARN-polimerazei. in prezenta glucozei, AMPc este degradat, CAP nu se poate asocia si ARNm nu este transcris.
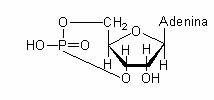
Structura moleculara a AMP (Adenozin-monofosfatul) ciclic.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2134
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved