| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
DOCUMENTE SIMILARE |
|||||||||||
|
|||||||||||
INFLUENTA FACTORILOR INTERNI ASUPRA CRESTERII
Cresterea plantelor este controlata de anumite substante endogene care fac parte din grupa hormonilor vegetali.
Hormonii sunt substante endogene, transportabile si active in cantitati foarte mici care favorizeaza desfasurarea proceselor metabolice. Ei au fost descoperiti pentru prima data la animale si denumiti ca atare de Starling (1906) (hormaein = a stimula). La plante, existenta hormonilor a fost presupusa inca din 1675 de catre savantul Malpighi, sub forma unei substante cu rol in reglarea cresterii pe care a denumit-o 'materia ad radices promovendas'.
Cercetarile
ulterioare efectuate de savantii Sachs si
Dupa modul de actiune asupra cresterii, hormonii vegetali pot fi stimulatori si inhibitori. Cele doua categorii de hormoni actioneaza competitiv in dirijarea cresterii, din balanta lor rezultand controlul acestui proces la plante.
8.4.1. Hormonii stimulatori de crestere
Hormonii stimulatori de crestere se clasifica in auxine, gibereline si citochinine.
Auxinele sunt compusi naturali, fiziologic polivalenti, care, in doze extrem de reduse, direct sau indirect, pot efectua atat cresterea cat si dezvoltarea plantelor, respectiv formarea organelor vegetative si generative.
Primele izolari ale auxinelor din tesuturile vegetale se datoresc cercetarile concomitente ale lui Went si Holodnii (1928) in coleoptilul de ovaz (Avena sativa). Went a demonstrat prezenta auxinei in varful coleoptilului, posibilitatea acesteia de a difuza pe agar-agar si rolul auxinei in curbarea coleoptilului spre sursa de lumina in miscarea de crestere numita fototropism. Sursa de auxina este varful coleoptilului, iar zona de reactie, care provoaca curbarea este subterminala.
In lucrarile sale Went a captat auxina din varfurile coleoptilelor de Avena sativa, prin trecerea ei in cuburi de agar (fig. 101). In cazul in care cuburile de agar cu auxina naturala au fost asezate pe suprafata unor coleoptile sectionate s-au produs curburi in cresterea plantelor in fuctie de pozitia lor de aranjare. Unghiul de curbura ar fi proportional cu concentratia substantei de crestere, exprimata in "unitati Avena", egale cu cantitatea de auxina care, aplicata pe coleoptilul decapitat de ovaz, produce o curbura de zece grade, la o temperatura de 220C si la o umiditate relativa a aerului de 92% (fig. 102).
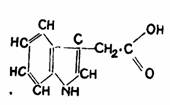
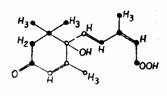
Prezenta in plante Cercetarile de extragere, purificare si identificare chimica au pus in evidenta prezenta auxinelor in diferite organe ale plantelor, in special cele cu crestere activa: varfuri ale tulpinii si radacinii, muguri, flori (polen, ovar), seminte in curs de germinare. Auxina naturala a fost identificata chimic cu acidul beta-indolilacetic, cu formula C10H9O2 notat prescurtat AIA (fig. 103).
|
|
Fig. 103 - Structura chimica a acidului beta-indolil acetic |
Biosinteza auxinelor porneste de la un
precursor care este aminoacidul triptofan. Acesta se
transforma succesiv in triptamina, acid beta-indolilpiruvic,
beta-indolilacetaldehida si acid beta-indolilacetic. Biosinteza este un proces metabolic catalizat de enzime, care are loc in
Din
Circulatia auxinelor in planta se face prin tesutul conducator liberian, pe baza diferentei de potential electric intre varful organelor, cu sarcina electrica - si baza organelor, cu sarcina electrica +. Auxinele fiind electronegative, circula in planta in sens bazipetal.
Auxinele pot migra si transversal, in cazul organelor asezate oriyontal (lastari) sau iluminate lateral (N. Dumitrescu, 1975), ceea ce explica geotropismul radacinilor si fototropismul tulpinilor.
Viteza de transport a auxinelor este de 4 mm/ora in radacini si 10-12mm/ora in tulpini.
Transportul polar al auxinelor in lastarul de mar a fost detectat prin folosirea izotopului radioactiv H3 in AIA (Kamboi si colab., 1997
Inactivarea auxinelor. Auxinele sunt degradate in plante sub actiunea luminii printr-un proces de oxidare, in care auxina activa este transformata in forma inactiva, numita lumi - auxin lactona.
Actiuni fiziologice La nivel celular, auxinele controleaza cresterea celulelor. In concentratii mai scazute, stimuleaza diviziunea, iar in concentratii mai ridicate extensia celulara.
In etapa diviziunii celulare auxinele stimuleaza sinteza acizilor nucleici, in special a ARN-m (mesager) cu rol in sinteza proteinelor. Acest efect a fost pus in evidenta in radacinile de sfecla de zahar (Duda si Cherri, 1971), in hipocotilul de soia (Holm si colab., 1970) si in semintele de fasole (Sacher, 1967).
Mecanismul de actiune al auxinelor in biosintezele organice cuprinde urmatoarele etape:
- in plasmalema auxina interactioneaza cu receptorul specific, iar complexul AIA - receptor activeaza functionarea pompei protonice, de H+ si stimuleaza procesele de absorbtie a elementelor minerale;
- in nucleoplasma auxina formeaza complexe proteice ce induc procesul de transcriptie, iar ARN-m patrunde in citoplasma unde activeaza biosinteza proteinelor;
- in citoplasma, auxina interactioneaza cu receptorii localizati in membranele reticulului endoplasmatic si activeaza biosinteza fosfolipidelor si altor compusi structurali (Polevoi, 1991
In etapa extensiei celulare auxinele stimuleaza cresterea in suprafata, elasticitatea, plasticitatea si permeabilitate membranei, precum si absorbtia apei si elementelor minerale
Mecanismul de actiune a auxinei asupra cresterii membranei sau peretelui celular este urmatorul: auxinele activeaza pompele protonice din plasmalema si transportul protonilor de la citoplasma la peretele celular. Transportul ionilor H+ favorizeaza absorbtia cationilor, in special K+ si Ca2+, dar si iesirea HCO3-. Ca rezultat are loc acidifierea peretilor celulari, fixarea Ca2+ si intensificarea activitatii enzimelor hidrolizante. Aceasta modifica componentii polimeri ai membranelor celulare si duce la slabirea legaturilor din peretii celulari. Ca urmare, sub actiunea fortei de turgescenta are loc cresterea prin extensie a acestora. Directia extensiei depinde de orientarea microfibrilelor de celuloza din membranele celulare (Duca, 1997).
Degradarea auxinei sub actiunea luminii controleaza raportul dintre etapele de extensie si diferentiere celulara.
Actiunea fiziologica a auxinelor este demonstrata de sporirea continutului acestora in timpul diferitelor procese de crestere din plante cum sunt germinarea semintelor, pornirea mugurilor, rizogeneza, morfogeneza tesutului conducator lemnos, cresterea in lungime si grosime a lastarului, formarea si cresterea fructelor.
Astfel, in timpul germinatiei semintelor de fasole, concentratia auxinelor creste in jurul embrionului, ceea ce determina atragerea substantelor cu rol plastic spre aceasta zona. La tomate, auxinele endogene determina hidroliza substantelor din endosperm, cresterea embrionului si a plantulei (Milica si colab., 1983). La vita de vie continutul de auxine din muguri este minim in timpul repausului (Chekol, 1995). Intreruperea repausului a produs o crestere a continutului de auxine in mugurii de pin, ceea ce permite reluarea capacitatii de crestere a acestora (Alden, 1971
Auxinele stimuleaza formarea si cresterea radacinilor. Rizogeneza butasilor de crizanteme (Tsukamoto si Matsubara, 1968) si a tubero-bulbilor de Freesia a fost asociata cu cresterea auxinelor in zonele de formare a radacinilor.
La
porumb, continutul de auxine din
Auxinele controleaza cresterea in lungime a lastarului la plantele lemnoase, continutul fiind maxim in timpul perioadei mari de crestere (Zimmermann si Brown, 1971), precum si cresterea in grosime, stimuland activitatea cambiului si morfogeneza tesutului conducator lemnos (Digby si Wareing, 1966
Absorbtia AIA in lastarii de mar testata cu H3, in H3AIA este maxima in timpul perioadei de crestere (Kambri si colab., 1997
Continutul de auxine creste in flori in timpul polenizarii, favorizand cresterea tubului polenic si interactiunea acestuia cu ovarul si ovulul in timpul fecundarii. Dupa fecundare, embrionul semintei reprezinta sursa de auxine care stimuleaza cresterea fructului (Milica si colab., 1983). Acest fenomen a fost pus in evidenta la porumb (Liu si Lee, 1995), la pomi si arbusti fructiferi. La vita de vie continutul de auxina in boabe este minim in faza de parga (Zhang si colab., 1997); in lipsa auxinelor in urma fecundarii se formeaza fructe fara seminte (Milica si colab., 1983
Giberelinele au fost descoperite de Kurosawa (1926) la plante de orez, bolnave de boala 'bakane' sau boala lastarilor lungi, produsa de ciuperca Gibberella fujikuroi. Din aceste plante Iabuta (1935) a izolat o substanta activa pe care a denumit-o giberelina. Ulterior au fost identificate prin metode moderne de separare si analiza (electroforeza, spectrometrie, cromatografie) 52 tipuri de gibereline, notate cu GA1-GA52.
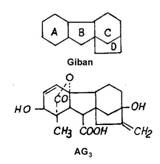
In structura chimica a giberelinelor exista un schelet comun numit gibban, la care se anexeaza anumite grupari laterale. Diferentele dintre cele 52 forme de gibereline constau in amplasarea gruparilor hidroxilice, metilice si carboxilice laterale, prezenta unei duble legaturi (la GA3, GA5, GA7,GA22) si a radicalului lactonic.
Prezenta in plante Giberelinele au fost identificate intr-un numar mare de specii de plante (100 dicotiledonate si 30 monocotiledonate), in diferite organe cum sunt varful tulpinii si radacinii, muguri, flori, seminte in curs de germinare, etc.
Biosinteza giberelinelor porneste de la un precursor numit acid mevalonic, care se transforma succesiv intr-un diterpenoid triciclic numit kauren, apoi kaurenol, acid kaurenic si acid giberelic (fig.104). Procesul de sinteza necesita prezenta ATP si a ionilor de Mg2+ si are loc in frunzele tinere, de unde giberelinele migreaza in varful radacinii si al tulpinii. In aceste zone giberelinele stimuleaza producerea si transportul descendent al auxinelor.
Circulatia giberelinelor in plante se face prin tesuturile conducatoare lemnos si liberian, cu o viteza de 5 cm/ora.
|
|
Fig. 104 - Structura chimica a acidului giberelic cu scheletul comun - giban |
Actiuni fiziologice La nivel celular mecanismul de actiune al giberelinelor consta in sporirea continutului de auxine, atat prin stimularea biosintezei, cat si prin franarea degradarii acestora. Astfel, giberelinele manifesta o actiune complementara auxinelor in etapa diviziunii, dar mai ales in etapa extensiei celulare.
Giberelinele stimuleaza biosinteza acizilor nucleici, in special ARN-m, cu rol in sinteza proteinelor. Cercetari recente demonstreaza ca la nivel celular giberelinele intensifica procesul de transcriptie.
Mecanismul de actiune al giberelinelor asupra inductiei genelor a fost pus in evidenta in timpul germinatiei semintelor de floarea soarelui. In timpul acestui proces giberelina trece din embrionul in crestere in endosperm, unde initiaza biosinteza enzimelor hidrolizante, alfa-amilazele, proteaza, fosfataza, beta-glucanaza, ribonucleaza. Enzimele hidrolizeaza rezervele din endosperm, iar produsii de hidroliza sunt utilizati la cresterea embrionului. Actiunea giberelinei determina mai intai modificari la nivelul reticulului endoplasmatic si al polizomilor ce determina sinteza alfa-amilazei, urmate de cresterea continutului de ARN.
Actiunea fiziologica a giberelinelor este demonstrata, ca si in cazul auxinelor, prin sporirea continutului endogen al acestora in timpul unor procese de crestere.
Astfel, continutul de gibereline creste in seminte in timpul germinatiei. In semintele germinate de grau au fost evidentiate giberelinele G1, G3, G8, G18 si G20 (Lenton si Appleford, 1991) si G92 (Penny si colab., 1994), care controleaza sinteza alfa-amilazei in embrion (Lenton si colab., 1994). La fasole, continutul de gibereline endogene creste in timpul germinatiei la 3 - 4 zile dupa imbibare, fiind mai ridicat in embrion si cotiledoane (Schirach - Szmiciel, 1979
La tuberculii de cartof, continutul de giberelina creste la intreruperea repausului mugurilor, fiind corelat cu ritmul de crestere al tulpinilor dupa plantare. La vita de vie continutul giberelinei endogene este minim in muguri in timpul repausului profund si creste spre sfarsitul starii de repaus (Chekol, 1994). Continutul de gibereline endogene creste si in mugurii de Salix viminalis si Betula pubescens, la intreruperea starii de repaus (Netien 1963, Michniewicz si Kriesel, 1972).
La porumb,
giberelinele endogene determina alungirea tulpinilor si marirea continutului de
auxina endogena (Monteiro, 1992). In radacini au fost puse in evidenta G1 si
G3 (Poloni
si colab., 1993), giberelinele stimuland activitatea enzimei nitrat -
reductaza. La tomate, in timpul cresterii intense au fost puse in evidenta
numeroase tipuri de giberelina in
Mecanismul de actiune al giberelinelor consta in stimularea alungirii internodurilor. De asemenea, giberelinele stimuleaza activitatea cambiului si morfogeneza tesutului conducator lemnos (Shiminger, 1971
Luckwill (1970) arata ca la mar giberelina sintetizata in
La
lalele continutul de giberelina este corelat cu
intensitatea cresterii lastarului florifer (Rebers si
colab., 1995). In timpul infloririi,
cresterea continutului de gibereline in flori stimuleaza cresterea corolei, de
exemplu la Mirabilis
Acumularea giberelinei are loc mai intai in androceu, apoi in gineceu favorizand polenizarea si fecundarea. Dupa fecundare, giberelinele provenite din embrionul semintei stimuleaza etapele initiale ale cresterii fructului.
Giberelina nu stimuleaza rizogeneza datorita rolului scazut in stimularea diviziunii celulare.
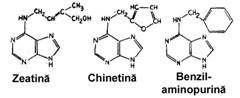
Citochininele. Spre deosebire de auxine si gibereline care au fost descoperite mai intai in plante pe baza efectelor de stimulare a cresterii si apoi identificate din punct de vedere chimic, citochininele au fost mai intai sintetizate pe cale chimica, ca substante cu rol in stimularea diviziunii celulare. Miller (1955) a obtinut un compus numit aminopurina care a stimulat diviziunea celulara in tesuturile calusului provenit din maduva tulpinii de tutun. Abia in 1963, Letham extrage si purifica citochinina naturala din boabele nemature de porumb zaharat, pe care o numeste zeatina. Ulterior au fost identificate la plante si alte citochinine, cum este izopentenil-adenina (fig. 105
|
|
Fig. 105 - Structura chimica a citochininelor naturale si sintetice |
Prezenta in planta Prezenta citochininelor a fost pusa in evidenta la o serie de plante in tesuturi cu diviziune celulara intensa, cum sunt semintele germinate, radacinile, lastarii, fructele tinere etc.
Biosinteza citochininelor se face pe baza degradarii acizilor nucleici, avand ca precursor acidul mevalonic. Sinteza are loc in radacini, de unde citochininele sunt transportate ascendent, odata cu seva bruta, prin tesutul conducator lemnos.
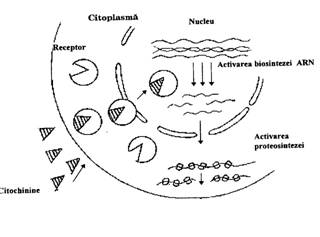
Actiuni fiziologice La nivel celular, actiunea fiziologica a citochininelor se manifesta prin stimularea diviziunii celulare. Citochininele stimuleaza biosinteza acizilor nucleici, in special a ARN-m, cu rol in sinteza proteinelor, stimuleaza biosinteza ARN-polimerazelor si intensifica activitatea functionala a nucleului. In citoplasma, citochininele determina intensificarea dezvoltarii reticulului endoplasmatic, sporirea numarului de ribozomi in special al polizomilor si intensifica translatia (fig. 106). In cloroplaste se formeaza noi grane si lamele, iar in mitocondrii creste numarul crestelor mitocondriale. Prin acest efect, citochininele intarzie imbatranirea si provoaca intinerirea tesuturilor.
|
|
|
Fig. 106 - Schema de actiune a citochininelor asupra metabolismului
celular in |
In planta rolul citochininelor este relevat de
continutul acestora in timpul diferitelor procese fiziologice. Astfel, continutul
de citochinine este ridicat in semintele in curs de
germinare. La mugurii de Populus x
robusta, continutul citochininelor este foarte scazut in timpul repausului,
dar creste la pornirea acestora (Hwet
si Wareing, 1973), iar la mugurii de
8.4.2. Hormonii inhibitori de crestere
Acidul abscisic este considerat principalul hormon cu actiune inhibitoare asupra cresterii plantelor. In modul sau de actiune, acidul abscisic manifesta o activitate competitiva stimulatorilor de crestere, auxine, gibereline si citochinine.
In anul 1963 Addicott a descoperit o substanta care accelereaza caderea sau abscisia fructelor de bumbac, pe care a denumit-o abscisina II. In 1964, Wareing a descoperit o substanta inhibitoare care induce repausul sau dormanta mugurilor, pe care a denumit-o dormina. Prin determinarea compozitiei chimice, aceste substante s-au dovedit identice, iar in anul 1967 au primit denumirea de acid abscisic (fig. 107).
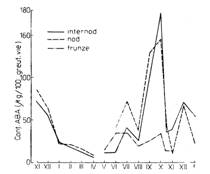
Prezenta in plante Acidul abscisic a fost identificat in organele vegetale la diferite specii, in legatura cu diferite procese fiziologice. Astfel, continutul de acid abscisic este ridicat in seminte si muguri in timpul starii de repaus. Acidul abscisic se acumuleaza in plantele anuale in perioada de imbatranire sau senescenta si determina oprirea cresterii lastarului la plantele perene. De asemenea, produce caderea frunzelor si stimuleaza coacerea fructelor. Cercetari recente demonstreaza participarea acidului abscisic in mecanismele de rezistenta a plantelor la conditiile de stress (fig. 108
Biosinteza acidului abscisic are loc in
|
|
|
|
Fig. 107 - Structura chimica a acidului abscisic ( |
Fig. 108. - Ciclul anual al continutului de acid abscisic in
lastari si |
Circulatia se face prin tesutul conducator liberian, fiind mai intensa in sens descendent decat ascendent, dar uneori si prin tesutul conducator lemnos (Davison si Young, 1973
Actiunea fiziologica La nivel celular acidul abscisic manifesta o actiune competitiva stimulatorilor. El inhiba diviziunea si extensia celulelor, dar favorizeaza diferentierea celulara. Acidul abscisic inhiba sinteza auxinelor, giberelinelor si citochininelor si a acizilor nucleici, in special a ARN-m, cu rol in sinteza proteinelor.
Acidul abscisic a manifestat o actiune competitiva cu auxinele si giberelinele. Actiunea competitiva cu giberelinele s-a manifestat in timpul repausului mugurilor (Shin si Rappaport, 1970), iar cu auxinele in timpul inradacinarii butasilor (Basu si colab., 1970). Acidul abscisic se acumuleaza in semintele de grau in repaus si scade in timpul germinatiei (Lenton si Appleford, 1991). De asemenea, se acumuleaza in mugurii in repaus de piersic (Corgan si Peyton, 1970), mar (Seelei si Powell, 1971) si vita de vie (Koussa si colab., 1994). Acidul abscisic favorizeaza coacerea fructelor de tomate (Drfling, 1970), mar (Rudnicki si Pieniazec, 1970) si lamai (Goldschmidt si colab., 1971
Acidul abscisic controleaza rezistenta la seceta a cerealelor. La grau continutul endogen creste pe soluri uscate (Rugby si colab., 1994), iar la porumb mentine potentialul de crestere al radacinii la stress hidric (Sharp si colab., 1992). La grau si porumb, acidul abscisic endogen mareste rezistenta la seceta prin reglarea conductantei stomatale (Quarrie si colab., 1994). La grau, acidul abscisic induce acumularea 4-aminobutiratului, mecanism implicat in rezistenta la stress (Reggiani si colab., 1993), iar la porumb controleaza biosinteza ARN-m si a proteinelor specifice rezistentei la seceta (Pages si colab., 1993), precum si acumularea prolinei (Ohier si Sharp, 1994
Acidul abscisic endogen induce rezistenta la inghet la plantele de grau favorizand acumularea de zaharuri, proteine solubile si substanta uscata (Lepnia si colab., 1994) si protejeaza plantulele de porumb de degradarea provocata de racire (Prasad si colab., 1994
La grau, acumularea acidului abscisic a redus efectele negative ale salinitatii solului asupra producerii de substanta uscata, a intensitatii transpiratiei si a continutului de clorofila a si b (Malibari, 1993
La pomii fructiferi continutul de acid abscisic endogen controleaza rezistenta la seceta, la piersic prin micsorarea conductantei stomatale (Caneia si colab., 1992), iar la mar prin reducerea fotosintezei si transpiratiei (Fernandez si colab., 1997
La vita de vie fotosinteza si transpiratia sunt reduse de catre acidul abscisic endogen prin micsorarea conductantei stomatale (Diring si colab., 1997
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5036
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved