| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Lipoliza
Lipidele influenteaza formarea aromei si texturii deoarece:
reprezinta o sursa de acizi grasi, in special cu catena scurta, care contribuie semnificativ la formarea aromei. Acizii grasi rezulta prin lipoliza, sub actiunea lipazelor. In unele tipuri de branzeturi, acizii grasi pot fi transformati in alte componente cu importanta pentru gust si aroma, in special metil cetone si lactone.
acizii grasi polinesaturati sufera oxidari care conduc la formarea diferitelor aldehide nesaturate, puternic aromate, determinand aparitia rancezirii oxidative. Oxidarea lipidelor in branzeturi pare a fi foarte limitata, probabil datorita potentialului redox scazut (-250 mV).
lipidele functioneaza ca solventi pentru componentele de gust si aroma, obtinute din lipide, proteine si lactoza. Lipidele pot absorbi din mediul inconjurator compusi care produc defecte de gust si miros.
Contributiile lipidelor la aroma branzeturilor sunt prezentate in fig.
LIPOLIZA: actiunea lipazelor
![]()
![]() Trigliceride 1,2 sau 2,3 digliceride 2 monogliceride
Trigliceride 1,2 sau 2,3 digliceride 2 monogliceride
REESTERIFICARE: actiunea esterazelor
![]() acid
gras + alcool ester
acid
gras + alcool ester
DEGRADARE OXIDATIVA:
- 2H + 2H2O
- 2H
![]()
![]()
![]() Acid gras saturat acid gras nesaturat acid β hidroxilat
Acid gras saturat acid gras nesaturat acid β hidroxilat
H2O
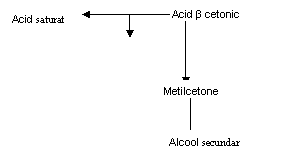
CO2
CO2
Fig. 10 Degaradarea grasimii in maturarea branzeturilor
Fractiunea grasa din branza este imporanta pentru formarea si perceperea aromei. Branza Cheddar obtinuta din lapte degresat nu are aroma caracteristica nici dupa 12 luni de depozitare (Ohren si Tuckey, 1967). Dezvoltarea redusa a aromei si texturii in branzeturile cu un continut redus de grasime reprezinta probleme tehnologice considerabile, care limiteaza comercializarea/desfacerea acestor produse.
Inlocuirea grasimii laptelui cu uleiuri vegetale sau minerale imbunatateste aroma perceputa in branza Cheddar fata de branza obtinuta din lapte degresat, ceea ce sugereaza ca un rol important al grasimii este de solvent a compusilor de aroma. Foda, s.a. (1974) au ajuns la concluzia ca emulsia naturala a grasimii laptelui a dus la obtinerea unor sortimente de branza mai bune decat grasimea laptelui emulsifiata in lapte degresat, sugerand ca interfata apa/grasime joaca un rol important in dezvoltarea aromei branzeturilor. Acesti autori considera ca grasimea naturala da o aroma superioara comparativ cu uleiurile vegetale sau minerale, deci compozitia acizilor grasi din grasimea laptelui este importanta pentru formarea aromei.
Ca in toate alimentale cu un mare continut de grasime, grasimea din lapte poate suferi degradari via lipoliza (enzimatice) sau oxidative ( chimice). Gradul de oxidare al lipidelor in branza este limitat (Downey,1969), probabil datorita potentialului redox scazut al branzei si prezentei antioxidantilor naturali.
Acizii grasi eliberati prin lipoliza contribuie in mod direct la formarea aromei, in special in branzeturile italiene si in sortimetele cu mucegai si prezinta o mai mica importanta in branza Cheddar si de Olanda, in special atunci cand branza foarte matura are un continut echilibrat in produsi de maturare.
Lipoliza extensiva este nedorita pentru multe sortimente: Cheddar, Gouda si branza Elvetiana, unde chiar un continut mediu de acizi grasi liberi poate fi privit drept rancezire. Bills si Day (1964) au gasit o serie de diferente minore intre concentratiile acizilor grasi liberi in branza Cheddar cu arome diferite. Proportia AGL (acizi grasi liberi) C6:0-C18:3 este similara celei din laptele gras, indicand faptul ca AGL au fost eliberati nespecific. Oricum acidul butiric liber a fost gasit in concentratii mai mari decat s-au putut explica prin proportia sa in grasimea laptelui, sugerand faptul ca a fost eliberat selectiv sau sintetizat de microflora branzeturilor.
Concentratia totala a AGL in diferite sortimete de branza este prezentata in tabelul 9
Tabelul 9. Concentratia AGL din branzeturi
|
Sortiment |
AGL, mg/kg |
Sortiment |
AGL, mg/kg |
|
|
Edam |
Gruyere | |||
|
Mozzarella |
Provolone | |||
|
Colby |
Brick | |||
|
Camembert |
Limburger | |||
|
Port Salud |
Parmesan | |||
|
Cheddar |
Romano | |||
|
Roquefort | |||
Bills si Day (1964) au subliniat faptul ca pH-ul branzei are o mare importanta asupra aromei AGL. La pH-ul branzei Cheddar (5,2) o cantitate importanta de AGL este prezenta sub forma de saruri, reducandu-se astfel aroma. Lipoliza in sortimentele italiene este extensiva si datorita actiunii preesterazelor gastrice (PEG) din cheagul utilizat la fabricarea acestor branzeturi. In sortimentele cu mucegai albastru lipoliza este extensiva datorita lipazelor sintetizate de speciile de Penicillium .
AGL reprezinta un substrat pentru formarea si a altor compusi de aroma. Catabolismul lor la metil-cetone in sortimentele maturate cu mucegai este foarte important, metilcetonele din lanturile intermediare fiind principalii compusi de aroma in branzeturile cu mucegai albastru.
Wang (1973-1975) a considerat ca lactonele din acizii grasi pot contribui semnificativ la formarea aromei branzei Cheddar daca sunt prezente in exces fata de valoarea preconizata. Alti produsi ai lipolizei care influenteaza aroma sunt esterii si amidele.
Lipazele din branzeturi
Lipazele provin in branza din lapte, renina, cultura starter, BLS, BLNS. Lipazele sunt hidrolaze care scindeaza esterii acizilor carboxilici (EC 3.1.1.)dupa reactia:
![]() R1-C-OR2
+ H2O R1-C-OH
+ R2OH
R1-C-OR2
+ H2O R1-C-OH
+ R2OH
In general in sortimentele in care lipoliza este extensiva, lipazele provin din enzima coagulanta (pasta de renina pentru sortimentele italiene), sau din culturile starter secundare (pentru sortimentele cu mucegai).
Lipazele prezinta diferite tipuri de specificitate:
sunt in mod specifice pentru legaturile esterice exterioare ale tri- sau digliceridelor (pozitia sn-1 si sn-3). Initial hidrolizeaza trigliceridele la 1,2 si 2,3 digliceride si apoi la 2-monogliceride. Acidul gras din pozitia sn-2 migreaza in pozitia vacanta sn-1 sau sn-3 si este apoi eliberat prin lipoliza. Deci lipazele hidrolizeaza trigliceridele in glicerol si 3 acizi grasi. In multe branzeturi lipoliza nu depaseste prima etapa.
Lipazele prezinta in mod obisnuit specificitate pentru o anumita lungime a catenei.
Unele lipaze prezinta specificitate pentru acizii grasi saturati sau nesaturati.
Lipazele sunt inactivate de Ca2+, care precipita acizii grasi ca sapunuri insolubile. Proteinele zerului(β-LG si serumalbumina) leaga acizii grasi si stimuleaza activitatea lipazelor. Lipazele sunt activate de sarurile biliare care emulsioneaza substratul reprezentat de trigliceride. Laptele contine o lipoproteinlipaza (LPL) si o serie de esteraze. Rolul fiziologic al LPL este implicarea in metabolismul trigliceridelor din plasma. LPL bovina este mai curand nespecifica si elibereaza usor acizii grasi din pozitiile 1 si 3 ale mono-, di- si trigliceridelor si din pozitia 1 a glicerofosfolipidelor. LPL din lapte elibereaza preferential acizii cu lanturi scurte si medii deoarece in trigliceridele din lapte acizii cu lanturi scurte sunt esterificati predonimant in pozitia sn-3 astfel incat sa fie mai accesibila hidroliza, sau datorita lanturilor lungi de acizi care sunt mai susceptibili la reincorporarea in trigliceride, reactie catalitata de LPL sintetaza. Specificitatea aparenta este determinata de solubilitatea si mobilitatea trigliceridelor care contribuie mai mult decat specificitatea de substrat a lipazei.
In laptele bovinelor mai mult de 80% din LPL este asociata cu micelele de cazeina si probabil este incorporata in branza. LPL determina o lipoliza avansata in laptele crud destinat fabricarii branzeturilor si poate contribui deasemenea la lipoliza laptelui pasteurizat pentru branzeturi din moment ce incalizirea la tempretauri mai mari de 780C timp de 10 s este necesara pentru inactivarea completa a enzimei (Driessen, 1989). Un extract de renina de calitate superioara nu prezinta activitate lipolitica, dar pasta de renina utilizata la fabricarea branzeturilor (Romano, Provolone) contine o lipaza denumita esteraza pregastrica (PGE), denumita si care catalizeaza intens procesul de proteoliza si este responsabila pentru aroma picanta a acestor sortimente. PGE denumita si lipaza orala, sau linguala a fost prezentata de Nelson (1977). Ea este secretata de glandele de la baza limbii, care sunt stimulate prin alaptare si este ulterior spalata in abomasum odata cu latele si saliva. Pasta de renina este preparata din abomasumul viteilor sau mieilor imediat dupa intarcare. Abomasumul este uscat, macinat sub forma de pasta, care este suspensionata in lapte sau apa inainte de a fi adaugata in branzeturi. Extractele care contin PGE pot fi obtinute din stomacul viteilor, mieilor, iezilor si au fost partial purificate din preparatele comerciale (Lec, 1980) putand fi izolate si din tesutul bucal. Enzima este o glicoproteina cu pH optim 7 si masa moleculara 49 kda.
PGE este puternic specifica fata de lanturile scurte de acizi esterificati in pozitia sn-3. Din moment ce laturile scurte si medii ale acizilor grasi din trigliceridele laptelui se situeaza predominant in pozitia sn-3, PGE elibereaza cantitati mari prin hidroliza. Diferentele usoare in ceea ce priveste specificitatea PGE provenita de la miei, iezi si vitei conduc la obtinerea branzeturilor italiene cu arome diferite, caracteristice.
Majoritatea celorlalte lipaze sunt necorespunzatoare pentru obtinerea branzeturilor de tip Italian datorita specificitatii nedorite, dar s-a observat ca anumite lipaze fungice pot fi utilizate ca alternative acceptabile (Fox, 1988, 1993).
Prezenta unei slabe activitati lipolitice data de speciile de Lactococcus a fost stabilita, iar in absenta unei microflore puternic lipolitice de suprafata aceasta actioneza, contribuind semnificativ la nivelul relativ scazut al lipolizei pentru sortimentele Cheddar si Olanda.
Fryer, s.a. (1967) a considerat ca desi prezinta o activitate lipolitica redusa, lactococii vor hidroliza grasimea din lapte intr-o proportie mare, daca vor fi prezenti in numar mare pe o periaoda lunga de timp (de ex. in timpul maturarii branzeturilor). Bucatile mici de branza aseptica acidifiate cu acid δ lactonogluconic inloc de culturi starter, contin cantitati reduse de AGL (Reiter, 1967). Umemoto (1968) a observat ca extractele purificate de bacterii lactice au fost mult mai active asupra tributiratului decat asupra tripalmitului sau trioleinei. Harper s.a. (1980) au raportat ca Lactococcus lactis ssp. lactis prezinta un sistem esterazic mai complex decat Lactococcus lactis ssp. cremoris
Au fost evidentiate diferente pentru tulpini lactice in ceea ce priveste capacitatea de sinteza a lipazelor si esterazelor, influentata de compozitia mediului de crestere si de varsta culturii ( Lactococii au activitate lipolitica mai puternica decat lactobacilii). Specificitatea lipazei lactococice poate fi modificata prin iradiere UV (Singh, 1981). Lipazele in extractele purificate a unui numar de tulpini de Lactococcus au fost mult mai active la 37O C, si la pH 7-8,5 si au fost mult mai active asupra trigliceridelor care contin lanturi scurte de acizi grasi (C4:0-C10:0) decat asupra lanturilor lungi (C12:0-C18:1). Bacteriile starter pot hidroliza mono si digliceridele produse de alte lipaze (de ex. produse de LPL, sau lipazele provenite din bacterii G-).
Sitemul esteraza/lipaza al speciilor de Lactococcus este mai putin cunoscut comparativ cu activitatea lor proteolitica. Izolarea variantelor negative ale lipazelor/esterazelor din Lactococcus ar permite evaluarea importantei acestor enzime asupra maturarii branzeturilor. Oterhorm (1982) a izolat o esteraza din Lactobacillus plantarum si a constatat ca este activa la pH 6,7 si 40oC. Enzima nu a fost afectata de metalele grele sau de prezenta cianurilor si a prezentat o preferinta puternica pentru substraturile solubile sub forma de emulsie. Esterazele intracelulare produse de Lactobacillus helveticus, Lb. delbrueckii ssp. bulgaricus si Lb. delbrueckii ssp. lactis au fost studiate si au prezentat specificitate pentru lanturile scurte de acizi grasi (El Soda, 1986). Temperatura optima pentru producerea esterazei este 40-45oC. Celulele din Lactobacillus casei au avut o activitate esterazica si lipazica mai mare in faza logaritmica decat in faza stationara de crestere (Pitkiewicz, 1987).
Micrococcus si Pediococcus, care pot constitui o parte a microflorei nonstarter a branzeturilor produc de asemenea lipaze, contribuind la maturare. Lipaza produsa de Micrococcus freunderenchii a fost inhibata de organofosforate si ionii metalici divalenti, dar intr-o masura mult mai mica a fost inhibata de EDTA. Activitate esterolitica au prezentat extractele din 5 tulpini de Micrococcus, care au hidrolizat in principal p-nitrofenil derivatii din acizii grasi.
Esteraza din Micrococcus ssp. ATCC 8459 a prezentat activitate optima la pH 8 si 40oC, fiind inhibata de organofosfati, metale divalente, sare si reactii redox. Activitate esterazica si lipazica a fost gasita si intr-un numar de tulpini de Pediococcus pentosaceus, de origine lactica.
Proprionibacterium shermanii poseda o lipaza care contribuie la lipoliza in varietatile olandeze de branza. Activitatea optima a lipazei intracelulare din Proprionibacterium shermanii a fost stabilita la pH 4,7 si 47oC.
Lipazele si esterazele din Brevibacterium linens au fost studiate si s-a gasit o intensa activitate esterazica si lipazica (Foissy, 1974).
Lipoliza pentru sortimentele maturate cu mucegai se datoreaza in principal lipazelor din Penicillium roqueforti sau Penicillium camemberti care produc lipaze extracelulare foarte active. P. camemberti elibereaza numai o lipaza activa la pH 9 si 35oC, in timp ce P. roqueforti produce doua lipaze, una cu pH optim 7,5-8, iar cealalta 6-6,5, care prezinta specificitati diferite.
Bacteriile psihotrofe, care predomina in microflora laptelui refrigerat sunt o sursa foarte importanta de lipaze cu activitate puternica in branza, in special daca concentratia lor depaseste 107 ufc/ml. Multe lipaze psihotrofe sunt stabile termic si pot cauza rancezirea branzeturilor pastrate o perioada lunga de timp. Importanta enzimelor eliberate de bacteriile psihotrofe este mult mai mare decat a proteinazelor eliberate de bacteriile psihotrofe, deoarece proteinazele se pierd in proportie mare in zer, in timp ce lipazele sunt adsorbite pe suprafata globulelor de grasime si raman in branza.
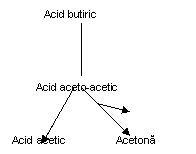
Catabolismul acizilor grasi liberi
Aroma branzeturilor cu mucegai albastru este dominata de prezenta 2-alcanone si cetone saturate (n-metil cetone), fiind prezente seriile omoloage ce contin atomi de carbon de la C3 la C15 in numar impar si unele lanturi pare. Concentratiile 2-alcanone din branza cu mucegai albastru variaza datorita reducerii acestora la 2-alcanoli (alcooli secundari). 2-heptanonele, 2-nonanonele si 2-undecanonele sunt metil-cetonele dominante in branza cu mucegai albastru in timpul maturarii (Dartley, 1971).
Metabolismul acizilor grasi in branzeturile cu Penicillium ssp. implica patru etape principale:
eliberarea acizilor grasi din lipaze;
oxidarea la β-cetoacizi;
decarboxilarea la 2-alcanone cu un atom de carbon mai putin;
reducerea 2-alcanonelor la 2-alcanolii corespunzatori (Hawke, 1966);
Etapa a patra este reversibila, in conditii de aerobioza. Concentratia 2-alcanonelor este in corespondenta cu lipoliza si acesti compusi pot fi formati prin actiunea mucegaiurilor asupra cetoacizilor prezenti in mod natural in concentratii mici in grasimea laptelui (cca. 1% din totalul acizilor grasi). 2-Alcanonele pot fi de asemenea formate prin oxidarea acizilor mononesaturati.
Un numar de factori influenteaza rata de producere a 2-alcanonelor in branzeturi (temperatura, pH, starea fiziologica a mucegaiurilor si concentratia acizilor grasi raportata la masa sporilor) (Adda, 1982).
Atat sporii rezistenti cat si miceliile sunt capabile de producerea 2-alcanonelor.
Lactonele sunt esteri ciclici, rezultati prin esterificarea intramoleculara a hidroacizilor, cu pierderea apei, pentru a forma o structura de inel. α- si β-lactonele sunt foarte reactive si instabile, aparand ca intermediare in sintezele organice. γ- si δ-lactonele sunt stabile si apar in branza. Lactonele au in general o aroma puternica, care desi nu este specifica branzeturilor poate avea un rol important in structura generala a aromei branzeturilor.
S-a stabilit ca γ- si δ-lactonele din laptele proaspat provin din hidroxiacizii corespunzatori, procesele oxidative din glanda mamara fiind o sursa principala de precursori ai lactonelor, care depinde de alimentatie, sezon, stadiul lactatiei, varsta si specie. Formarea γ- si δ-lactonele este spontana, urmand eliberarii hidroxiacizilor din trigliceride si de aceea formarea lactonelor trebuie corelata cu lipoliza. Wong (1975) a gasit concentratii mari de lactone in branza Cheddar si a sugerat ca acestea pot proveni prin reducerea cetoacizilor corespunzatori sau prin metabolismul microbian.
Stabilirea conditiilor de ultrafiltrare pentru obtinerea concentratului proteic din lapte
Analiza parametrilor de operare la ultrafiltrarea laptelui degresat
Separarea prin ultrafiltrare atat a cazeinei cat si a proteinelor serice cu indepartarea partiala a lactozei si a sarurilor minerale permite obtinerea unui concetrat proteic cu proteine practic nedenaturate. Concentratia finala a substantelor proteice, lactozei si substantelor minerale din concentrat depinde de gradul de concentrare realizat prin ultrafiltrare si de parametrii fizici principali ai procesului (temperatura, presiune).
O importanta aplicatie a tehnologiei ultrafiltrarii este realizarea concetratelor proteice care sa poata fi utilizate ca materie prima in industria branzeturilor, principalele avantaje fiind cresterea productivitatii si posibilitatea de standardiza compozitia materiei prime in vederea obtinerii unor loturi omogene din punct de vedere compozitional.
S-au realizat studii privind variatia parametrilor care influenteaza operatia de ultrafiltrare a laptelui degresat precum si variatia componentelor retentatului in cursul obtinerii concentratului proteic din lapte degresat.Concentrarea laptelui degresat a fost realizata pana la un factor de concentrare volumica FCV=VL/VR unde VL-volumul de lapte si VR-volumul retentatului.
Factorii care influenteaza operatia de ultrafiltrare a laptelui degresat
Experimentul in care s-a studiat evolutia fluxului de permeat in functie de factorul de concentrare (Fig. 11) s-a desfasurat la presiune de 1,4 at (I), 1,2 at (II), 0.1 at (III) la temperatura de lucru de 40 C.
Se poate observa faptul ca fluxul de permeat variaza invers proportional cu factorul de concentrare. La un factor de concentrare x2, valoarea fluxului de permeat scade cu 51% in comparatie cu fluxul initial. Pentru ultimele trei valori ale factorului de concentrare (41.8;1.9 si 2.0) fluxul de permeat s-a stabilizat in jurul valorii de 17 l/m2h (Proba I) si respectiv 15 l/m2h pentru probele II si III, datorita aparitiei fenomenului de colmatare care a determinat formarea unui strat de gel proteic de rezistenta constanta pe suprafata membranei.

Fig. 11. Variatia fluxului de permeat in functie de factorul de concentrare
Efectul presiunii si al temperaturii este prezentat in fig. 12. Experimetele au fost efectuate prin colectarea retentatului in vasul de alimentare. Se poate observa faptul ca asupra variatiei fluxului de permeat parametrul de proces temperatura are o influenta mai mare decat presiunea.
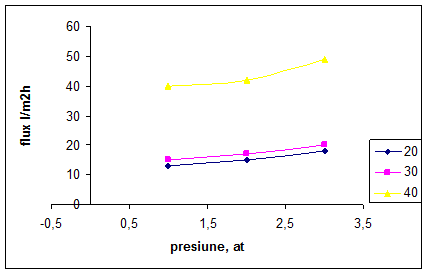
Fig. 12. Efectul presiunii si temperaturii asupra fluxului de permeat
Cresterea temperaturii de la 20 C la 40 C a condus la o marire a fluxului de permeat de 2,6 ori la presiune atmosferica, de 2,7 ori la p=2 at si respectiv de 2,8 ori la p=3 at.
Temperatura si presiunea variaza liniar cu debitul de permeat doar la valori scazute ale concentratiei de proteine.
Temperatura actioneaza favorabil asupra cresterii fluxului de permeat si datorita faptului ca la temperaturi ridicate se reduce vascozitatea fluidului de alimentare (lapte degresat), ceea ce micsoreaza necesarul de energie pentru pompa. De asemena, mentinerea unor valori marite ale temperaturii poate minimiza riscul aparitiei unor probleme legate de cresterea numarului de microorganisme in retentat. Tinand cont de aceste aspecte temperatura de lucru ar trebui mentinuta la valori cat mai ridicate posibil, fara a neglija influenta negativa asupra compozitiei fluidului de alimentare.
Analiza compozitionala a retentatului obtinut prin ultrafiltrarea laptelui degresat
Concentratul proteic din lapte degresat (CPL) a fost obtinut in conditii de statie pilot cu ajutorul instalatiei MAVIBRAN si a fost ultilizat ulterior ca materie prima pentru fabricarea branzeturilor.
In timpul procesului de ultrafiltrare au loc modificari ale retentiei componentelor laptelui degresat. Studiul modului in care componentele laptelui degresat se regasesc in retentat poate oferi informatii importante pentru unele etape ale fabricarii branzeturilor (folosind ca materie prima retentatul) cum ar fi coagularea si sinereza. S-au analizat 3 componente ale laptelui degresat (proteine, lactoza si substante minerale) si variatia continutului de proteine in cursul procesului de obtinere a CPL.
Pentru calculul componentelor retentatului s-a utilizat marimea denumita procent de recuperare (PR):
PR= (kg component in retentat/kg component in lapte)x100
Din anliza datelor prezentate in fig. 13 se poate constata ca s-a obtinut un procent de recuperare a proteinelor de 79.6% pentru factorul de concentrare utilizat. Cea mai importanta problema este legata de utilizarea retentatelor ultrafiltrate din lapte degresat la fabricarea branzeturilor este capacitatea tampon mai ridicata datorita cresterii continutului de substante minerale (in special calciul care ramane legat de cazeina).

Fig. 13. Procentul de recuperare al continutului de proteine in functie de factorul de concentrare (FC)
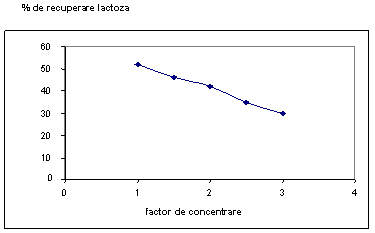
Fig. 14. Procentul de recuperare al continutului de lactoza in functie de factorul de concentrare
In tabelul 10 se prezinta compozitia laptelui degresat si a retentatelor obtinute pentru diferite valori ale factorului de concentrare. Prin reducerea continutului de lactoza se micsoreaza capacitatea de tamponare a CPL, care reprezinta principalul incovenient al utilizarii CPL la fabricarea branzeturilor.
Tabelul 10. Compozitia laptelui degresat si a retentatelor obtinute din lapte degresat
|
Compozitie % |
Factor de concentrare |
||||
|
X 1 |
X 1.5 |
X 2 |
X 2.5 |
X 3 |
|
|
Substanta uscata | |||||
|
Proteine | |||||
|
Lipide | |||||
|
Lactoza | |||||
|
Substante minerale | |||||
Analizand datele prezentate in fig. 13 si 14 se poate constata o reducere semnificativa a continutului de lactoza si saruri minerale dein retentat care poate conduce la obtinerea unui coagul care sa se formeze intr-un timp scurt, iar cantitatea de zer eliminata sa fie foarte mica.
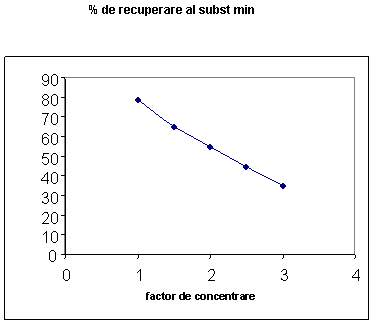
Fig. 15. Procentul de recuperare al continutului de substante minerale in functie de factorul de concentrare
Concluzii privind parametrii procesului de ultrafiltrare
In urma analizei datelor obtinute se pot face urmatoarele aprecieri:
Procesul de ultrafiltrare al laptelui degresat este influentat de factorul de concentrare, presiune si temperatura, valoarea fluxului de permeat scazand cu 48% la FCV x 2. Prin cresterea temperaturii cu 20 C se obtine o marire a fluxului de permeat de 2.71 ori la o presiune de 2 at.
Utilizand instalatia de MF/UF MAVIBRAN se poate realiza in conditii de statie pilot concentrarea proteinelor din lapte degresat cu o compozitie care sa ii permita utilizarea atat la fabricarea branzeturilor cat si la obtinerea concetratelor proteice uscate.
Calculul procentului de recuperare PR al componentelor retentatului (proteine, lactoza, substante minerale) permite analiza variatiei acestora in timpul procesului de UF si estimarea modificarilor ulterioare produse in timpul coagularii si maturarii.
In urma experimentelor s-au stabilit parametrii optimi de lucru pentru instalatia MAVIBRAN
Tabelul 11. Parametrii optimi de lucru stabiliti experimental
|
Parametru |
Factor de concentrare |
Presiune, at |
Temperatura, C |
PR proteine, % |
|
Valoare |
X 3 |
Metode experimetale pentru caracterizarea proceselor biochimice la maturarea branzeturilor
Determinarea continutului se substanta uscata
Analiza s-a facut dupa metoda standard, de referinta, prin evaporarea apei in etuva termoreglabila, urmata racirea la exicator si catarirea la masa constanta.
Aparatura: etuva termoreglabila, creuzet ceramic, balanta analitica, exicator cu capac, sticla de ceas, spatule.
Exprimare: umiditatea = (m2-m0/m1-m2)x100
unde: m1= masa capsulei cu produsul inainte de uscare, g;
m2= masa capsulei cu produsul dupa uscare, g;
m0 =masa capsulei goale, g.
Determinarea pH-ului
Determinarea s-a facut cu ajutorul aparatului pH meter 1100 CALIMATIC, portabil dotat cu senzor de temperatura PT 100 si cu electrod combinat de penetrare INGOLD care contine solutie saturata KCl si Ag/AgCl. Determinarile s-au facut la diferite temperaturi, deoarece aparatul s-a calibrat automat la fiecare analiza.
Determinarea aciditatii titrabile
S-a realizat prin titrarea a 10 g produs cu NaOH 0,1 n, in prezenta fenolftaleinei ca indicator. Exprimarea s-a facut in grade Thorner ( T).
Aparatura: pahar conic de 100 mL, pipeta cu bula de 10 mL, biureta gradata.
Exprimarea: aciditatea xV
Unde V = mL NaOH 0,1 n folositi la titrare.
Determinarea continutului de sare (NaCl) -Metoda Volhard
Metoda utilizata are ca principiu trecerea in suspensie apoasa a probei de analizat si acidularea acesteia cu acid azotic. Ionii de clor se titreaza potentiometric cu solutie de azotat de argint cu titru cunoscut.
Aparatura: balanta analitica, pH-metru prevazut cu electrod de referinta (electrod cu sulfat de mercur) si un electrod de masurare (electrod de argint), agitator mecanic, biureta, eprubete, pipete, cilindri gradati, fiola de cantarire, pahar Berzelius, baloane Erlenmeyer.
Reactivi: acid azotic, solutie 65%, clorura de potasiu, solutie saturata, azotat de argint, solutie 0,1 n. In cazul branzeturilor exprimarea rezultatelor se face dupa relatia:
NaCl% = 0,005841 x V1x100/m (g/100g).
Unde: V1=volumul solutiei 0,1 n de azotat de argint folosit la titrarea probei, mL
m= masa probei luate in analiza, g.
Determniarea continutului de grasime -metoda acid butirometrica (Gerber)
Determinarea continutului de grasime s-a efectuat folosind un butirometru pentru separarea grasimii prin centrifugare. S-au mai folosit alcool izoamilic si acid sulfuric concentrat. Sticlarie si materiale folosite: pipete 1 mL si pipete automate, baie termoreglabila, centrifuga tip Gerber cu 800-1200 rot/min, balanta analitica, folii de plastic. Reactivi : acid sulfuric d = 1,817, alcool izoamilic d = 0,810.
Determinarea continutului de lactoza
Determinarea lactozei s-a realizat cu ajutorul aparatului MILKOSCAN S50. Probele recoltate au fost mojarate foarte fin, diluate cu apa distilata, filtrate prin filtru sub vid. Filtratul astfel obtinut s-a analizat, rezultatul calculandu-se in functie de valoarea indicata de aparatul care a analizat proba si de cantiatea de branza luata in analiza precum si in functie de gradul de dilutie.
Metode de analiza a fractiunilor azotoase.
Metodologia de evaluare a gradului de maturare a tinut seama de sotimentul de branza analizat
Obiectivele studiului releva necesitatea aprecierii activitatii agentilor proteolitici in branzeturi
Profilul peptidelor fractiunii solubile la pH 4,6 (sau etanol-insolubile si a fractiunilor solubile formate) ar trebuie determinate prin cromatografie lichida cu inversare de faza (RP-HPLC). Prin urea-PAGE, CE si RP-HPLC pot fi efectuate si o serie de analize chimice/analitice ale profilului obtinut.
Metode chimice de evaluare a proteolizei la maturarea branzeturilor
Pentru a evalua proteolize in timpul maturarii, o schema de fractionare trebuie utilizata. Bondzynski a descris pana la un anumit nivel maturarea branzeturilor prin analiza continutului de N solubil in apa (WSN) si in acif fosfotungstic (PTA-SN) si a introdus expresii ca dimensiunea si profunzimea maturarii, pentru ceea ce reprezinta cele doua fractiuni separate. Aceste metode au fost dezvoltate in continuare si introducerea fractionarii la pH 4.4-4.6 (pH 4.4-SN) pentru a precipita toate cazeinele a facut posibila compararea rezultatelor obtinute din analiza mai multor tipuri de branzeturi cu un continut diferit de sare si valori diferite de pH;
Multiplele metode de fractionare utilizate determina diferite aspecte si sunt adaptate in moduri diferite la varietatea sortimentala deosebit de bogata a branzeturilor din lumea intreaga. Standardizarea va intampina probleme dificile si ar putea sa nu fie utila.In Europa s-a incercat armonizarea tehnicilor utilizate si in tabelul 12 sunt prezentate doua dintre scehemele de fractionare cele mai utilizate. Cele doua scheme sunt utilizate pentru evaluarea proteolizei in branzeturi in timpul maturarii, prin analiza continutului de N al fiecarii fractiuni utilizand metoda Kjeldhal.
Tabelul 12.
|
Metode de fractionare a componentelor din branza |
|
|
Fractiuni ce contin |
Continut |
|
apa |
Proteine (cu exceptia majoritatii cazeinelor), toate petidele, aminoacizii si compusii mici cu N, cum sunt aminele, ureea si azotul |
|
Acida la pH 4.4-4.6 |
Proteine (cu exceptia majoritatii cazeinelor), toate petidele, aminoacizii si compusii mici cu N, cum sunt aminele, ureea si azotul |
|
12% TCA (acid triclor acetic) |
Peptide de dimensiuni medii si mici, aminoacizi si compusi mici cu N, cum sunt aminele, ureea si azotul |
|
PTA ( acid fosfotungstic) |
Peptide foarte mici, aminoacizi si compusii mici cu n, cu exceptia aminoacizilor dibazici si a amoniacului |
Principala diferenta intre schemele de fractionare consta in prima etapa, care poate fi ori dispersarea branzei in apa, ori dispersarea intr-o solutie de citrat. Solubilitatea cazeinelor in solutie de citrat difera de solubilitatea in apa , atunci cand pH e scazut prin adaugarea de acid;
Cazeinele nu sunt redizolvate in solutia de citrat, atunci cand pH este scazut sub pH-ul izolelectric ceea ce face usor de precipitat toate cazeinele prin scaderea la valoarea pH 4.4. In metodele descrise ulterior se va adauga o cantitate suficienta pentru a asigura scaderea pH-ului aproape de valoarea 4.4.

Fractionarea branzeturilor se realiza anterior si cu ajutorul TCA, deoarece aceasta metoda este utilizata adeseori pentru analiza laptelui (aceasta este explicatia este utilizata in mod eronat in cazul branzei denumirea de NPN-azot neproteic- care, in cazul branzei nu poate fi definit).Exista posibilitatea uneori de a inlocui nPTA-SN cu TCA-SN, ca o masura a gradului de maturare. TCA-SN contine peptide care sunt precipitate cu PTA si care pot fi importante atat pentru structura branzeturilor cat si pentru gustul amar. De obicei, pentru o evaluare in detaliu sunt utilizate ambele metode de fractionare; datorita toxicitatii soventilor utilizati atat pentru fractionarea cu TCA-SN cat si cu PTA-SN precum si necesitatea de a utiliza fractiunile pentru cromatografie, electroforeza si analize senzoriale, mai multe inceracri au fost aplicate in scopul de inlocui reactivii cu etanol. Tehnica pare promitatoare, dar mai sunt necesare cercetari in aceasta directie.
Procedura analizei continutului de N din fractiuni prin metoda Kjeldhal este de lunga durata si adesea extrem de laborioasa. Multe metode au fost evaluate in scopul de a inlocui aceasta metoda, majoritatea analizand continutul de grupe amino libere, si aceastea pot fi aplicate in anumite conditii.Oricum metoda Kjeldhal este necesara pentru evaluarea proteolizei branzei atata timp cat ramane singura metoda care defineste strict ce se masoara in fractiunile de branzeturi, respectiv continutul de N.
Fractionarea branzei cu apa este o metoda foarte folosita, in special pentru branza Cheddar care are valori scazute si relativ constante de pH. Metoda este usor de aplicat, nu se adauga substante chimice suplimentare, ceea ce faciliteaza ulterior liofilizarea fractiunilor si extractia cu solventi organici.
O schema de fractionare care sa determine proteoliza branzei trebuie sa cuprinda analize Kjeldhal pentru N total (TN), fractiunea solubila in apa (WSN), fractiunea solubila in acid triclor acetic (TCA-SN) si fractiunea N solubila in acid fosfotungstic (PTA), respectiv analiza continutului de substanta uscata (DMC-dry matter content).
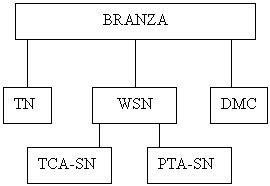
![]()
10 g branza + 50 mL apa deionizata omogenizata 5 min/400C ( utilizand un Stomacher sau un Ultra Turrax -1 min/10 000 rpm, 1 min pauza, 1 min/ 10 000 rpm)
![]()
![]() pastrare
1 h la 400C centrifugare la 3 000 g/30 min/40C sau 30
min/200C si apoi racire la 40C; Suspensia este in final filtrata, de preferata
prin vata de sticla inainte de
determinarea continutului de N prin metoda Kjeldahl.
pastrare
1 h la 400C centrifugare la 3 000 g/30 min/40C sau 30
min/200C si apoi racire la 40C; Suspensia este in final filtrata, de preferata
prin vata de sticla inainte de
determinarea continutului de N prin metoda Kjeldahl.
![]() 25
mL WSN + 25 mL 24% TCA pastrare la temp camerei 2 h si filtrare printr- hartie de filtru
Whatman nr 40;
25
mL WSN + 25 mL 24% TCA pastrare la temp camerei 2 h si filtrare printr- hartie de filtru
Whatman nr 40;
10 mL WSN + 7.0 mL 3.95 mol/L solutie H2SO4 + 3 mL PTA 33%. Concentratia finala a PTA va fi 5%; Amestecul este echilibrat peste noapte la 40C si filtrat cu filtru
Whatman nr 40 inainte de analiza Kjeldahl;
Analize recomandate/ analize duble DMC, analize triple pentru N, TN, doua fractiuni WSN, o fractiune PTA pentru fiecare fractiune WSN si doua titrari Kjeldahl pentru fiecare fractiune.
Fractionarea unei dispersii de citrat a branzei in conditii de pH controlate ofera posibilitatea compararii proteolizei in branza la diferite valori de pH cu perioade diferite de maturare sau obtinuta in procese diferite. Analiza substantei uscate nu e necesara si toate rezultatele se exprima ca % din azotul total (TN), total exprimat pentru aceeasi solutie de citrat. Aceasta usureaza calculul si scade variatia datorata diferentelor dintre mostrele de branza, ca in cazul WSN, NT si respectiv substanta uscata. In plus este deasemenea avantajos ca rezulatele sa nu fie dependente de analizele substantei uscate in branza, deoarece acestea nu sunt intotdeuna usor de obtinut, fara o variatie foarte mare.
In metoda la pH 4.4-SN descrisa o cantitate constanta de acid este adaugata pentru a asigura o valoare a ph -ului apropiata de valoarea.4.4; Aceasta metoda poate fi utilizata pentru majoritatea sortimentelor de branza, dar pH-ul trebuie verificat dupa adaugarea de acid atunci cand un nou sortiment de branza este analizat pentru prima data. O dispreisie de branza in citrat este este realizata si din aceasta dispersie sunt utilizate probe pentru analize Kjeldhal -determinarea azotului total -TN, N solubil la pH 4.4 ( pH 4.4-SN) si N solubil in PTA (PTA-SN). O mostra pentru TCA-SN este luata din fractiunea pH4.4-SN .
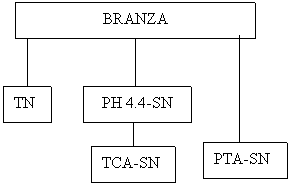
Branza maruntita (10g) + 50 mL solutie calda (40-500C) 0.5 M solutie tri-sodium citrat, amestecata cu amestecator magnetic (!) timp de 60 min si apoi racit la temperatura camerei si adus la 200 mL cu apa deionizata;
Din acesta dispersie de citrat, 10 mL sunt analizati prin metoda Kjeldhal pentru TN; fractionarea la pH 4.4-SN se realizeaza prin adaugarea a 11.3 mL 1M HCl in 80 mL dispersie de citrat, sub agitare puternica, la 120C, obtinand un ph in intervalul 4.35-4.55. Volumul este marit pana la 100 ml prin adaugarea de apa. Proba este filtrata si 25 mL sunt analizati prin metoda kjeldahl pentru pH 4.4-SN.
Pentru a analiza PTA-SN, se iau 40 mL dispersie de citrat si sunt adaugati in 25 mL PTA (20%) si volumul este adus la 100 mL cu H2SO4 (25%) pentru a obtine o concetratie de 2.5% PTA.
Amestecul este pastrat la 40 C peste noapte si filtrat. Pentru analiza Kjeldahl se utilizeaza 50 mL de filtrat.
Fractionarea cu 12% TCA este obtinuta din fractiunea pH 4.4-SN (50 mL proba + 50 mL TCA 24%). Amestecul este pastrat la 40C peste noapte si filtrat, iar apoi analizat prin metoda Kjeldahl. Fractionarea cu TCA trebuie sa fie posibila si din solutia de citrat dar sunt necesare investigatii in aceasta directie.
Analize recomandate: analize duble pentru dispersia de branza cu citrat, o analiza ptr fractiunea de citrat la pH 4.4, o analiza a TCA pentru fiecare fractionare la pH 4.4-SN, una pentru fractionarea cu PTA din fiecare dispersie de citrat, si doua tittrari Kjeldahl pentru fiecare fractiune.
Mii de copusi cu N sunt prezenti in branza si compozitia lor se schimba in timpul perioadei de maturare. Deci copozitia fiecarei fractiuni este complexa si variaza pe parcursul maturarii.
Cazeinele native si peptidele mari produse din actiunea plasminei si a coagulantului sunt precipitate atunci cand branza este dispersata in apa sau cand se adauga acid intr-o dispersie de citrat pentru a cobora valoarea de pH la aproximativ 4.6; In multe cazuri aceste doua fractiuni sunt similare si cantitatea precipitata din branza este similara cu cea precipitata la pH 4.4. o diferenta poate apare pentru branzeturile cu continut ridicat de sare sau pH ridicat, cum sunt branzeturile maurate la suprafata si cele cu mucegai albastru si daca enzimele proteolitice (altele decat cele traditionale) sunt utilizate in timpul procesarii.Comparand doua sortimente de branza (Appenzeller si Parmigiano Reggiano) in privinta continutului WSN si a pH-ului pH 4.4-SN s-a realizat Fig 5; cu exceptia a 3 Appenzeller, toate rezultatele s-au situat pe aceeasi linie, demonstrand aceleasi rezultate pentru ambele metode de fractionare. Cele trei rezultate diferite s-a demostrat ulterior prin analize HPLC ca ele contin o serie de peptide care elueaza tarziu si care nu au fost prezente in alte branzeturi, toate provenind de la aceeasi fabrica de produse lactate;
Mai multe peptide solubile in apa au fost izolate si identificate in Gouda, Parmigian Regiano, Comte, si Cheddar. Peptidele cele mai mari solubile identificate pana acum in branza parmigiano reggiano si Cheddar sunt formate din 20-30 amino-acizi si provin majoritatea din b-cazeina, regiunea cu reziduurile 50-100 si doua peptide mai degraba mari din as2 (3-33, 4-36), care au fost deasemenea identificate in branza Parmigiano Reggiano. Peptidele din as1 -cazeina sunt mult mai rapid hidrolizate in toate sortimentele de branza si doar peptidele mici provenind din as1 -cazeina au fost identificate in branza matura;
Peptidele sun precipitate in 12% TCA deoarece au loc interactii intre TCA si peptide provenite din cel putin doua procese:
interactii electrostatice cu aminoacizii bazici, legaturi de hidrogen cu reziduurile laterale polare ale lanturilor si peptidele schelet
interactii hidrofobe intre cele trei grupari continand clor ale TCA si zonele hidrofobe ale peptidelor (!)
peptidele solubile in 12% TCA variaza in dimensiuni si pot contine de la 2 la 22 de resturi de aminoacizi in timp ce peptidele total insolubile au o lungine mai mare 10 resturi,iar peptidele partial solubile pot varia intre 7-30 de resturi si toate peptidele cu 2 pana la 6 resturi sunt solubile. Numarul fractiunilo cu masa moleculara mai mica de 1 kdalton, solubile in 12% TCA-SN (corespunzator unui nr de 8 aminoacizi) creste in timpul maturarii si se pare ca ajunge la 38, 42 si 62% in branza Emental de 15, 60 si 120 zile. Nu s-a putut stabili nici o corelatie intre indicii de hidrofobicitate si solubilitatea peptidelor in 12% TCA, dar conform timpului de retentie in analiza RP-HPLC cu 0.1% TFA si un gradient de acetonitril peptidele au fost separate in 3 grupe: peptide solubile care au eluat dupa 24%, isolubile, dupa 30 % acetonitril si partial solubile, in intervalul intermediar.
Mai multe peptide fosforilate continand pana la 23 aminoacizi si 1-5 grupe fosfat, provenite in special de la capetele amino-terminale ale b-cazeinei au fost identificate din fractiunea TCA-SN a branzei Parmigiano-Regiano. Majoritatea peptidelor nefosforilate gasite in acelasi studiu au fost formate din doar 5-10 aminoacizi, indicand influenta grupelor fosfat asupra solubilitatii in TCA.
Fractiunea TCA-SN este obtinuta in doua scheme de fractionare si rezultatul este de asteptat sa fie diferit;
Compusii cu N cu masa moleculara pana la aprox 600 Da, continand in principal aminoacizi si foarte putine peptide sunt solubili in PTA. Aminoacizii dibazici (lizina, arginina, ornitina), amoniacul si alti compusi organici cu N sunt precipitati. Partea cu aminoacizi a fractiunii PTA-SN variaza de la un sortiment de branza la altul si cu varsta. Cantitatea de anioacizi din PTA-SN a branzei Cheddar a fost estimata la 75-82% in branza de 6 luni si 78% in branza Herrgard de trei luni.
Cat pentru TCA-SN, comparatiile cu PTA-SN trebuie facute cu precautie si in functie de tipul de fractionare aplica cu apa sau acid citric. Pentru o precipitare corecta este necesara efectuarea combinatiei potrivite intre acidul citric adaugat si cel sulfuric.
Miile de peptide care pot fi prezente in branza sunt fractionate pe baza diferitelor proprietati: cum ar fi masa moleculara, structura conformationala si hidrofobicitate, dar nu cu delimitari foarte exacte/precise. De obicei continutul de N a diferitelor fractiuni ofera cateva informatii despre compozitia compusilor cu N in branza. Primele fractiuni contin mai multe fractiuni succesive asadar este dificil sa fie descrisa distribuia acestor peptide cu proprietati diferite direct din continutul de N al fractiunilor.
Cantitatile estimate de compusi cu N cu diferite proprietati pot fi stabilite prin calcularea diferentelor dintre continutul de N al fractiunilor. (tabelul 12 ).
Tabelul 12.
|
Definirea grupelor de compusi cu N cu diferite proprietati |
||
|
grupe de compusi cu N |
Calcul |
Proprietatile branzeturilor corelate cu fractiunile de N calculate |
|
N cazeinic |
[TN] - [WSN] sau [TN] - [pH 4.4-SN] |
Rezistenta la rupere si intindere Gustul dulce |
|
Peptide mari |
[WSN] - [TCA-SN] sau [pH 4.4-SN] - [TCA-SN] |
Rezistenta la intindere, elasticitatea Gustul amar Peptide care elueaza tarziu din analiza HPLC |
|
Peptide mici |
[TCA-SN] -[PTA-SN] |
Intensitatea aromei branzeturilor, gustul sarat si amar |
|
aminoacizi |
[PTA-SN] |
Intensitatea aromei si gustul sarat |
Denumirile uzulale pentru grupele calculate din table sunt usor de utilizat, dar anumite masuri de precautie trebuie luate deoarece ele nu descriu exact continutul fractiunilor. Fractiunile cazeinice contin cazeinele si produsii primary de hidroliza ai acestora. Trebuie mentionat ca dimensiunea unei peptide incluse in denumirea de peptide mari poate fi mai mica decat dimensiunea unor peptide incluse in categoria peptidelor mici deoarece solubilitatea depinde de mai multi factori, printre care hidrofobicitatea este deosebit de importanta. Compozitia peptidelor din categoria peptidelor mari a fost studiata pentru branza Parmigiano Reggiano. Aminoacizii includ, cu exceptia majoritatii aminoacizilor, cateva peptide chiar mici si alti compusi mici cu N.
Fractionarea compusilor cu N in branza cu etanol a fost evaluata pentru ca era necesar ca acesta sa indeplineasca diferite cerinte.
Noile tehnici analitice utilizate pentru studiul compozitiei fractiunilor, cum ar fi HPLC si electroforeza pot fi aplicate mai usor in aceste conditii.Evaluarea senzoriala poate fi deasemenea aplicata si agentii de precipitare sunt mai putin poluanti pentru mediu.
S-a sugerat ca 70% EtOH ar trebui sa inlocuiasca 12% TCA. In studiile recente s-a aratat ca relatia dintre cei doi agenti de precipitare este destul de complicata.
S-a concluzionat ca extractia cu 12% TCA si 30% EtOH a fost similara in cazul branzei Emmental tinere, in timp ce pentru branza matura, continutul de N dintr-o solutie 12% TCA a fost similar cu cel din 70% EtOH. Branzeturile cu perioade de maturare intermediare trebuie fractionate cu EtOH in concentratii intre 30 si 70 % pentru a obtine aceleasi rezultate ale continutului de N ca in precipitarea cu 12 % TCA. Utilizand electroforeza SDS, Rohm s.a. a aratat ca 70% din N solubil in EtOH are masa moleculara mai mica de 6.2 kdaltoni, in timp ce 50% EtOH dizolva compusi cu masa de pana la 10 kdaltoni si 30% EtOH dizolva compusi Cu masa de 16.9 kdaltoni.
Din moment ce raportul acestor compusi se modifica in timpum maturarii nu e de mirare sa gasim ca diferite concentratii de EtOH corespund la 12% TCA.
Dinamica generala a fractiunilor azotului
In tabelul 13 sunt prezentate analizele fractiunilor azotului si ale principalelor componentele din sortimentul de braanza analizat.
Se poate considera ca toti parametrii analizati au o evolutie normala pentru un proces de maturare. Fractiunile azotului au o evolutie constant crescatoare, cu rate diferite de crestere de la o etapa la alta, in functie de fractiunea analizata.
In cazul analizei fractiunilor azotului s-au inregistrat urmatoarele variatii:
-azotul total a avut variatii pana la 37,8% intre etapele maturarii analizate, lucru explicabil prin analizarea bucatilor in diferite stadii de frabicatie si a bucatilor diferite de bramza in fiecare etapa de maturare;
-azotul solubil in apa a avut o evolutie constant crescatoare, cu o panta mai mica pana la 20 zile, urmata de o panta abrupta pana la 60 zile, dupa care procesul de crestere s-a oprit;
-azotul solubil la pH 4,6 a avut o evolutie constant crescatoare cu o panta mai mica pana la 20 zile, urmata de o panta abrupta pana la 60 zile, dupa care procesul de crestere s-a oprit;
-azotul solubil in TCA are o evoultie ascendenta constanta pe toata durata fabricatiei si maturarii si chiar dupa incheierea maturarii;
-azotul solubil in TCA urmeaza aceeasi traiectorie ca si azotul solubil la pH 4,6, o evolutie constant crescatoare, cu o panta mai mica pana la 20 zile, urmata de o panta mai mare pana la 30 zile dupa care procesul se opreste.
-azotul amoniacal a avut ca si azotul solubil in TCA o evolutie ascendenta constanta pe toata durata fabricatiei si maturarii si chiar dupa incheierea maturarii.
Tabelul 13
|
Nr. Crt. |
Analiza |
Branza inainte de presare |
Cas baschiu la 24 ore |
Branza la 10 zile |
Branza la 20 zile |
Branza la 30 zile |
Branza la 60 zile |
|
Azot total | |||||||
|
Azot solubil in apa | |||||||
|
Azot solubil la pH 4,6 | |||||||
|
Azot solubil in TCA | |||||||
|
Azot solubil la PTA | |||||||
|
Azot amoniacal | |||||||
|
Substanta uscata, % | |||||||
|
Proteine, % | |||||||
|
Proteine/SU, % |
| ||||||
|
Grasime | |||||||
|
Grasime/SU, % |
In ceea ce priveste parametrii SU, grasimea si proteinele se constata o evolutie aproape constanta de la o etapa la alta, cu unele variatii;
-Variatiile substantei uscate sunt date de evolutia apei in semifabricate, la fabricatie, zvantare, maturare si post maturare, cu rate de eliminare a apei mai mari in prima parte dupa care eliminarea apei dupa 20 zile inceteaza;
-Variatiile proteinei in produs sunt date de evolutia eliminarii apei, urmand o panta asemenatoare cu evolutia substatei uscate.
Evolutia fractiunii de azot solubil in apa
Dinamica fractiunii de azot solubit in variantele cu si fara concentrat proteic au fost asemanatoare (Fig.16) . In prima zi de maturare, concentratia azotului solubil in apa raportat la azotul total NSA/NT a fost 20,37% pentru varianta in care s-a folosit contrat proteic (BCP) respectiv 16,45% pentru varianta conventionala (BCN).
Concentratia mai amre de NSA/NT in BCP se datoreaza retinerii unei parti a proteinelor din zer in masa branzei. S-a constatat ca branza Telemea fabricata din lapte UF are un continut mai mare in NSA/NT dacat cea conventionala.
In a doua zi de maturare concentratia de NSA/NT a crecut cu 50% in cazul variantei BCP fata de 100% in cazul variantei BCN. Aceasta diferenta se datoreaza continutului mai mare de apa in BCN, care determina o proteoliza mai avansata la aceasta varianta.
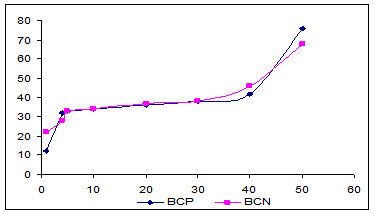
Formarea fractiunilor NSA a continuat mai lent in ambele variante de branza Telemea din ziua a 2-a pana in ziua a 30-a.
Dupa aceasta perioada procesul de degradare a proteinelor s-a accentuat, iar la sfarsitul perioadei de examinare s-au gasit 66,88% in varianta BCP respectiv 76,4% in BCN din azotul solubil in apa.
Evolutia concentratiei cazeinelor si fractiunii de azot solubil la pH 4,6 (4,6-NS)
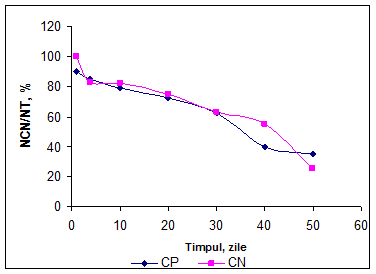
Pe intreaga durata de maturare are loc o reducere continua a concentratiei fractiunii cazeinice (tabel 14).In ambele variante de branza Telemea, degradarea cazeinelor a fost asemanatoare pana in a 5-a zi, cand in varianta CP a continuat aproape liniar pana in a 30-a zi de maturare. Apoi degradarea cazeinelor s-a intensificat pana la sfarsitul perioadei de analiza (a 50-a zi).In varianta CN degradarea cazeinelor a fost redusa pana in a 20-a zi, apoi procesul de degradare s-a intensificat pana in a 40-a zi cand a devenit rapid, pana la sfarsitul perioadei de analiza.
Evolutia franctiunii azotului solubil la pH 4,6 este inversa fata de cea a azotului cazeinic. In cursul perioadei de examinare cresterea concentratiei fractiunii 4,6-NS a fost mai intensa in varianta CP dacat in varianta CN.
In prima zi de maturare concentratia 4,6-NS a fost 5,96% in varianta CP respectiv 6,06% in varianta CN. Pana in a 5-a zi de maturare concentratia a crescut in proportie de 189% in varianta CP, respectiv 166% in varianta CN.
Tabelul 14. Variatia concentratiei cazeinelor pe durata maturarii celor doua variante de branza Telemea
|
Ziua |
Concentratia cazeinelor % |
Perioada |
|
|
CP |
CN |
||
|
Maturare propriu-zisa |
|||
|
Post maturare |
|||
Aceasta diferenta poate fi explicata prin faptul ca varianta CP contine mai multa cazeina provenita din concentratul proteic, prin ultrafiltrare folosit pentru standardizarea laptelui. Aceste cazeine sunt ulterior hidrolizate de dand NS in concentratie mare.
In varianta CP cresterea concentratiei fractiunii NS a continuat ceva mai lent decat la inceput, dar mai repede decat in varianta CN si cu un traseu aproape liniar pana in ziua a 30-a cand cresterea concentratiei fractiunii NS a devenit mai intensiva spre sfarsitul perioadei de analiza.
Cresterea concentratiei fractiunii pH4,6 NS in varianta CN dupa a 5-a zi a fost mult mai lenta si aproape constanta pana in a 20-a zi cand a inceput o crestere semnificativa. Dupa a 40-a zi de analiza s-a constatat o crestere semnificativa.
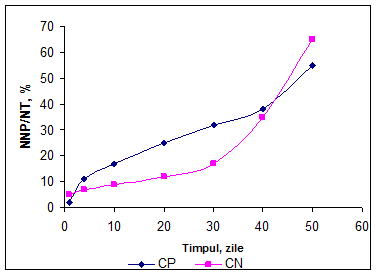
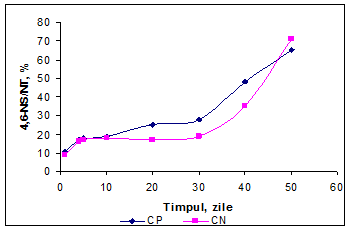
Se observa o crestere cantitativa relativ constanta pentru ambele variante, dar o usoara accelerare a cresterii pentru CN dupa 40 zile.
In primele 5 zile de maturare cresterea concentratiei NNP a fost mai mare in varianta CP si a fost aproape constanta in cazul CP, ceea ce indica o activitate mai mare a endopeptidazelor din microflora branzei in varianta CP, deoarece substratul disponibil este mai bogat comparativ cu varianta CN. In varianta CN cresterea a fost mai lenta pana in a 20-a zi cand s-a inregistrat o crestere aproape logaritmica pana la sfarsitul perioadei de analiza.
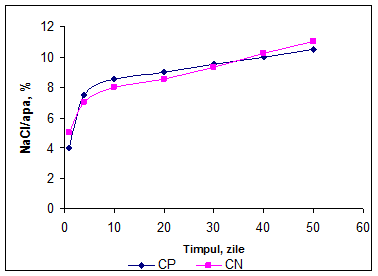
Diferenta in evolutia concentratiei fractiunilor azotoase in cele doua probe poate fi determinata si de concentratia sarii in apa. In fig. 20 se observa evolutia continutului de NaCl in apa, iar in a 5-a zi se inregistreaza o crestere usor mai mare a concentratiei de NaCl/apa pentru varianta CN, comparativ cu CN, ceea ce afecteaza dezvoltarea microorganismelor din cultura starter folosita si inhiba partial activitatea proteolitica a enzimei coagulante reziduale din pasta branzei.
Hidroliza proteinelor din branza in timpul maturarii are o mare influenta asupra structurii, consistentei, digestibilitatii, gustului si aromei branzei maturate. Substantele rezultate din degradarea branzei joaca un rol decisiv si in formarea gustului si aromei specifice sortimentului de branza.
In cercetarile efectuate s-a urmarit procesul de degradare a substantelor proteice si evolutia fractiunilor azotoase in cursul maturarii in doua variante si s-au constatat urmatoarele:
Structura coagulului obtinut din lapte standardizat prin adaugarea de concentrat proteic din lapte obtinut prin ultrafiltrare
Caracteristicile coagulului si ale casului obtinut dupa deshidratarea acestuia sunt hotaratoare pentru proprietatile reologice ale branzeturilor si pentru calitatea acestora.
Sunt cunoscute deosebirile dintre proprietatile coagulului lactic si cele ale coagulului enzimatic definite prin notiunea de porozitate. Porozitatea reprezinta golurile ocupate de faza lichida sau gazoasa. Ele reprezinta diferenta dintre volumul total si volumul substantei uscate, avand valoarea de 90% pentru un coagul din lapte integral, inainte de eliminarea aerului.
Permeabilitatea asigura circulatia zerului catre exterior.Viteza de expulzare a zerului depinde de:
permeabilitatea coagulului
dimensiunea componentelor substantei uscate
deschiderea porilor
forma si dimensiunea porilor
forma spatiilor libere ramase
permeabilitatea pastei branzei
Golurile ocupate de faza lichida intr-un cas format sunt de dimensiuni foarte diferite. Se poate distinge o porozitate specifica granulelor de cas si o porozitate specifica micelelor de cazeina cat si agregate de micele cu o anumita porozitate la interior. Exista si asocieri de porozitate ale elementelor solide de aceeasi marime sau de dimensiune diferita.
Problematica porozitatii este complexa si nu a fost complet elucidata pana in prezent. In cercetarea efectuata s-a urmarit stabilirea influentei proteinelor serice asupra formarii p-cazeinatului monocalcic si implicatiile acestor substante in fromarea structurii pastei.
Variatia concentratiei p-cazeinatului monocalcic
In procesul de fabricatie a branzeturilor proteinele insolubile din coagulul obtinut in urma actiunii enzimei coagulante se tranforma treptat intr-o forma solubila intr-o solutie de NaCl 5%. Aceasta fractiune proteica este reprezentata de p-cazeinatul monocalcic care rezulta in urma actiunii acidului lactic asupra coagulului. P-cazeinatul monocalcic poseda o capacitate adeziva mai mare decat alti paracazeinati, avand in consecinta o influenta importanta asupra proprietatilor reologice ale pastei branzeturilor.
Este evidenta importanta stabilirii concentratiei p-cazeinatului monocalcic si a evolutiei acestuia in cursul maturarii branzei Telemea asupra structurii acestuia in ambele variante de fabricatie: conventional si cu adaus de concentrat proteic din lapte.
Obtinerea concentratului proteic prin ultrafiltrare si adaugarea acestuia in lapte inainte de coagulare modifica raportul dintre CN si PZ ceea ce poate avea consecinte asupra structurii branyeturilor cel putin sub doua aspecte:
1. se reduce proportia de p-cazeinat monocalcic fata de totalul proteinelor din lapte;
2. se mareste gradul de hidratare a proteinelor deoarece este cunoscut ca PZ sunt mai hidrofile decat CN, ceea ce va inluenta structura pastei.
Tabelul 15. Continutul in N din paracazeinatul monocalcic in diferite stadii de fabricatie a branzei Telemea
|
Stadiul de fabricatie |
N-din paraczeinat mg %SU |
Evolutia proportiei de paracazeinat monocalcic fata de coagulul proaspat |
||
|
CN |
CP |
CN |
CP |
|
|
Coagul proaspat | ||||
|
Inainte de sarare | ||||
|
Dupa 8 ore | ||||
|
Dupa 9 zile | ||||
|
Dupa 17 zile | ||||
|
Dupa 24 zile | ||||
In tabelul 15 se prezinta evolutia paracazeinatului monocalcic in cele doua variante de branza cu si fara concentrat proteic pe durata perioadei de maturare.
Tabelul 16. Corelatia paracazeinat monocalcic cu structura pastei
|
Tipul branzei |
p-CN mono.Ca fata de coagul proaspat |
Caracteristica pastei |
Apreciere punctaj total |
|
Telemea- conventional | |||
|
Telemea cu CPL |
In tabelul 16 este prezentata o corelatie intre structura pastei exprimata prin adezivitate si paracazeinatul monocalcic. Se constata ca in varianta CP desi continutul de paracazeinat monocalcic este mai redus in comparatie cu CN, datorita prezentei proteinelor serice retinute in coagul prin UF, aprecierea senzoriala este favorabila.
Determinarea concentratiei de animoacizi din branza
Branzeturile ca si celelalte produse lactate sunt o sursa traditionala de substante nutritive in alimentatie.
Branza contine grasime, provenita din lapte, in procent de 12% in branza proaspata si pana la 20-30% in branzeturile maturate. Aceasta grasime in substanta uscata prezinta un raport echilibrat intre acizii grasi saturati si nesaturati precum si un coeficient de digestibilitate ridicat, de 88-94%.
Dar, importanta nutritiva a branzei se datoreaza cu precadere continutului sau de proteine cu valoare biologica ridicata, branza fiind un aliment proteic prin excelenta. Continutul de proteine variaza intre 10-30%, invers proprtional fata de continutul de grasime. Cca 100 g branza cu consistenta tare acopera 40-50% din necesarul zilnic de proteine pentru un adult.
Branza poate contribui semnificativ la asigurarea aminoacizilor esentiali. In tabelul 17 este prezentata o comparatie intre compozitia laptelui si a branzei, in aminoacizi si proteina de referinta, care indica concentratia ideala a aminoacizilor esentiali in proteina din dieta. Se poate vedea ca proteinele din branza asigura necesitatile de aminoacizi esentiali ca si cele din lapte, cu exceptia celei de metionina si cisteina. Coeficientul de utilizare digestiva pentru proteinele din branzeturi este ridicat, deoarece aceste proteine sunt predigerate printr-o proteoliza bacteriana in timpul maturarii, iar acest coeficient este de 97-98%.
Tabelul 17. Concentratiile in aminoacizi esentiali din lapte si branza in comparatie cu proteina de referinta
|
Aminoacid esential |
Concentratie g/kg proteina |
||
|
Proteina de referinta, FAO/OMS |
Proteine din lapte |
Proteine din branzeturi |
|
|
Triptofan | |||
|
Fenilalanina + tirozina | |||
|
Leucina | |||
|
Izoleucina |
|
||
|
Treonina | |||
|
Metionina+ cistina | |||
|
Lizina | |||
|
Valina | |||
|
Total | |||
Concentratiile de calciu si fosfor ale branzeturilor sunt la fel de importante ca cele ale laptelui, deoarece 100g de branza proaspata acopera 30-40% din necesarul zilnic de Ca si 12-20% din cel de P, iar 100 g branza tare poate asigura complet necesarul zilnic de Ca si contribuie cu 40-50% la completarea necesarului de P.
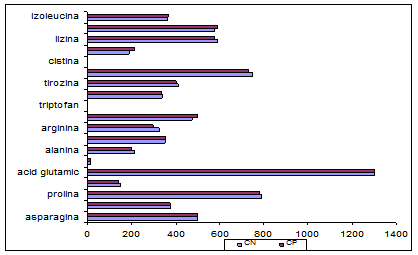
Se observa ca in varianta CP continutul de aminoacizi este de regula mai mare decat pentru varianta CN.
Trebuie remarcat faptul ca un continut mai mare de grasime in branza determina o cantitate mai redusa de Ca si P. Branzeturile produse prin coagulare cu cheag, de obicei, au un continut mai mare de Ca si P decat cele fabricate prin coagularea acida a laptelui.
Concentratiile in g/kg pentru alte minerale variaza in limitele : Na, 0.3-18.5 ; K, 0.5-3.8 ; Mg, 0.1-0.7.
Domeniul valorilor de concetratii (mg/kg) ale altor elemente in branza este: Fe, 0.3-12.0; Cu, 0.2-3.6: Mn, 0.3-5.3; Mo, 0.05-0.5; Zn, 2.7-120; F, 0.1-3.0; I, 0.05-1.0; Co, 0.004-0.038.
Concentratia vitaminelor liposolubile in branza depinde de continutul de grasime. Vitamina A din lapte trece in cantitate mare in branza (80-85%), in timp ce vitaminele hidrosolubile se gasesc in cantitati mici in branza.
S-a determinat efectul utilizarii CPL asupra continutului de aminoacizi din branza. In fig. 21 sunt prezentati aminoacizii continuti de cele doua variante de branza Telemea cu si fara adaos de CP
Concluzii
In urma analizei aminoacizilor din branza s-a constatat o crestere relativa a continutului in aminoacizi in varianta CP, mai ales a celor esentiali, ceea ce determina o crestere a valorii nutritive a branzei fabricate prin standardizarea continutului de proteina in laptele destinat fabricarii branzei Telemea, datorita retineii unei parti din proteinele serice in masa branzei.
Tabelul 18. Continutul de aminoacizi
|
Aminoacid g/kg prot |
Cazeina |
Prot. zer |
Prot. totala |
CP |
CN |
|
Izoleucina* | |||||
|
Leucina* | |||||
|
Lizina* | |||||
|
Metionina* | |||||
|
Cistina* |
Nu |
Nu |
|||
|
Fenilalanina* | |||||
|
Tirozina* | |||||
|
Treonina* | |||||
|
Triptofan* |
Nu |
Nu |
|||
|
Valina* | |||||
|
Arginina | |||||
|
Histidina | |||||
|
Alanina | |||||
|
Acid aspartic | |||||
|
Acid glutamic | |||||
|
Glicina | |||||
|
Prolina | |||||
|
Serina | |||||
|
Asparagina |
Nu |
Nu |
Nu |
Aceasta diferenta este datorata inglobarii proteinelor din zer in masa branzei din varianta CP. Proteinele zerului au o compozitie diferita de cea a cazeinei, ceea ce modifica proportia aminoacizilor comparativ cu varianta CN (Tabelul 18).
Aceasta explicatie este valabila si in cazul celorlalti aminoacizi determinati. Triptofanul si cistina nu au fost determinati din motive tehnice, iar prezenta acizilor aspartic si glutamic si absenta glutaminei si asparaginei pot fi explicate prin transformari metabolice in procesul de maturare al branzei Telemea.
Pentru ambele variante au fost analizate probele in a 20-a zi de maturare.
Deoarece exsista o lunga traditie de a utiliza diverse metode de fractionare si multe sortimente de branzeturi, toate aveand anumite particularitati de fractionare, este deosebit de greu sa standardizam aceste metode. Schemele de fractionare descrise sunt recomandate ori de cate ori nu sunt necesare scheme particulare de fractionare.
Exista doua metode larg raspandite astazi pentru a incepe fractionarea -cu apa sau cu solutie de citrat- deci doua scheme de fractionare recomandate. Multe laboratoare utilizeaza astazi tehnici de varf pentru activitatea de fractionare si deoarece nr de personal este redus aceasta duce la etape de fractionare laborioase, foarte dificil de aplicat. Cele doua scheme de fractionare au fost creeate cu scopul de a facilita activitatea de laborator si in scopul obtinerii unor rezultate adecvate in ceea ce priveste proteoliza branzeturilor.
Rezultatele cercetarilor privind evolutia proteolizei in timpul maturarii propriu zise si post maturare demonstraza complexitatea si dinamica acestui proces. Cu ajutorul metodelor de analiza s-au urmarit procesele caracteristice proteolizei primare si secundare. Evolutia lor cantitativa releva potentialul si secvensa actiunilor sistemelor proteolitice definite si specifice.
In faza initiala intervin enzimele coagulante si plasmina prin actiuni proteloitice specifice si limitate. Hidroliza selectiva conduce la aparitia fractiunilor proteice cu masa moleculara mare.
Complexul enzimatic exopeptidazic extra si intracelular actioneaza asupra acestor fractiuni. Aparitia produselor de hidroliza caracteristice demonstreaza predominanta activitatilor endopeptidazice pe durata intregului proces de maturare.
Evolutia fractiunii de azot solubil in PTA este rezultatul actiunii enzimelor endopeptidazice. Echipamentul enzimatic intracelular al genurilor DDT si LDT cuprimde in preincipal amino si dipeptidaze cat si dipeptidilaminopeptidaze cu specificitate limitata sau foarte larga. Sistemele proteolitice ale speciilor LDB si SST prezinta actiuni sinergetice prin care se intensifica productia de aminoacizi liberi si de substante azotoase cu masa moleculara mica.
Analiza atributelor texturale
Pentru evaluarea atributelor texturale s-au analizat senzorial doua probe de branza cu CP si conventionala CN. La examinare au participat 9 degustatori care au atrbuit fiecarui descriptor analizat un numar de puncte. Prin rotunjirea punctelor acordate s-au obtinut urmatoarele valori:
|
Caracteristici texturale |
Domeniu de punctaj |
Proba1 |
Proba2 |
|
Consistenta | |||
|
Compactitatea | |||
|
Fermitatea | |||
|
Coezivitatea | |||
|
Aderenta | |||
|
Cremozitatea | |||
|
Gumozitatea | |||
|
Asprime | |||
|
Punctaj total | |||
|
Punctaj pondarea cu factor 0.3 |
Se observa ca media punctajului pentru branza CP este superioara celei CN. Diferenta de 6 puncte in favoarea CP se concretizeaza in rezultate mult mai bune in ceea ce priveste consistenta, coezivitatea, cremozitatea si compactitatea branzei. Aplicarea factorului de mediere (0.3) prin care s-a stabilit ponderea caracteristicilor texturale a determinat reducerea diferentelor intre cele 2 probe.
Concluzii
Pe langa cresterea valorii nutritive si micsorarea pierderilor in zer prin fabricarea branzei cu CPL se obtin imbunatatiri semnificative ale atributelor texturale. Aceste imbunatatiri vin in intampinarea cerinetelor consumatorilor privind inoirea si diversificarea produselor traditionale.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3808
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved