| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
Buněčný cyklus a jeho regulace
Gametofyt i sporofyt jsou odvozeny z jedné buňky – gametofyt ze spory, sporofyt ze zygoty. Buňka je základní funkční jednotkou organizmu, nové buňky vznikají dělením buněk stávajících (nevznikají de novo). Mateřská buňka se rozdělí na dvě buňky dceřinné. V mnohobuněčném organizmu s diferencovanými buňkami a pletivy se dělí jen určité buňky. Tyto buňky tvoří specifické soubory – dělivá pletiva neboli meristémy, které jsou v rostlině specificky lokalizovány. Za určitých podmínek, především při porušení integrity rostliny, mohou některé již funkčně specializované buňky schopnost dělení obnovit (totipotence; kap.1.2).
Při dělení buňky se do buněk dceřinných předává úplná genetická informace uložená v jádru, plastidech i mitochondriích. Zatímco organely se rozcházejí do dceřinných buněk jako celky (v počtu přibližně stejném nebo více či méně odlišném), informace v jádře se musí nejprve kompletně zdvojnásobit – replikovat – a odpovídající, věrné a úplné kopie se musí rozdělit do dvou nových celků. Po rozdělení jádra mateřské buňky a vytvoření dvou jader dceřinných – mitóze – se vytvoří buněčná přepážka, která rozdělí cytoplazmu včetně organel a dalších jejích struktur (např. endoplazmatického retikula, vakuol, cytoskeletu) a dokončí vznik buněk dceřinných. Vznik přepážky a oddělení dceřinných buněk se nazývá cytokineze. Jsou-li si dceřinné buňky podobné velikostí i obsahem, označuje se dělení jako ekvální. Jestliže je na anatomické úrovni mezi dceřinnými buňkami patrný rozdíl, nazývá se takové buněčné dělení inekvální. (Inekvální dělení je časté např. při dělení zygoty, nebo u těch buněk prokambia, jejichž dceřinné produkty se diferencují v sítkový element floému a jeho buňku průvodní).
Cytokineze nemusí vždy nutně dělení jádra následovat. Proběhne-li více jaderných dělení bez vytvoření přepážek (v literatuře se lze setkat s termínem volné novotvoření jaderné), vzniká více- až mnohojaderný útvar, zvaný syncytium. Tvorba syncytia je přirozenou součástí určitých ontogenetických fází a v příslušných kapitolách bude zmíněna.
▼ Speciálním případem dělení jádra je dělení redukční – meióza, při níž dochází k rozdělení homologních chromozomů a jejich chromatid do čtyř nových celků a ke snížení počtu chromozomů ve vznikající jádrech ze 2n na n a snížení obsahu DNA na 1C (kap. 10.).
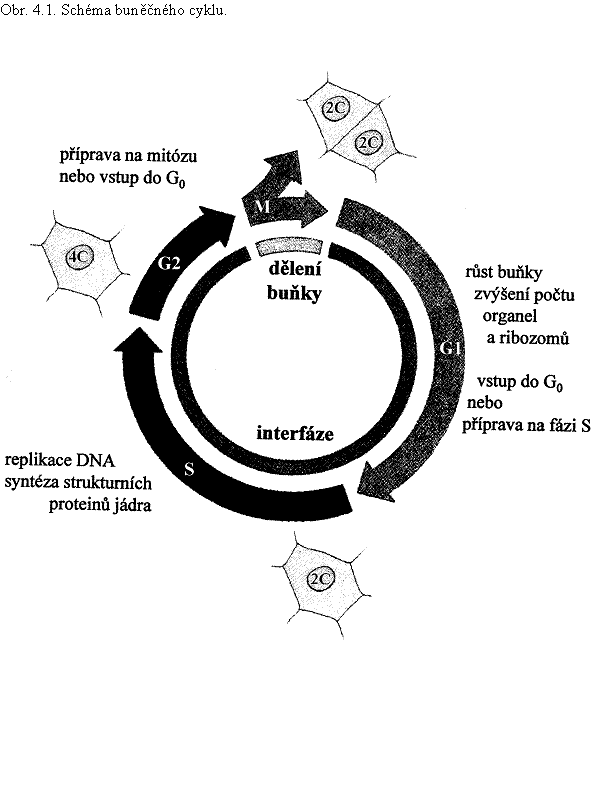
Mitóza (M) je od mitózy následující oddělena interfází (obr. 4.1.). Během interfáze se buňka zvětší a připraví se na vstup do další mitózy. Sled procesů, které se uskuteční během mitózy, cytokineze a interfáze, se označuje jako buněčný cyklus (nedojde-li k cytokinezi, používá se často přesnější název – cyklus jaderný).
Interfáze se dělí na tři fáze, které se značí G1 (z angl. gap – mezera, přerušení, přestávka, propast), S (z angl. synthetic), a G2. O délce buněčného cyklu lze najít v literatuře údaje poněkud rozdílné – u aktivně se dělících buněk se údaje pohybují mezi 19 až 24 h, z čehož mitóza trvá 1 až 2 hodiny. Např. buněčný cyklus buněk aktivního kořenového meristému u Vicia faba trvá 19 h, z čehož fáze G1 trvá pět hodin, fáze S sedm hodin, fáze G2 pět hodina mitóza dvě hodiny. Fáze G1 a G2 jsou obecně časově proměnlivější než M a S, při rychlém dělení se u rostlinných buněk zkracuje především fáze G2.
Ve fázi G1 se buňka zvětšuje, syntetizují se složky buněčné stěny a membrán, zvyšuje se počet ribozomů, zmnožují se organely, metabolicky se buňka připravuje na následující fázi S, vstup do mitózy je blokován. Ve fázi G1 může buňka různě dlouho setrvat nebo z buněčného cyklu vystoupit do fáze G0, přestat se dělit a dle pozice v pletivu a organizmu se funkčně specializovat. Přechod z fáze G1 do S se označuje jako start (angl. start checkpoint), je důležitým kontrolním bodem buněčného cyklu, v němž působí mnoho známých i dosud neznámých regulačních mechanizmů. (V anglické literatuře lze pro fázi G1 najít také označení pre-DNA synthetic phase nebo post-mitotic phase).
Ve fázi S působí regulační i katalytické proteiny a mechanizmy zajišťující iniciaci replikace jaderné DNA, vlastní replikaci DNA (syntéza antiparalelních řetězců k řetězcům stávajícím) a jejich bezchybný průběh a výsledek. Je nutný dostatek nukleotidů (ATP, CTP, GTP, TTP) i strukturních proteinů nutných k uspořádání dvojnásobného množství DNA (např. histonů).
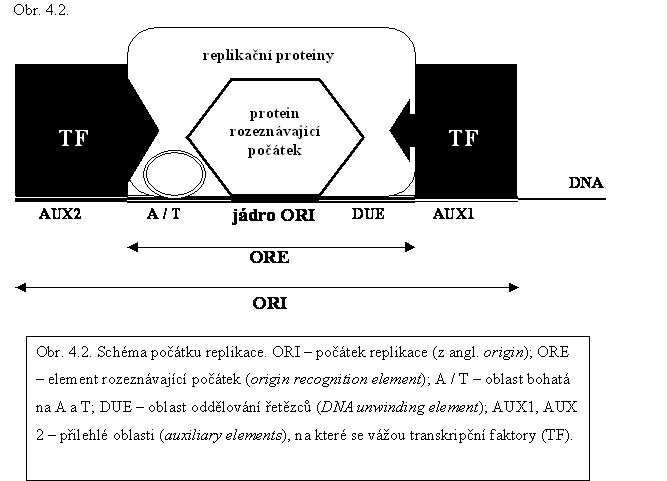
▼Replikace jaderné DNA – syntéza komplementárních řetězců k řetězcům stávajícím – je semikonzervativní, tzn. že v každé ze dvou vznikajících molekul dvouřetězcové DNA pochází jeden z původní DNA a druhý je nově syntetizován. Replikace začíná na určitých nukleotidových sekvencích – replikačních počátcích, značených ORI (z angl. origin replicaton initiation; asi 100 bp; obr. 4.2.). Po celou dobu buněčného cyklu jsou replikační počátky asociovány s proteinovým komplexem zvaným ORC (z angl. origin recognition complex), který slouží jako orientační bod pro nalezení a rozeznání počátku. Replikační počátky jsou na DNA u rostlin dvouděložných rozmístěny ve vzdálenosti asi 66 000 bp a přibližně 47 000 bp u rostlin jednoděložných. Část replikovaná z jednoho počátku se nazývá replikon. V různých počátcích začíná replikace v různou dobu, obvykle se nejdříve replikují aktivně se transkribující oblasti, v nichž je chromatin rozvolněn. Každý replikon se v S fázi zreplikuje jen jednou.
Replikační počátek ORI má tři důležité oblasti – centrální rozpoznávací oblast ORE (z angl. origin recognition element), kam se vážou proteiny důležité k zahájení replikace a dvě přiléhající oblasti – oblast značenou A/T (jeden řetězec je bohatý na T a komplementární na A) a oblast DUE (z angl. DNA unwinding element). Obě oblasti jsou důležité pro oddělení řetězců (odvíjení DNA). Periferně k těmto oblastem přiléhají pomocné elementy, značené aux1 a aux2 (z angl. auxiliary elements), na které se vážou transktipční faktory.
V oblasti počátku, která je bohatá na A a T, enzymy DNA-helikázy katalyzují oddělování řetězců DNA. Separace řetězců probíhá za spotřeby energie, která se získává štěpením ATP. Po oddělení řetězců se na další element replikačního počátku, ORE naváže replikační protein A (RP-A; z angl. replication protein) a další proteiny s katalytickou a strukturní funkcí. Vytvoří se kooperující replikační komplex (aparát), zvaný replikační vidlice. Na rozvinutou jednořetězcovou DNA se vážou proteiny SSB (z angl. single strand binding), které brání opětnému spárování DNA. Replikace probíhá od počátku na obou řetězcích DNA a oběma směry. Vznik fosfodiesterové vazby mezi nukleotidy katalyzují DNA-polymerázy. U eukaryot byly v jádře nalezeny DNA-polymerázy α, δ a ε. DNA-polymerázy připojují 5´-konec nukleotidu na 3´-konec řetězce, tzn. že potřebují primer (syntéza 5´→ 3´). Primery jsou krátké úseky RNA (asi 10 bází), které se syntetizují podle DNA mateřského řetězce. Jejich syntézu zajišťují polymerázy, zvané DNA-primázy. Tyto polymerázy z mateřského řetězce DNA záhy disociují, poskytnou však 3´-konec řetězce, na který může syntéza nového řetězce DNA, katalyzovaná DNA-polymerázami δ a ε, navázat. Podle mateřského řetězce čteného ve směru 3´→ 5´ se nový antiparalelní řetězec syntetizuje ve směru 5´→ 3´. Syntéza tohoto nového řetězce DNA je kontinuální a stačí jí jen jeden primer, nový řetězec se nazývá vedoucí (angl. leading strand). Druhý mateřský řetězec, který se vzhledem k replikačnímu komplexu odvíjí ve směru 5´→ 3´, se replikuje diskontinuálně po krátkých úsecích, z nichž každý potřebuje vlastní primer. Syntézu DNA katalyzuje DNA-polymeráza α. Replikace tohoto mateřského řetězce probíhá pomaleji a vznikající řetězec se označuje jako opožďující se nebo váznoucí (angl. lagging strand). RNA-primer a úsek nového řetězce DNA se nazývají Okazakiho fragmenty. RNA-primery jsou z nového řetězce odstraňovány nukleázou, která štěpí Okazakiho fragment mezi RNA a DNA, a RNázou, která RNA primeru degraduje. DNA-polymeráza, zvaná opravná (charakteru δ/ε), dosyntetizuje chybějící část nového řetězce ve směru 5´→ 3´. Spojení 3´-konce tohoto úseku DNA s 5-´koncem následujícího fragmentu DNA zajišťuje DNA-ligáza. DNA-polymerázy δ a ε jsou k mateřskému řetězci vázány proteinem, který se označuje jako protein svírací (angl. sliding clamp) nebo jako kofaktor PCNA (z angl. proliferating cell nuclear antigen). Tento nezbytný kofaktor DNA-polymeráz obepíná jako prstenec rodičovskou DNA a polymerázu na řetězci přidržuje. Polymerázy mají také exonuklelázovou aktivitu a jsou schopny opravit své případné chyby v předcházejícím kroku.
Replikace končí tam, kde se setkají řetězce různých replikačních počátků. Helikázy, primázy, DNA-polymerázy, DNA-ligázy, PCNA a další proteiny nezbytné pro správnou replikaci DNA tvoří komplex, zvaný replizom.
Lineární DNA chromozómů je ukončena telomerami, oblastmi DNA bohatými na G, na které se váže komplex s enzymatickou aktivitou, zvaný telomeráza. Telomeráza obsahuje RNA, která slouží jako templát pro prodloužení 3´-konce mateřského řetězce DNA. Prodloužený konec poskytne možnost syntézy primerů a umožní vznik nového kompletního nezkráceného řetězce DNA.
Při replikaci DNA, jejíž jeden řetězec je v určitých oblastech metylován (hemimetylovaná DNA), je řetězec, syntetizovaný podle metylovaného řetězce, také metylován.
Plastidy a mitochondrie se během buněčného cyklu množí dle potřeb buňky, replikace jejich genomu a zvýšení počtu organel se časově nemusí krýt s replikací genomu jaderného. Replikace jejich DNA probíhá v příslušné organele vlastním replikačním aparátem.
Po ukončení replikace jaderné DNA buňka vstupuje do fáze G2. Tato fáze je přípravou na vstup do mitózy. Prověří se, zda replikace jaderné DNA byla úplná a zda byly případné chyby opraveny. Přechod z fáze G2 do M je u rostlin předmětem přísnější kontroly než přechod z G1 do S, neboť rostlinná buňka může ve fázi G2 setrvat, přestat se dělit a začít se funkčně specializovat (často se vyskytuje např. u buněk mezofylu). Na konci fáze G2 se připravuje kondenzace chromozomů a jádro se umísťuje do odpovídající pozice v buňce (v případech ekválního dělení je jádro obvykle uprostřed buňky), odstraňují se blokády vstupu do mitózy.
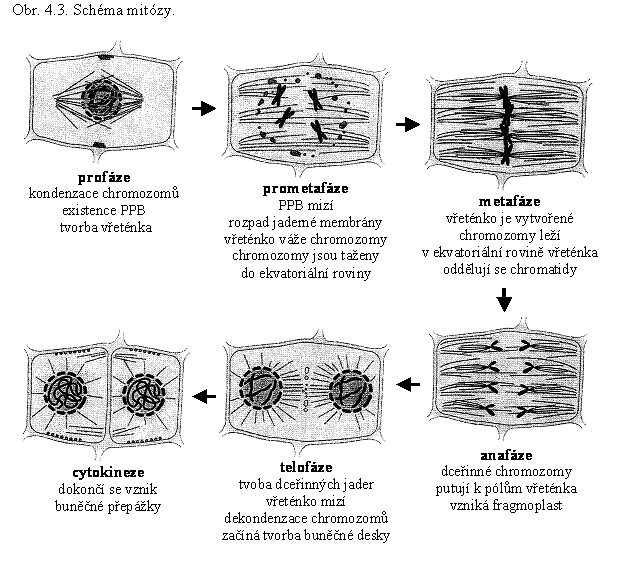
Mitóza, ve skutečnosti plynulý sled událostí, je tradičně dělena na několik fází: profázi, metafázi, anafázi a telofázi (obr. 4.3.).
▼ Profáze chromatin kondenzuje, sesterské chromatidy jsou spojeny v centromeře, jadérko přestává být zřetelné, mikrotubuly začínají tvořit profázní dělicí vřeténko s difúzními póly, v pozdní profázi začíná desintegrace jaderné membrány a preprofázního pásu mikrotubulů – PPB (z angl. preprofase band; kap.4.2).
Metafáze – obalové membrány jádra se rozpouštějí, chromozomy putují do ekvatoriální roviny mitotického vřeténka, mikrotubuly se svými +konci vážou na kinetochory (proteiny vázané na centromery chromozomů).
Anafáze – chromatidy se oddělují a přemísťují k pólům, v ekvatoriální rovině se objevuje fragmoplast.
Telofáze – tvoří se obalové membrány nových jader, chromatin dceřinných jader dekondenzuje.
Jednotlivé fáze mitózy mají vlastní regulační mechanizmy. Rozdělení mateřské buňky na buňky dceřinné je završeno vytvořením buněčné přepážky – cytokinezí.
Buněčný cyklus je řízen, kontrolován a koordinován na mnoha úrovních, četnými mechanizmy a jeho průběh odráží nejen poměry ve vlastní buňce, ale i v buňkách sousedních, v pletivu, orgánu, organizmu i v okolním prostředí. Významnou úlohu v regulaci buněčného cyklu hrají katalytické proteinové komplexy s kinázovou aktivitou, zvané cyklin-dependentní proteinové kinázy, značené CDK.
4.1.Cyklin-dependentní kinázy
■ Regulace buněčného cyklu je dlouhodobě předmětem nejen intelektuálního zájmu ale také experimentální práce na úrovni biochemické a molekulárně biologické. Jako experimentální materiál jsou používány především kvasinky a živočichové. Studium regulace buněčného cyklu se těší neobyčejnému zájmu a podpoře především v medicíně. Přes intenzivní snahu zůstává poznání regulace buněčného cyklu značně neúplné. Informace o regulaci buněčného cyklu u rostlin jsou ve srovnání s informacemi o těchto procesech u kvasinek, živočichů a člověka značně omezené, jakkoli se předpokládá i dokládá, že základní principy a mechanizmy jsou u eukaryot velmi podobné.
Proteinové kinázy jsou enzymy, které katalyzují přenos fosfátové skupiny z ATP na skupinu OH na serinu, tyrozinu, threoninu nebo histidinu příslušného proteinového substrátu. V rostlinách se proteinové kinázy vyskytují ve značném množství – odhaduje se, že geny kódující proteinové kinázy představují 2 až 4% genomu, hrají důležitou roli nejen v regulaci buněčného cyklu, ale i při příjmu a přenosu signálu.
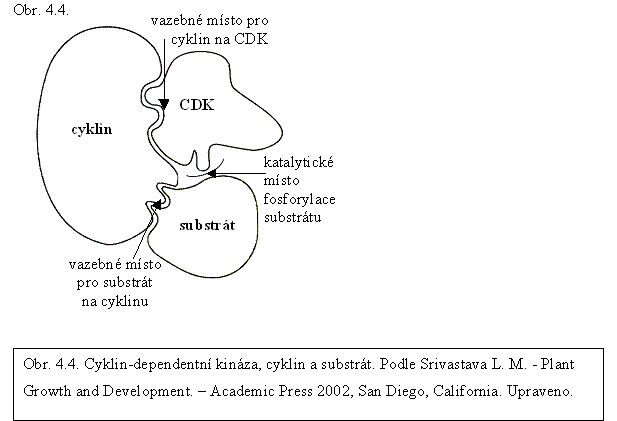
V průběhu buněčného cyklu hrají důležitou roli kinázy, jejichž aktivita a substrátová specifita je závislá na regulačních proteinech zvaných cykliny (obr. 4.4.). Tyto kinázy se označují jako cyklin-dependentní kinázy, CDKs (z angl. cyclin dependent kinases). Syntéza a odbourávání jednotlivých typů cyklinů poskytuje mnoho možností pro řízení buněčného cyklu. Vazba cyklinů na základní katalytickou jednotku kinázy je důležitý mechanizmus regulace schopnosti kinázy fosforylovat další proteiny, není však jediný. Druhý významný mechanizmus je fosforylace a defosforylace základní katalytické jednotky na několika funkčně specifických místech (tyrozinový zbytek v pozici 15 a threoninové zbytky v pozici 14 a 160 nebo 167). Další z dosud známých regulačních mechanizmů je vazba inhibičních proteinů CKIs (z angl. cyclin dependent kinase inhibitors).
Základní katalytické jednotky CDK (dle hmotnosti 33 až 35 kDa značené také p34) jsou produkty malé rodiny genů CDC2/CDC28 (z angl. cell dividing cycle), na N-konci nesou doménu, na kterou se vážou cykliny. U rostlin je dnes známo sedm tříd CDK – značí se A až G a každý protein se může vyskytovat v několika podobách (dále se rozlišují čísly). Přímá regulace buněčného cyklu byla prokázána u kináz třídy A a B – CDKA a CDKB (kinázy tříd C a E působí na úrovni transkripce, D a F ovlivňují buněčný cyklus prostřednictvím dalších kináz, funkce CDK třídy G je zatím neznámá).
Proteiny CDKA jsou přítomny během celého buněčného cyklu. Nesou charakteristickou doménu značenou PSTAIRE (sekvence aminokyselin značených jednopísmenovým kódem = prolin, serin, threonin, alanin, izoleucin, arginin, kyselina glutamová), která umožňuje vazbu cyklinů. CDKA i její mRNA mohou být v malém množství přítomny i v buňkách, které se aktivně nedělí.
Proteiny CDKB se objevují v pozdní fázi G2 a přetrvávají v prvních fázích mitózy. Doména pro vazbu cyklinů je poněkud odlišná, PPTALRE (prolin, prolin, threonin, alanin leucin, arginin, kyselina glutamová) u podtřídy CDKB1 nebo jiná podobná v podtřídě CDKB2. Proteiny CDKB jsou charakteristické pro rostliny, u jiných organizmů zatím nebyly nalezeny. U Arabidopsis jsou v současnosti identifikovány obě podtřídy – CDKB1 i CDKB2, každá je reprezentována dvěma typy.
4.1.1 Cykliny
Cykliny jsou proteiny o hmotnosti 30 až 65 kDa, kódované větším počtem vzájemně příbuzných strukturních genů, jejichž transkripce je řízena různými regulačními signály, např. fytohormony nebo hladinou sacharidů. U rostlin je počet cyklinů ve srovnání s jinými organizmy značně vysoký (v současnosti se udává kolem 60). Cykliny se vážou na základní katalytickou jednotku a vyskytují se jen v buňkách, které se aktivně dělí. Vazbu na CDK zajišťuje sekvence asi 100 aminokyselin, zvaná cyklinový box.
▲ U živočichů je známo 13 tříd cyklinů (značených A až L a T), jednotlivé proteiny se dále rozlišují číslicemi. Rostlinné cykliny lze zařadit do sedmi z nich (A, B, C, D, H, P a T). Nomenklatura a klasifikace se mění a zpřesňuje, což při studiu literárních pramenů činí někdy situaci poněkud nepřehlednou. U Arabidopsis je v době vzniku tohoto textu (2007) identifikováno celkem 32 cyklinů a nomenklaturně zařazeno do kategorií A (10), B (11), D (10) a H (1), odhaduje se, že v genomu může být kódováno asi 49 cyklinů.
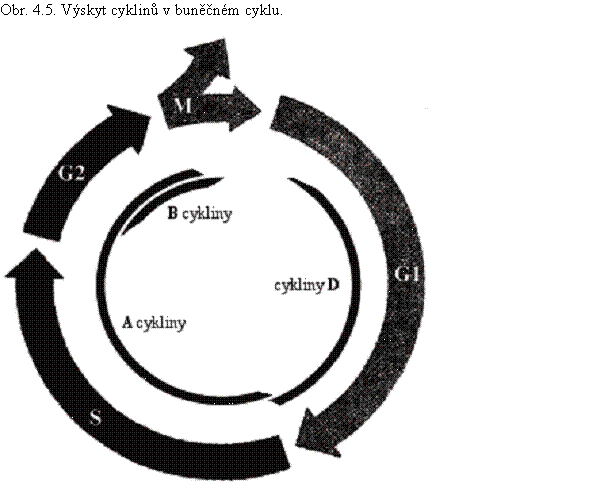
Rostlinné cykliny se značí podle podobnosti s cykliny živočišnými písmeny a čísly (např. A1;1, A2;3, B1;4). Podle fáze buněčného cyklu, ve které se vyskytují a působí, byly cykliny původně děleny do dvou základních skupin – cykliny zvané mitotické (vyskytující se ve fázích S, G2, M) a cykliny fáze G1. Toto dělení se v přehledných pojednáních stále používá. Do skupiny mitotických cyklinů patří cykliny A a B (dříve známé jako M-cykliny), jako cykliny fáze G1 jsou známy především cykliny D (dříve uváděná existence cyklinů E u rostlin byla v poslední době zpochybněna).
Cykliny A se objevují na přechodu fáze G1 do S a přetrvávají do fáze G2, cykliny B jsou důležité pro přechod z fáze G2 do mitózy (obr. 4.5.). Cykliny typu A a B1 mají tzv. mitotický destrukční box, tj. sekvenci aminokyselin, která je rozeznávána ubikvitinovou ligázou E3 a proteiny určuje k rychlému odbourání na proteazomu 26 S během mitózy (kap. 3.4; nejintenzivnější v době přechodu do metafáze). Mechanizmus inaktivace cyklinů B2 během mitózy je zatím nejasný.
Do skupiny cyklinů G1 patří cykliny typu D. Jsou přítomné v G1 fázi a degradované na počátku fáze S. Nemají mitotický destrukční box, jejich vysoká nestálost je dána přítomností sekvencí PEST (prolin, glutamát/aspartát, serin/threonin) na C-konci proteinu. Jejich hladina je ovlivňována mimo jiné sacharózou a cytokininy. Jeden typ cyklinů D, značený CYCD4;1 je přítomný v S, G2 a na počátku fáze M a oba typy CDKB s ním mohou tvořit komplex.
4.1.2 Fosforylace CDK
Fosforylace je další mechanizmus regulující aktivitu a substrátovou specifitu cyklin-dependentních kináz. Základní katalytická jednotka je fosforylována specifickými kinázami a defosforylována příslušnými fosfatázami. V G1 fázi je CDKA fosforylována v pozici Tyr 15 nebo Thr 14, fosforylace v těchto pozicích blokuje vstup buňky do mitózy. Fosforylaci v této pozici katalyzuje kináza WEE1 (ze skotského wee – malý, maličký; podobná kináza byla původně objevena u kvasinek Saccharomyces pombe na základě analýzy mutantu, který se vyznačoval rychlým dělením doprovázeným zmenšováním buněk). WEE1 později fosforyluje i CDKB. Ve fázi S je CDKA fosforylována v pozici Thr 160 (nebo 167), fosforylaci katalyzuje kináza CAK (z angl. CDK activating kinase). Fosforylace na tomto místě je pro vstup do mitózy nezbytná, stejně jako odštěpení fosfátu z pozic na Tyr 15 a Thr 14, ke kterému dochází ve fázi G2. U kvasinek je odstranění fosfátu z této pozice katalyzováno fosfatázou kódovanou genem Cdc25. U rostlin je existence a působení této fosfatázy nejasné a zpochybňované.
▲Na základě funkční podobnosti CDKB1;1 s fosfatázou CDC25 u Drosophila byla vyslovena atraktivní hypotéza, že fosfatáza CDC25 byla u rostlin v evoluci nahrazena tímto pro rostliny specifickým typem CDK.
Syntéza, aktivace a inaktivace kináz a fosfatáz, které katalytickou jednotku fosforylují a defosforylují, představují další oblasti regulace průběhu buněčného cyklu.
4.1.3 Inhibiční proteiny CKI
Inhibiční proteiny CKI tvoří u rostlin skupinu početnou a heterogenní. Charakteristickým znakem je doména tvořená 31 aminokyselinami, u rostlin lokalizovaná na C-konci proteinu a nezbytná pro interakci s CDKA a cykliny – především s cykliny D. Savčí inhibitory CDK mají stejnou doménu na N-konci, značí se Kip (z angl. kinase inhibitory proteins) a na základě této podobnosti se rostlinné inhibitory CDK často značí také KRP (z angl. Kip-related-proteins). U Arabidopsis bylo zatím identifikováno sedm CKI, značených KRP1 až 7. Inaktivace CDK působená CKI nemá destrukční charakter a je reverzibilní.
Syntéza inhibitorů CDK, jejich odbourávání, lokalizace v buňce (cytoplazma – jádro) i migrace do sousedních buněk skrze plazmodezmy poskytují možnost koordinovat dělení buněk v cytoplazmatických doménách i řídit průběh buněčného cyklu působením celé řady vnějších faktorů. CKI jsou rozeznávány a určeny k odbourání komplexními ubikvitinovými ligázami typu SCF (kap. 3.4.
4.1.4 Substráty CDK
Substrátová specifita a aktivita cyklin-dependentních kináz je určena charakterem základní katalytické jednotky (CDKA, CDKB), typem cyklinu a pozicí navázané fosfátové skupiny. Tento systém současně poskytuje mnoho možností pro působení různých regulačních mechanizmů. Zatím nejsou ani zdaleka známy všechny substráty, které cyklin-dependentní kinázy v jednotlivých fázích buněčného cyklu fosforylují.
Ve fázi G1 je jedním ze známých substrátů pro kinázu CDKA/cyklin D komplex transkripčních faktorů skupiny E2F/DP z angl. elongation factor/dimerisation protein) s proteinem pRb (z angl. protein Retinoblastoma; retinoblastom je oční nádor u dětí, způsobený mutací genu RB; protein RB je považován za hlavní brzdu buněčného cyklu a je-li nefunkční, buňky se dělí a tvoří nádor). Transkripční faktory E2F/DP se jako homo- nebo heterodimery vážou na promotory genů, které kódují proteiny ovlivňující replikaci DNA a průběh S-fáze, a také na promotory genů svých vlastních. Mezi geny, které mají v promotorové oblasti jeden nebo více cis-elementů pro vazbu E2F/DP, patří např. geny kódující DNA-polymerázu α, PCNA (svírací protein; kap. 4.) a malou podjednotku ribonukleotidreduktázy.
Ve fázi G1 je na dimery transkripčních faktorů E2F/DP vázán protein pRb a inaktivuje je. Vstup do S-fáze je tak blokován. Komplex CDKA/cyklin D fosforyluje pRb (silně, na mnoha místech) a fosforylovaný pRb se z komplexu uvolňuje. Geny kódující transkripční faktory E2F/DP se exprimují a jejich produkty ovlivňují expresi genů, kódující proteiny nezbytné pro replikaci DNA a průběh fáze S. V pozdní S fázi komplex CDKA/cyklin A fosforyluje transkripční faktory E2F/DP, které tím ztrácejí schopnost se vázat na DNA. Exprese genů kódujících transkripční faktory E2F/DP i replikační a regulační proteiny fáze S ustává (obr. 4.6. ).
▲ U Arabidopsis bylo identifikováno šest různých proteinů E2F, dva DP a jeden pRb, u kukuřice existují tři různé proteiny pRb, pšenice má tři geny pro E2F. (V literatuře lze najít odhad, že u Arabidopsis má asi 6000 genů v regulačních oblastech jedno nebo více vazebných míst pro proteiny E2F/DP, celkový počet míst se odhaduje na 10 000.).
Při přechodu z fáze G2 do M byly u Arabidopsis prokázány funkční komplexy CDKA/cyklin A, CDKB/cyklin B a CDK/cyklin D4;1, základní jednotka nesmí být fosforylovaná v pozici Tyr15 a musí být fosforylovaná na Thr160. První komplex (objevený na začátku 80. let minulého století ) byl označen jako MPF (z angl. mitosis promoting factor) a toto označení se souhrnně stále používá pro všechny typy komplexů u rostlin i u jiných organizmů.
MPF fosforyluje řadu strukturních proteinů přímo, další nepřímo tím, že fosforyluje a tak aktivuje další proteinové kinázy. Fosforylované proteiny mění svou funkci, často jsou rozeznávány ubikvitinovými ligázami E3, které zajistí jejich polyubikvitinaci a určí je k degradaci proteazomem 26S.
Substráty, které MPF fosforylují na přechodu z fáze G2 do mitózy, v profázi a metafázi u rostlin jsou ve srovnání s kvasinkami a živočichy doloženy značně omezeně. Substrátem je např. histon H1 (význam jeho fosforylace a degradace není však dostatečně objasněn; může se jednat o nadbytečný H1). Přímo nebo nepřímo jsou fosforylovány proteiny umožňující existenci euchromatinu a proteiny, které zajistily desintegraci obalových membrán jádra v profázi. Fosforylovány jsou proteiny asociované s mikrotubuly, mikrotubuly jsou destabilizovány což umožňuje dynamické přeměny struktur cytoskeletu během mitózy, např. rozpad preprofázního pásu a tvorbu vřeténka. Fosforylovány jsou RNA-polymerázy II, poly(A)-polymerázy, což souvisí s vysokým stupněm kondenzace chromatinu, který omezuje transkripci. Významná je přímá fosforylace cyklinů (volných i vázaných v MPF), která vrcholí v metafázi, vede k inaktivaci MPF a umožňuje dokončení mitózy. V pozdní metafázi jsou fosforylací jedné z podjednotek aktivovány komplexy APC (z angl. anaphase promoting complex; lze najít i označení cyklozom – APC/C), které působí jako E3 ligázy (kap. 3.4). APC určí k degradaci fosforylované cykliny, nepřímo navodí také odbourání proteinů působících soudržnost chromatid a umožní tak jejich oddělení.
▲APC komplexy (APC/C) jsou u rostlin tvořeny (dle poznání v r. 2007) 12 podjednotkami. Podjednotky APC2 a APC11 tvoří katalytické jádro komplexu a slouží k interakci s konjugačními enzymy ubikvitinu E2. Další podjednotky komplexu, tzv. selektorové proteiny, rozeznávají a podílejí se na vazbě proteinů, určených k ubikvitinaci. Komplexy APC se vyskytují v mnoha podobách a vážou ubikvitin na celou řadu substrátů, např. na fosforylované cykliny A, B (volné i vázané v MPF). Důležitým substrátem je také sekurin, který inhibuje činnost separázy – enzymu štěpícího koheziny. Koheziny jsou proteinové komplexy zajišťující soudržnost sesterských chromatid (syntetizují se už v S fázi). APC tak nepřímo umožňuje dokončení oddělení chromatid v pozdní metafázi a nástup anafáze. Na rozdíl od E3 ligáz typu SCF, ligázy typu APC rozeznávají fosforylované i nefosforylované proteiny.
4.2 Cytokineze
Cytokineze je vytvoření buněčné přepážky mezi dceřinnými buňkami a představuje konečnou fázi rozdělení buňky. Rovina, v níž přepážka vznikne, je určena již na konci fáze G2 a souvisí s polohou jádra v buňce před vstupem do mitózy. Vstup do mitózy je provázen výraznými změnami v uspořádání kortikálního cytoskeletu (buněčným kortexem se rozumí oblast cytoplazmy pod plazmatickou membránou, bohatá namikrotubuly a aktinová filamenta).
Na konci fáze G2 a v profázi z kortikálního cytoskeletu mizí mikrotubuly a zůstávají jen v prstenci, v jehož centru leží jádro. Tento prstenec mikrotubulů přiléhající k plazmalemě se označuje jako preprofázní pás, značený PPB (z angl. preprophase band). PPB mizí v pozdní profázi I přes časově omezenou existenci PPB se předpokládá, že PPB vymezuje ekvatoriální rovinu dělicího vřeténka a předurčuje rovinu, v níž vznikne buněčná přepážka.
Předpoklad, že poloha PPB souvisí s umístěním jádra v buňce, byl experimentálně potvrzen. Přemístění jádra na začátku profáze vedlo k dezintegraci existujících základů PPB a vytvoření nového PPB, jehož lokalizace odpovídala nové poloze jádra.).
Současně s rozpadem mikrotubulů preprofázního pásu mizí z oblasti buněčného kortexu také aktin a vzniká bezaktinová zóna, označovaná ADZ (z angl. actin depleted zone), která přetrvává po celou mitózy. Do této oblasti migrují vezikuly Golgiho aparátu a cisterny endoplazmatického retikula. Tato zóna bývá v literatuře označována jako Golgiho pás (angl. Golgi belt).
Při tvorbě buněčné přepážky hraje důležitou úlohu fragmoplast, Golgiho aparát a endoplazmatické retikulum.
Fragmoplast je dynamická cytoskeletální struktura, charakteristická pro tzv. „ konvenční“ cytokinezi u somatických buněk sporofytu Začíná se tvořit v pozdní anafázi v centrální oblasti ekvatoriální roviny vřeténka v podobě dvou antiparalelních (rovnoběžných avšak zrcadlově obrácených) prstenců volných mikrotubulů vřeténka, mikrotubulů nově tvořených, aktinových filament a proteinů s těmito cytoskeletálními strukturami asociovanými. Mikrotubuly i mikrofilamenta stojí kolmo k rovině, v níž vznikne přepážka. Polární orientace mikrotubulů je stejná jako u dělicího vřeténka (+konce směřují k ekvatoriální rovině, -konce k pólům). Fragmoplast slouží především transportu vezikulů, které se oddělují z trans-cisteren diktyozomů (Golgiho aparát) a nesou materiál na tvorbu přepážky mezi dceřinnými buňkami. Předpokládá se, že fragmoplast funguje i jako konstrukce pro vznik a stabilizaci nejranější podoby buněčné přepážky. Současně vymezuje zónu, v níž se vedle Golgiho vezikulů vyskytují jen malé cisterny endoplazmatického retikula (zřídka i velmi malé vakuoly) a v níž chybí plastidy, mitochondrie, ribozomy i diktyozomy (v anglické literatuře se vyskytuje termín zone of exclusion, který lze přeložit jako zakázaná zóna).
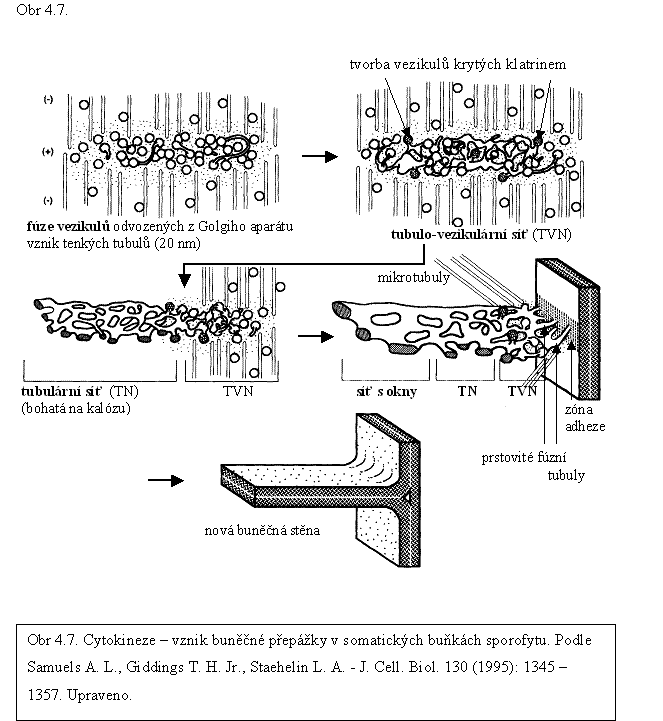
Přepážka má nejprve podobu ploché cisterny diskovitého tvaru, zvané buněčná deska (angl. cell plate). Je tvořena membránou na povrchu a polysacharidy, především pektiny, hemicelulózami a kalózou v lumenu. Vzniká splýváním vezikulů v ekvatoriální rovině dělicího vřeténka. V meristémech sporofytu se buněčná deska začíná tvořit v centrální části ekvatoriální roviny a roste centrifugálně, tj. směrem k plazmalemě mateřské buňky, se kterou se spojí v oblasti předem určené již na konci interfáze preprofázním pásem mikrotubulů (PPB). Buněčné deska se utváří v několika plynule navazujících vývojových fázích, které na rostoucí desce existují současně (obr. 4.7.).
▲ První fází tvorby buněčné desky je transport, hromadění a splývání vezikulů Golgiho sítě (obr. 4.7.). Vezikuly (o velikosti asi 60 nm) tvoří velmi dlouhé tenké vychlípeniny tubulárního (trubicovitého) charakteru (průměr asi 20 nm), jimiž se kontaktují a vzájemně propojují v kontinuální hustě propletenou tubulo-vezikulární síť prostoupenou cytosolem (původní kulovité vezikuly jsou na síti identifikovatelné, průměr tubulů se zvětšuje na přibližně 35 nm). Splývající vezikuly jsou obklopeny hustou cytoplazmou bez ribozomů a organel (angl. fuzzy coat). V membráně se začíná syntetizovat kalóza, která se ukládá do vnitřního prostoru (lumenu) buněčné desky. Tubulo-vezikulární síť se postupně mění ve volně propletenou hladkou tubulární síť, která je uspořádana v několika paralelních vzájemně propojených rovinách a zvětšuje se asociací s dalšími vezikulárními strukturami. V membráně je syntetizováno značné množství kalózy, začíná se objevovat celulóza tvořící ojedinělé mikrofibrily. Mikrotubuly fragmoplastu v této fázi depolymerizují, nové mikrotubuly fragmoplastu se tvoří na obvodu buněčné desky, tzv. „vedoucím“ okraji a průměr fragmoplastu se zvětšuje. V dalším vývoji se tubuly zplošťují a buněčná deska nabývá podoby vlnící se plošné, hladké sítě s velkými oky (angl. fenestrated sheet), zvanými též póry, otvory nebo kanálky. Membrána buněčné desky zůstává v přerušeních kontinuální. Vznik plošné hladké sítě je provázen zvýšenou syntézou celulózy, jejíž množství však ve srovnání s primární buněčnou stěnou mateřské buňky zůstává velmi nízké. Oky buněčné desky mohou procházet prstovité výběžky nebo provazce membrány endoplazmatického retikula, kolem nichž zůstává zachován nepřerušený pruh cytoplazmy. V těchto místech se mohou vytvořit primární plazmodezmy nebo se oka uzavřou.
Spojení buněčné desky s plazmalemou mateřské buňky začíná tvorbou velkého množství prstovitých výběžků na okrajích buněčné desky (angl. finger tubes), které se prodlužují a splývají s plazmatickou membránou mateřské buňky. K pruhu plazmalemy, k němuž se buněčná deska připojuje, přiléhal na začátku mitózy preprofázní pás. Existuje představa, že plazmalema byla v těchto místech preprofázním pásem modifikována a tak ke spojení s „vedoucím“ okrajem buněčné desky předurčena. (Modifikace plazmalemy odolává mechanické zátěži centrifugací, může však být narušena plazmolýzou, mechanickým poraněním nebo působením určitých látek, např. metanolem).
Buňky jsou definitivně oddělené ve chvíli, kdy membrána buněčné desky splyne s plazmalemou na celém svém okraji. Splynutí s plazmalemou buněčnou přepážku prostorově stabilizuje (ustává vlnění). Membrána buněčné desky se postupně mění ve funkční plazmalemu, v lumenu se formuje primární buněčná stěna, napojující se na primární buněčnou stěnu mateřské buňky. V membráně výrazně stoupá množství i aktivita celulózasyntázových (terminálních) komplexů. Pohyb komplexů v membráně a směr ukládání vznikajících celulózních mikrofibril do amorfní polysacharidové matrix ovlivňují mikrotubuly kortikálního cytoskeletu, který se opět formuje. Pektinová vrstva střední lamely se diferencuje od střední lamely mateřské buňky centripetálně.
▲Syntézu dalších strukturních polysacharidů (hemicelulóz a pektinů) a strukturních i katalytických proteinů buněčné stěny zajišťuje endoplazmatické retikulum a Golgiho aparát, materiál je dopravován ve vezikulech anterográdním transportem. Přebytečný nebo nevhodný materiál je z buněčné stěny a plazmatické membrány odstraňován retrográdním transportem vezikulů krytých klatrinem po celou dobu vzniku buněčné desky a její přeměny ve zralou plazmalemu a buněčnou stěnu.
Vznik přepážek v meiocytech a celularizace syncytií, např. nukleárního endospermu, probíhá poněkud odlišně, např. se netvoří preprofázní pás mikrotubulů, který určuje rovinu vznikající přepážky, vznikají sekundární (adventivní) fragmoplasty mezi nesesterskými jádry na styčných plochách nukleárně cytoplazmatických domén (NCD). Tyto procesy budou zmíněny v odpovídajících kapitolách (kap.7.1.1, kap. 10.1).
Plazmodezmy jsou struktury propojující protoplasty sousedních buněk v symplast. Přerušení buněčné stěny, na příčném řezu kruhovité, je kryté plazmatickou membránou, ke které přiléhá vrstva cytoplazmy (tzv. cytoplazmatický rukáv, angl. cytoplasmic sleeve). Středem plazmodezmu prochází stěsnaný válcovitý útvar tvořený lipidy, který propojuje membrány endoplazmatického retikula sousedních buněk – dezmotubulus. Centrální část dezmotubulu je vyplněna proteiny (anglicky se tato výplňová struktura označuje jako rod). Lumen endoplazmatického retikula, na rozdíl od plazmalemy, cytoplazmy a membrány ER, není kontinuální nebo že je jeho kontinuita značně omezená. Vnitřní (cytoplazmatická) strana plazmalemy v plazmodezmu a vnější povrch lipidové vrstvy desmotubulu jsou kryty globulárními proteiny. Tyto dvě proteinové vrstvy jsou propojeny proteiny vláknitými (angl. spoke proteins), které cytoplazmu dělí obvykle na sedm až devět podélných pruhů. Toto rozdělení patrně ovlivňuje a určuje velikost molekul, které mohou plazmodesmy procházet difúzí (charakteristika: SEL = size exclusion limit 700 až 1000 daltonů; je modifikovatelná). Vstup do plazmodezmu může být na jedné nebo na obou stranách oproti střední části zúžen. Primární plazmodezmy vznikají při tvorbě přepážky mezi dceřinými buňkami, sekundární plazmodezmy vznikají podle aktuální potřeby transportu zvětšující se nebo senescentní buňky. Plazmodezmy mohou také aktivně zanikat, např. ve stresových situacích jako je napadení virem nebo plazmolýza. Některé viry však mohou plazmodezmy modifikovat a touto cestou se v rostlině šířit.

|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2324
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved