| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
DOCUMENTE SIMILARE |
|
POLISACÁRIDOS
Concepto y clasificación
Los polisacáridos están formados por la unión de más de 12 monosacáridos por enlaces O-glicosídicos. Se suelen clasificar como:
homopolisacáridos: formados por el mismo monosacárido
heteropolisacáridos: “ por varios tipos de monosacáridos diferentes
Otra clasificación es la de polisacáridos
simples : formados por monosacáridos simples (Glc,Gal)
derivados: “ por derivados de monosacáridos (acetilados, azúcares ácidos)
Glucanos más abundantes
La celulosa es la molécula de origen natural más abundante que existe en la naturaleza. Esto se debe a que la celulosa forma parte de las paredes celulares de los vegetales, y estos forman una gran biomasa. Su función es pues, estructural.
Vista al microscopio electrónico, la celulosa está formada por fibras que están, a su vez, formadas por un conjunto de cadenas de glucosa no ramificadas y dispuestas en paralelo unas a otras.
Todas las unidades son ![]() -D-Glc unidas por enlaces glicosídicos
-D-Glc unidas por enlaces glicosídicos ![]() (1 4). Es un
“homopolisacárido simple”. La unidad repetitiva es el disacárido “celobiosa”.
(1 4). Es un
“homopolisacárido simple”. La unidad repetitiva es el disacárido “celobiosa”.
La celulosa se organiza en cadenas lineales, y estas en fibras, por el establecimiento de puentes de H.
Dentro de la propia cadena de celulosa se pueden establecer puentes de H que mantienen esta estructura lineal, pero también entre cadenas paralelas de celulosa.
Esta estructura es ideal para su función biológica. Su elevada capacidad de formar puentes de H le da firmeza y es responsable de la insolubilidad en agua de este polisacárido.
En las paredes celulares de las plantas, estas fibras de celulosa se refuerzan por la presencia de otras sustancias que actúan a modo de cemento, como son la “hemicelulosa”, “pectina”, y “lignina”.
La hemicelulosa es un polisacárido formado por D-xilosa. La pectina está formada por el ácido D-galacturónico, y la lignina es una sustancia más compleja (polifenólica).
Todas ellas contribuyen a la rigidez de la pared.
La celulosa no puede ser digerida por los animales superiores excepto en el caso de los rumiantes. Además de estos últimos, también pueden aprovecharla diversos microorganismos, insectos, caracoles A todos ellos se les denomina “celulolíticos”.
La quitina es otro polisacárido relacionado
con la celulosa. Está formada por la unión de moléculas de
N-acetil-glucosamina, unidas por enlaces ![]() (1
(1
Al igual que sucedía con la celulosa, como
los enlaces son ![]() (1 para que el enlace glicosídico se pueda
formar, una molécula tiene que estar invertida con respecto a la otra.
(1 para que el enlace glicosídico se pueda
formar, una molécula tiene que estar invertida con respecto a la otra.
La unidad repetida en este caso es el disacárido “quitobiosa”.
Como en la celulosa, en la quitina se pueden formar puentes de H, y por tanto es un polisacárido que también cumple una función estructural. Es el principal componente del exoesqueleto de artrópodos y también de las paredes celulares de microorganismos como levaduras y hongos.
Polisacáridos con función de almacenamiento
El almidón es un polisacárido que está
presente en los vegetales de modo exclusivo. Está formado por la unión de
moléculas de glucosa (![]() -D-Glc), y nos lo encontramos en forma de gránulos
citoplasmáticos o también dentro de los plastos de los vegetales.
-D-Glc), y nos lo encontramos en forma de gránulos
citoplasmáticos o también dentro de los plastos de los vegetales.
Los enlaces pueden ser de 2 tipos porque el almidón está formado por 2 componentes, la “amilosa” y la “amilopectina”.
En la amilosa, los enlaces son todos ![]() (1 4),
mientras que en la amilopectina, son también
(1 4),
mientras que en la amilopectina, son también ![]() (1 4), pero
a intervalos regulares nos encontramos con enlaces
(1 4), pero
a intervalos regulares nos encontramos con enlaces ![]() (1 6). Es
decir, la amilosa está formada por cadenas lineales, y la amilopectina por
cadenas ramificadas, de modo que cada enlace
(1 6). Es
decir, la amilosa está formada por cadenas lineales, y la amilopectina por
cadenas ramificadas, de modo que cada enlace ![]() (1 6)
representa un punto de ramificación.
(1 6)
representa un punto de ramificación.
La estructura es estos dos componentes es la siguiente:
La amilosa es helicoidal, debido al establecimiento de puentes de H. En las cadenas de amilosa también se establecen puentes, pero de modo diferente a los de la glucosa.
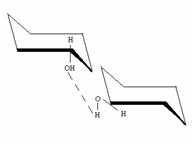
La estructura secundaria de la amilopectina no se conoce con tanto detalle. Posee forma arborescente debida a las ramificaciones.
Cada molécula de celulosa presenta un extremo reductor y uno no reductor. Esto mismo pasa con la amilosa.
En el caso de la amilopectina, una molécula tiene un único extremo reductor pero muchos no reductores, debido a sus ramificaciones (tantos como ellas).
Otro polisacárido que también está relacionado funcional y estructuralmente con la amilopectina es el glucógeno, polisacárido de reserva del medio animal. Se encuentra en el hígado y en el músculo.
Su estructura es similar a la de la
amilopectina, pero este está más ramificado. Los enlaces son ![]() (1 6), y se
sitúan cada 8 unidades de glucosa en el
glucógeno, y en la amilopectina cada 25 – 30. Ambos son digeribles por los
animales superiores (con encimas “amilasa”).
(1 6), y se
sitúan cada 8 unidades de glucosa en el
glucógeno, y en la amilopectina cada 25 – 30. Ambos son digeribles por los
animales superiores (con encimas “amilasa”).
Mucopolisacáridos ácidos o glicosaminoglicanos
Son importantes en el reino animal, y más en los vertebrados. La características generales de este tipo de polisacáridos son que:
están formados por aminoazúcares, normalmente por N-acetil-glucosamina y N-acetil-calactosamina, y de aquí sus nombres.
Suelen poseer grupos cargados negativamente (carbonato, sulfato).
No suelen existir en estado libre, sino asociados a proteínas, formando los proteoglicanos, o también a asociaciones de lípidos y proteínas.
Dentro de estos glicosaminoglicanos vamos a estudiar la estructura de tres fundamentalmente:
El “ácido hialurónico” lo encontramos en el tejido conectivo (en la sustancia basalque rellena los espacios extracelulares de éste), en el líquido senovial, humor vítreo y cordón umbilical.
Se trata de un polímero lineal no
ramificado. La estructura que se repite es un disacárido formado por la unión
del ácido D-glucurónico y de la N-acetil-glucosamina, mediante enlaces ![]() (1 3). Cada
disacárido se une al siguiente por enlaces
(1 3). Cada
disacárido se une al siguiente por enlaces ![]() (1 4). Estos
dos enlaces se alternan en la cadena.
(1 4). Estos
dos enlaces se alternan en la cadena.
Presentan una gran viscosidad, debida a la
presencia de estos grupos ácidos ![]() que a PH fisiológico
presentan carga negativa, están ionizados(
que a PH fisiológico
presentan carga negativa, están ionizados(![]() ), lo que favorece una hidratación excesiva, y la formación
de puentes de H intercatenarios.
), lo que favorece una hidratación excesiva, y la formación
de puentes de H intercatenarios.
Otros dos glicosaminoglicanos también
asociados con el tejido conectivo son el “sulfato de condroitina o condroitín
sulfato” y “el queratán sulfato”. En ambos, el ptatrón de enlaces glicosídicos
es alternante ![]() (1 3),
(1 3), ![]() (1 4) como
en el caso del ác. hialurónico.
(1 4) como
en el caso del ác. hialurónico.
En su composición intervienen
N-acil-azúcares y también llevan grupos cargados negativamente, pudiendo
aparecer grupos sulfato ahora (![]() ). En el caso del condroitín , el disacárido está formado por
el ác. D-Glucurónico y por la N-acetil-D-galactosamina. Aquí la N-acetil-gal
está sulfatada en el
). En el caso del condroitín , el disacárido está formado por
el ác. D-Glucurónico y por la N-acetil-D-galactosamina. Aquí la N-acetil-gal
está sulfatada en el ![]() .
.
La longitud de la cadena del sulfato de
condroitina puede variar entre 15 y 150 unidades de disacárido, y la extensión
de la cadena, así como el grado de sulfatación dependen de la edad del tejido y
de algunas condiciones patológicas. En el caso del sulfato de queratán, el
disacárido repetido está formado por la galactosa y la N-acetil-galactosamina,
que se unen por enlaces ![]() (1 4) y
(1 4) y ![]() (1 3) el
intermediario. La galactosamina también está sulfatadoa en el
(1 3) el
intermediario. La galactosamina también está sulfatadoa en el ![]() . Aquí, a diferencia, interviene la galactosa simple, y no
tenemos grupos carboxilato.
. Aquí, a diferencia, interviene la galactosa simple, y no
tenemos grupos carboxilato.
Aquí la longitud de cadena oscila entre 10 y 50 unidades de disacárido y es menor que la del sulfato de condroitina.
![]()
En resumen: Ácido hialurónico grupos carboxilato
Base común Heparán sulfato grupos carboxilato y sulfato
Queratán sulfato grupos sulfato
Otro tipo de mucopolisacáridos ácidos forman las paredes de las bacterias, y son un entramado estructural que rodea completamente a la célula.
Es una estructura continua (como una bolsa), constituída por cadenas de polisacáridos lagas y paralelas que están unidas entre sí transversalmente a ciertos intervalos por cadenas laterales que son péptidos cortos.
Los polisacáridos están formados por la
unión de la N-acetil-D-glucosamina y el ácido N-acetil-murámico mediante
enlaces ![]() (1 4). El
componente ácido es un monosacárido complejo que lleva un radical en el
(1 4). El
componente ácido es un monosacárido complejo que lleva un radical en el ![]() , lugar donde va unida la cadena lateral polipeptídica corta
que hace de unión d entre polisacáridos.
, lugar donde va unida la cadena lateral polipeptídica corta
que hace de unión d entre polisacáridos.
El péptido suele ser un tetrapéptido cuya composición varía entre las especies bacterianas.
A esta estructura continua que rodea a la célula se le denomina “péptidoglucano” o “mureína”. Hay algunos antibióticos como la penicilina, cuya acción consiste en inhibir una de las múltiples etapas de la síntesis encimática de los peptidoglucanos.
Enlaces azúcares – proteínas
Los dos más importantes que suelen aparecer en las proteínas son:
N-acetil glucosamina + cadena proteica: la unión se establece por el grupo amino de la “asparagina” y se llama una N-GLICOSILACIÓN.
Otro tipo de enlace es un O-glicosídico con la treonina o serina. O-GLICOSILACIÓN.
EJERCICIOS:
1.- Una muestra de 75 mg de celulosa se hidrolizó en medio ácido. En el hidrolizado se encontraron 75 mg de glucosa. scuál es la pureza de la muestra de celulosa?
Como la unión Glc-Glc es un enlace
glucosídico, se gana una molécula de ![]() en cada hidrólisis de 2 moléculas, y su peso hay que
descontarlo.
en cada hidrólisis de 2 moléculas, y su peso hay que
descontarlo.
El peso molecular de la glucosa es 180.
El peso “ de las glucosas unidas por
enlace glicosídico es 180 – 18(peso del ![]() ) 180 – 18 = 162.
) 180 – 18 = 162.
Entonces, la relación de pérdida de masa
será de ![]() , que a su vez es el peso que aumentamos al hidrolizar, ya
que al romper los enlaces les aportamos la molécula de agua que habían perdido
al formarse el mismo.
, que a su vez es el peso que aumentamos al hidrolizar, ya
que al romper los enlaces les aportamos la molécula de agua que habían perdido
al formarse el mismo.
Lo máximo que podríamos tener sería 75 + 1,1 , que sería el 100%
![]() 75
+ 1,1 100%
75
+ 1,1 100%
75 x% x = 90,9%
2.- Por metilación exhaustiva de 32,4 gr de
amilopectina seguida de hidrolización ácida se obtienen 10 ![]() de 2,3,4,6
tetrametil-glucosa.
de 2,3,4,6
tetrametil-glucosa.
sCuáles son los otros productos (derivados metilados) que espera obtener?
sCuánto de cada uno?
sQué tanto % de residuos de glucosa están unidos por enlaces (1
Si el peso molecular de la amilopectina es
de ![]() , sCuántas ramificaciones posee una molécula de amilopectina?
, sCuántas ramificaciones posee una molécula de amilopectina?
Amilopectina = glucosas con enlace (1 ) + ramificaciones periódicas con es.(1
Hay cuatro tipos de Glc distintas: - La del C anomérico libre (una en cada molécula de amilopectina)
- Las unidas por e(1 4) (de la cadena principal)
- las que forman los puntos de ramificación, con es.(1
-
las de los extremos no reductores (OH del ![]() libre).
libre).
La 2,3,4,6 tetrametil-glucosa
corresponderá a la del extremo no reductor, pues está unida a la cadena por el ![]() y tiene libres los C
2,3,4 y 6.
y tiene libres los C
2,3,4 y 6.
Por cada extremo no reductor tenemos una ramificación y podemos considerar que hay igual cantidad de puntos de ramificación que extremos no reductores. Estos puntos de ramificación tienen libres los C 2 y 3, y por tanto obtendremos 2,3 dimetil-glucosa.
Las glucosas de las cadenas tienen libres los C 2,3 y 6 y nos darán 2,3,6 trimetil-glucosa.
Nos queda la glucosa del extremo reductor, que nos dará 2,3,6 trimetil-glucosa, y no del 1, porque se elimina en la hidrólisis ácida.
Para calcular que % de cada uno haremos lo siguiente: como tenemos un total de 32,4gr de amilopectina,
![]()
![]() como los extremos no reductores hemos visto que nos darían 2,3,4,6
tetrametil-glc y hemos obtenido 10
como los extremos no reductores hemos visto que nos darían 2,3,4,6
tetrametil-glc y hemos obtenido 10![]() ,
,
200 100% x = 5% de extremos no reductores, y
10 x% consecuentemete lo mismo de puntos
de ramificación.
Partiendo de esto, y sabiendo que el
número de glucosas que tenemos es ![]() , el 5% de estas es 3700 ramificaciones por molécula de
amilopectina.
, el 5% de estas es 3700 ramificaciones por molécula de
amilopectina.
También hay otro 5 % de extremos.
El % de enlaces (1 4) es el otro 90% .
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 939
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved