| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
DOCUMENTE SIMILARE |
|
PÉPTIDOS Y PROTEÍNAS
El enlace peptídico, que une aminoácidos para formar
péptidos y proteínas, químicamente es un enlace de tipo amida, que se
forma por condensación entre el grupo ![]() carboxilo y
carboxilo y ![]() amino de otro con liberación de una molécula de
amino de otro con liberación de una molécula de ![]() . Lo que se forma es un dipéptido.
. Lo que se forma es un dipéptido.
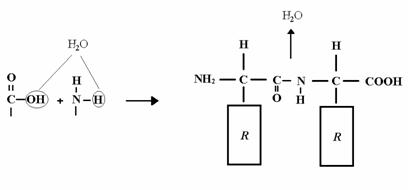
Con otro aminoácido más tendríamos un tripéptido, y así sucesivamente (tetrapéptido).
Los péptidos se nombran empezando por el aminoácido con el grupo amino libre y acabando con el que tiene el grupo carboxilo libre (terminal). A los aminoácidos se les coloca el sufijo “–IL” excepto al último. Si son largos, se escribe la secuencia con las abreviaturas o bien con nombres propios.
El enlace peptídico tiene unas características muy importantes porque de ellas dependen las características del péptido y las proteínas.
En primer lugar, es un enlace muy resistente a la hidrólisis, con lo que las proteínas son muy estables.
Tanto el enlace C-O como el C-N tienen carácter parcial de doble enlace. Esto impide la libre rotación de la molécula alrededor del enlace peptídico, y como resultado, los 6 átomos que participan en el enlace son “coplanarios”, de tal manera que como las proteínas se pliegan para formar la estructura secundaria, lo que gira son los planos sucesivos de los enlaces peptídicos.
Partiendo de los 20 aminoácidos proteicos, el número de péptidos posibles es enorme, de echo, la variedad existente es muy grande.
Dentro de los péptidos de importancia
biológica destacaremos en primer lugar el “glutatión”, un tripéptido que es el ![]() - glutamil – cisteinil – glicina.
- glutamil – cisteinil – glicina.
Es muy importante porque actúa como protector de las condiciones reductoras en el citosol celular, ya que la cisteína posee un grupo SH y el glutatión puede formar dímeros por puentes disulfuro, pasando de la forma oxidada a la reducida.
Es uno de los péptidos más pequeños con interés fisiológico.
Las encefalinas son más grandes. Hay la Met-encefalina y Leu-encefalina, llamadas así porque se diferencian solo en el aminoácido carboxilo-terminal.
Son pentapéptidos (5) y se encuentran en el cerebro donde intervienen en procesos relacionados con la percepción del dolor.
También tenemos hormonas como la vasopresina y oxitocina, nonapéptidos (9). La primera regula la reabsorción renal de agua y la segunda la secreción de leche.
De mayor tamaño tenemos oligopéptidos de hasta 34 aminoácidos con función hormonal.
h Un polímero de aminoácidos deja de ser un péptido para pasar a ser considerado proteína según dos criterios:
en primer lugar, la longitud (hasta 50 aá. Es un péptido) esto es bastante ambiguo, debido a que por ejemplo, la hormona paratiroidea, con 84 aminoácidos suele considerarse un péptido.
En segundo lugar, y más importante, la existencia biológica.
La cadena de aminoácidos que se sintetiza en el laboratorio y cuya existencia no ha sido demostrada en un organismo no será proteína por muy larga que sea. Si existe en la naturaleza, ello dependerá, ahora sí, de su longitud. Aún así, el límite es difuso.
Clasificación de las proteínas
Las proteínas se pueden clasificar según varios criterios, y todos son complementarios. Nos centraremos en 4 criterios:
Composición
Forma
Solubilidad
Función
- Según la composición, estas pueden ser simples o conjugadas. Las simples están constituidas solo por aminoácidos y las conjugadas contienen algún otro componente de diferente naturaleza química.
Cuando este componente es un glúcido, son glicoproteínas.
Si es un lípido, lipoproteínas.
Si es un ácido nucleico, nucleoproteínas
otra variedad son los grupos prostéticos como el grupo “hemo”, que da hemoproteínas, o un metal, que da metaloproteínas
Según su forma, clasificaremos a las proteínas en globulares y fibrosas. Las primeras tienen forma esférica u ovoide, y suelen ser solubles, mientras que las segundas son alargadas e insolubles, constituyendo la base de los tejidos estructurales, por lo que también se las denomina escleroproteínas.
Como las globulares son solubles y las fibrosas no, podemos decir que la siguiente clasificación está relacionada con esta y puede considerarse como una subdivisión.
Según la solubilidad, diferenciamos entre:
![]() Albúminas, que son solubles en agua fría. Estas 2 tienen
Albúminas, que son solubles en agua fría. Estas 2 tienen
Globlulinas, que son solubles en disoluciones salinas diluídas origen animal
Glutelinas, que son de origen vegetal, insolubles en agua y disoluciones salinas, pero sí en ácidos y bases diluidos.
Prolaminas, son de origen vegetal, y solubles en alcoholes de bajo peso molecular.
Protaminas, proteínas básicas que
se encuentran en los fluidos seminales y son solubles en agua y ![]() diluído.
diluído.
Histonas, que interaccionan con el DNA en los núcleos y son básicas, solubles en agua y ácidos diluídos.
Dado que este grupo es de proteínas solubles, todas ellas serán, pues, globulares.
Según su función (esta clasificación es la más heterogénea y la más informativa), pueden ser:
Encimas, que catalizan las reacciones biológicas
Transportadoras, como la
hemoglobina (de ![]() ) o proteínas de membrana (de metabolitos)
) o proteínas de membrana (de metabolitos)
De reserva, como almacén de energía, como la ovoalbúmina.
Contráctiles, como la actina y miosina, que intervienen en la contracción muscular
Estructurales, que son las más abundantes, de entre las que destaca el colágeno.
De defensa
Hormonal
Como factores tróficos. Se desempeña con el desarrollo de tejidos a nivel embrionario.
Receptores, de entre los que destacan los hormonales, proteínas de membrana a las cuales se une una hormona. Esta unión desencadena una acción hormonal.
Proteínas cromosómicas como las histonas
Toxinas, que abundan en microorganismos, insectos y reptiles
Estructura de las proteínas
Las proteínas son macromoléculas, en general bastante grandes (5000 aá. pequeña), con una estructura tridimensional muy compleja cuya determinación es una tarea difícil. Existen métodos como la cristalografía por rayos X, y en la actualidad se conoce menos del 10% de las estructuras de las proteínas aisladas de organismos.
Las posibilidades que tienen los aminoácidos de plegarse son grandes, pero cada proteína adopta una estructura espacial única, que se conoce como estructura nativa, y que está directamente relacionada con la función biológica de esta, de manera que si se altera esta estructura nativa, se ve alterada su función.
Según las relaciones entre estructura y función, tenemos 3 tipos:
proteínas homólogas
isólogas
análogas
Las homólogas, perteneciendo a diferentes especies, presentan similar función y similar estructura.
Las isólogas, van a ser originarias de la misma especie, desempeñan una función similar, pero tienen diferencias estructurales que las hacen más adecuadas a funciones específicas de esos organismos.
Las análogas también son originarias de la misma especie, su estructura es similar pero con diferencias en su función. En este caso, se cree que sus genes derivan de un gen ancestral común por duplicación y mutación.
El estudio estructural se realiza estableciendo 4 niveles que son las estructuras primaria, secundaria, terciaria y cuaternaria. Seguiremos estos cuatro niveles.
La estructura primaria
Es la secuencia de aminoácidos. Nos indica el orden en el que están unidos los aminoácidos en la cadena polipeptídica.
En las proteínas conjugadas, nos indica también los lugares de unión con glúcidos o diferentes grupos prostéticos.
En la estructura primaria están las claves o información básica que determina las estructuras superiores, pero de momento no podemos predecir la estructura secundaria de las proteínas a partir de la primaria, nada más que e regiones pequeñas de estas proteíanas.
Esquemáticamente,
la estructura primaria puede definirse como largas cadenas zigzagueantes de
enlaces peptídicos y de carbonos![]() alternantes, estando las cadenas laterales de los
aminoácidos a los carbonos
alternantes, estando las cadenas laterales de los
aminoácidos a los carbonos![]() y denominándose residuos aminoacídicos.
y denominándose residuos aminoacídicos.
Existen varias técnicas diferentes para determinar la estructura primaria de la proteínas, una clásica, que es el “método de secuenciación de Edman”, que consiste en:
Primero tenemos que disponer de la proteína purificada, y si está formada por diferentes cadenas polipeptídicas enrolladas, separarlas.
Luego estas las rompemos en péptidos de 20 aminoácidos.
Seguidamente separamos estos péptidos cortos y los hacemos reaccionar con el fenilisotiocianato, que se une al aminoácido con posición aminoterminal específicamente, y da lugar a que el enlace que une este aminoácido con el siguiente, sea más sensible a hidrólisis, con lo que luego se somete a una suave, y solo se romperá este enlace peptídico.
Obtenemos el derivado del aminoácido (aá. + FITC) y un péptido con un aminoácido menos, con el que podemos realizar un segundo ciclo de secuenciación.
Los aminoácidos que separamos los podemos determinar con técnicas como la cromatografía.
Estructura secundaria:
Nos define las disposiciones regionales y con elementos de simetría que nos podemos encontrar en determinadas regiones en las cadenas polipeptídicas.
Las posibilidades son enormes, sin embargo existe una restricción muy importante a todas estas posibilidades, que consiste en la planaridad del enlace peptídico, de tal manera que los parámetros que cambian en la cadena son los ángulos de giro entre planos consecutivos de este péptido. Los valores de estos ángulos nos definen las estructuras secundarias.
Existen muchas
estructuras secundarias, pero entre las más estables se encuentran la hélice ![]() , la lámina
, la lámina ![]() (o hoja plegada), y
son las más estudiadas.
(o hoja plegada), y
son las más estudiadas.
Hélice ![]() :
:
En esta disposición la cadena polipeptídica se enrolla sobre sí misma como la rosca de un sacacorchos. La rotación es hacia la derecha, y las cadenas laterales de lo aminoácidos se disponen hacia el exterior por el volumen que ocupan.
Cada aminoácido está girado 100s con respecto al anterior, lo que quiere decir que hay 3,6 residuos de aminoácido cada vuelta completa.
Cada cuatro aminoácidos los enlaces peptídicos se encuentran cercanos, pero en niveles diferentes de la hélice, por lo tanto se pueden establecer puentes de hidrógeno entre estos enlaces ya que los grupos C-O y N-H tienen posición trans.
Estos puentes de
hidrógeno son la principal fuerza que estabiliza la estructura en hélice ![]() y son determinantes de
lo que se denomina la longitud del paso de rosca, que es de 54 nm en la hélice
y son determinantes de
lo que se denomina la longitud del paso de rosca, que es de 54 nm en la hélice![]() .
.
En la hélice ![]() los puentes de
hidrógeno se establecen entre el grupo C-O del aminoácido que ocupa la
posición “n” y el grupo ácido del aminoácido con posición n+4.
los puentes de
hidrógeno se establecen entre el grupo C-O del aminoácido que ocupa la
posición “n” y el grupo ácido del aminoácido con posición n+4.
Existen variantes
de esta estructura. Destacaremos 2 comunes estudiadas en las proteínas, la
hélice ![]() y la hélice
y la hélice ![]() .
.
En la hélice ![]() tenemos 3 residuos por
vuelta ya que los puentes de hidrógeno se establecen entre el grupo C-O
del aminoácido “n” y el N-H del aminoácido “n+3”.
tenemos 3 residuos por
vuelta ya que los puentes de hidrógeno se establecen entre el grupo C-O
del aminoácido “n” y el N-H del aminoácido “n+3”.
En la hélice ![]() , más ancha, nos encontramos con 4,4 residuos por vuelta;
aquí los puentes de hidrógeno se establecen entre el grupo C-O del
aminoácido “n” y el N-H del “n+5”.
, más ancha, nos encontramos con 4,4 residuos por vuelta;
aquí los puentes de hidrógeno se establecen entre el grupo C-O del
aminoácido “n” y el N-H del “n+5”.
Lámina ![]() o lámina plegada:
o lámina plegada:
Aquí la cadenas polipeptídicas se disponen manteniendo su estructura en zig-zag extendidas y en dos posibles disposiciones, la paralela, en la que las diferentes cadenas tienen el extremo amino y carboxilo del mismo lado, y antiparalela, con el grupo amino y carboxilo contrapuestos. Esta última, la antiparalela es más frecuente.
La lámina ![]() tiene una disposición
más compacta que la hélice
tiene una disposición
más compacta que la hélice![]() . En la lámina
. En la lámina![]() la estabilidad depende de puentes de hidrógeno. Aquí los
puentes se establecen entre el O de un grupo C-O y el H de
un N-H de la cadena paralela próxima, son puentes intercatenarios,
mientras que en la hélice
la estabilidad depende de puentes de hidrógeno. Aquí los
puentes se establecen entre el O de un grupo C-O y el H de
un N-H de la cadena paralela próxima, son puentes intercatenarios,
mientras que en la hélice ![]() eran intracatenarios.
eran intracatenarios.
Los residuos se
sitúan alternativamente hacia arriba o abajo dependiendo del plano definido por
la lámina![]() .
.
Los aminoácidos con cadenas laterales pequeñas son más compatibles con esta disposición, debido a que tienen menos impedimentos estéricos.
En el caso de las proteínas globulares puede suceder que las dos cadenas que interaccionan por puentes de hidrógeno pueden ser fragmentos de la misma cadena, por medio de un giro.
Estructura terciaria:
Define el conjunto de todas las interacciones tales como puentes de hidrógeno, enlaces disulfuro, enlaces de Van der Waals que rigen el plegamiento de la cadena completa en el espacio, y engloba todas las estructuras secundarias que pueden existir en distintas zonas de la molécula.
Además, define otras zonas como las de “ovillo estadístico”, zonas de giro o torsión, donde las cadenas se pliegan sobre sí mismas
Destacaremos los
giros ![]() , que son bastante bruscos, por el establecimiento de puentes
de hidrógeno entre los grupos C-O del aminoácido “n” y el grupo N-H
del aminoácido “n+3”, empleados para conectar láminas
, que son bastante bruscos, por el establecimiento de puentes
de hidrógeno entre los grupos C-O del aminoácido “n” y el grupo N-H
del aminoácido “n+3”, empleados para conectar láminas ![]() antiparalelas.
antiparalelas.
También existen estructuras al azar con plegamientos al azar.
En algunos casos la cadena polipeptídica puede tener varias zonas de plegamiento con elasticidad propia. A estas se les denomina “dominios”.
En proteínas
fibrosas las estructuras secundaria y terciaria coinciden, las cadenas
laterales se orientan todas igual. Un ejemplo es la ![]() -queratina del pelo, que sirvió para determinar las dimensiones
moleculares de las hélices
-queratina del pelo, que sirvió para determinar las dimensiones
moleculares de las hélices ![]()
Estructura cuaternaria:
Define las interacciones entre varias cadenas polipeptídicas para formar una proteína. Las proteínas que solo son una cadena, son monómeros, si tienen dos cadenas son dímeros, si tres, trímeros, tetrámeros.
Son homodímeros si las dos subunidades son iguales, y heterodímeros si son distintas. Las fuerzas que estabilizan la estructura cuaternaria son variadas y son las mismas que las que estabilizan la estructura terciaria (enlaces disulfuro, puentes de hidrógeno, interacciones iónicas).
Es frecuente que en las proteínas oligoméricas puedan existir varias estructuras cuaternarias, varios estados conformacionales que afecten a la actividad de la proteína, como en las encimas alostéricas o la hemoglobina.
EL COLÁGENO
Es la proteína fibrosa más abundante del cuerpo y posee una estructura secundaria característica que es la hélice P.
El colágeno es la proteína base del tejido conjuntivo y el constituyente orgánico básico de los tejidos calcificados.
La estructura primaria es única, se caracteriza por tener un 30% de glicinas, y alrededor de un 20% de prolina e hidroxiprolina. Estas dos son características del colágeno. Solo la elastina, que es otra proteína de tendones y cartílago también contiene estos dos amionoácidos en esta proporción.
La estructura
secundaria. La hélice P es más rígida y extendida que la hélice ![]() (con cada aminoácido ascendía 18 nm) y aquí 0,29 nm.
(con cada aminoácido ascendía 18 nm) y aquí 0,29 nm.
Esto se debe a:
la presencia de prolina e
hidroxiprolina, aminoácidos cíclicos con un anillo del cual forma parte el N![]() , que da lugar a que el anillo sea rígido estéricamente a la
cadena polipeptídica,
, que da lugar a que el anillo sea rígido estéricamente a la
cadena polipeptídica,
a que al estar el N![]() formando parte del anillo no puede formar puentes de
hidrógeno por no tener hidrógenos,
formando parte del anillo no puede formar puentes de
hidrógeno por no tener hidrógenos,
y a que los OH’s de la hidroxiprolina establecen estos puentes de hidrógeno que estabilizan a esta hélice P.
La hélice P es levógira, se enrolla formando una triple hélice dextrógira que forma el tropocolágeno, que presenta 2 nm de diámetro y 260 nm de longitud.
Uno de cada 3 aminoácidos son glicinas.
En el tropocolágeno uno de cada tres aminoácidos orienta su radical hacia adentro de la hélice, y solo la glicina puede hacer esto.
Otra característica del colágeno es que estas cadenas polipeptídicas pueden unirse por enlaces transversales con otras cadenas que se establecen entre los residuos de la lisina y el lisinal (derivado de la lisina).
Estos enlaces también están entre diferentes unidades de tropocolágeno que se asocian para formar las protofibrillas y fibrillas del colágeno. Esta unión también se establece entre enlaces de residuos de lisina y lisinal.
El colágeno es una
glucoproteína, y existen diferentes tipos de colágeno que se diferencian entre
sí por la composición de las cadenas polipeptídicas que los forman. Existen dos
tipos de cadena, la ![]() y la
y la ![]() , que se diferencian en la secuencia de aminoácidos, en las
glicosilaciones y en el número de OH’s de la prolina.
, que se diferencian en la secuencia de aminoácidos, en las
glicosilaciones y en el número de OH’s de la prolina.
Estas características de los distintos tipos de colágenos adaptan a los mismos para su función en el cuerpo.
Comportamiento de las proteínas en disolución
La mayor parte de las proteínas se encuentran disueltas en los líquidos biológicos, donde cumplen su función. Estos líquidos son disoluciones salinas acuosas.
Las propiedades derivan de dos características de las proteínas:
Su naturaleza poliiónica
Su naturaleza macromolecular
Con relación a su naturaleza poliiónica, tanto lo extremos de la cadena polipeptídica como los residuos de los aminoácidos ácidos y básicos contribuyen a que la proteína tenga cargas en su periferia, que se comporte como un poliión. Por este motivo hay contraiones orgánicos en la disolución que rodean la proteína por atracción electrostática y dejan su carga neutra.
Los aminoácidos
poseen sus constantes de ionización, pero al estar unidos a la proteína, la
presencia de otros grupos cercanos hace que se modifique, de manera que el pK
de los residuos varíe en ![]() unidad.
unidad.
Las proteínas, al igual también poseen un punto isoeléctrico (p.i.) en el que para ese pH su carga neta es cero.
Si pH < p.i. la proteína tiene carga positiva
Si pH > p.i. la proteína tiene carga negativa.
El punto isoeléctrico depende de la composición de aminoácidos de modo que si el contenido en aminoácidos básicos y ácidos es parecido, el p.i. se encuentra entre 6 y 8, si predominan los aminoácidos ácidos el p.i. será más bajo, como en el caso de la pepsina, cuyo p.i. es 1,5. gracias a esto, la pepsina puede resistir y desenvolverse por el ácido jugo gástrico e hidrolizar proteínas de tipo neutro
Las histonas son básicas, y pueden unirse al DNA, cargado negativamente.
El carácter poliiónico hace que la solubilidad dependa del pH, contenido salino del medio
Como norma general, una proteína es más soluble si las interacciones entre moléculas proteicas se reducen al mínimo.
Respecto al PH, la solubilidad es mínima en el p.i. ya que no existe repulsión entre las moléculas y no se solubilizan
Con respecto al contenido salino, el efecto es bifásico, a una cierta concentración de sales, estas actúan como contraiónes, mientras que a una concentración elevada insolubilizan las proteínas ya que los iones se rodean de una esfera de solbatación con moléculas de agua, y estas no están libres en la disolución.
Con respecto a la característica de ser macromoléculas, al serlo forman disoluciones coloidales, fundamentales para membranas semipermeables. No las pueden atravesar por su tamaño. Esto se usa también para diálisis.
Hacen también de receptores iónicos que atraen a pequeños iones.
Desnaturalización de proteínas
La desnaturalización es el proceso por el cual una proteína cambia su estructura nativa, lo que conlleva a la pérdida de su actividad biológica.
Es provocada por el calor, PH’s extremos o por agentes que rompen enlaces no covalentes como los detergentes o la urea.
Las estructuras afectadas son la terciaria y la cuaternaria. La proteína se despliega y los residuos hidrófobos que están dirigidos hacia el interior de la proteína, al desnaturalizarse afloran al exterior con la consiguiente isolubilización de la proteína.
Solo en los casos más extremos se ve afectada la estructura secundaria, pero nunca la primaria. Para romper los enlaces peptídicos hay que recurrir a tratamientos más fuertes o digestión con encimas específicas (proteasas) como la pepsina.
Plegamiento de las proteínas ( Cómo adoptan su estructura en 3D)
El mecanismo es muy complejo, en general está dirigido por la tendencia a alcanzar estados de mínima energía.
Una fuerza muy importante es la hidrofobia de los residuos de los aminoácidos, que dirige el plegamiento, de forma que los residuos polares se colocan hacia la periferia de la proteína en contacto con la solución acuosa, mientras que los residuos apolares o hidrófobos tienden a protegerse del agua colocándose en el interior de la proteína.
El plegamiento no es espontáneo a partir de cualquier conformación una vez que la proteína está formada, lo que explica que gran parte de las desnaturalizaciones sean irreversibles.
Cuando la proteína está siendo sintetizada, existen otras proteínas que se llaman “chaperoninas”, que colaboran en que se pliegue correctamente la cadena en crecimiento. Para ello, se unen a la cadena desplegada impidiendo interacciones no deseadas, en un proceso ligado a la hidrólisis del ATP.
En el plegamiento intervienen otras proteínas encimáticas como “isomerasas de puentes de sulfuro”, etc.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1306
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved