| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
ANTIGENELE, DEFINITIE, STRUCTURA,CLASIFICARE SI CONDITIILE ANTIGENITATII
Antigenele (Atg) sunt substante care induc un raspuns umoral sau celular.
1. STRUCTURA UNEI MOLECULE DE ANTIGEN
Din toata molecula de Atg., numai unele arii mici cu suprafata de aproximativ 7 m2 , situate obligatoriu la suprafata moleculei, ii confera specificitate antigenica.

Fig 11 Diagrama
ilustrand diferen]ia dintre antigene imunogene [i neimunogene.
Pe suprafata unei singure molecule poate exista un numar diferit de epitopi cu specificitati multiple, molecula nefiind de fapt, 'un antigen unic', ci un 'mozaic de antigene', cu un numar mai mare sau mai mic de epitopi. De exemplu, pe o molecula de ovalbumina care are greutatea moleculara de cca 40.000 da., exista 5 epitopi diferiti, iar pe una de tiroglobulina cu greutatea moleculara 70.000 da., exista 40 de epitopi. Epitopul are deci o functie haptenica simpla, reactioneaza specific cu anticorpul, dar singur nu poate declansa sinteza acestuia.(Fig. 11 )
Pentru aceasta, este necesara interventia unei componente care sa-l 'poarte' conferind complexului format o anumita dimensiune care-l face 'vizibil' pentru limfocite. Aceasta componenta moleculara se numeste 'grupare purtatoare' sau 'carrier' si este data de catre restul moleculei. Schematic, un imunogen este alcatuit dintr-o grupare purtatoare care-i confera
dimensiuni si o grupare determinanta sau epitop care-i confera specificitatea antigenica. (Fig.12).
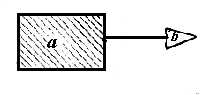
Fig.12. Partile componente ale unei molecule de antigen:
a) gruparea purtatoare (carrier);
b) determinantul antigenic (epitopul);
Gruparea purtatoare poate 'purta' un singur epitop sau un numar multiplu de epitopi cu una sau mai multe specificitati antigenice. Pentru fiecare dintre acesti epitopi, in populatia de celule limfoide, exista un corespondent reprezentat de un grup restrans (clona) de limfocite care exprima pe suprafata lor receptori unici si specifici.
2. CONDITIILE Atg
Antigenele poseda doua calitati definitorii: imunogenitatea si specificitatea.
A. Imunogenitatea depinde de:
A.1. Dimensiunea moleculei: cea mai mica molecula cunoscuta care este capabila sa induca raspuns imun (RI) este Vasopresina (gr. mol. 1000 Da). Se admite insa, ca factor general, ca o molecula poate avea proprietati antigenice daca greutatea sa moleculara este mai mare de 5000 Da. O astfel de molecula poate contine in structura sa mai multi epitopi identici sau diferiti. In functie de numarul si diversitatea epitopilor o macromolecula antigenica va reactiona cu mai multi anticorpi diferiti.
Numarul de molecule de anticorpi cu care poate reactiona o molecula de Atg. reprezinta 'valenta antigenului'. Exista insa molecule foarte mici care, prin forma lor primara, nu intrunesc conditia de marime care sa le confere proprietati imunogenice (haptene).
Ele vor deveni antigenice numai dupa ce se vor combina cu macromoleculele gazdei. Combinatiile respective se numesc 'haptenice'. Fata de aceste combinatii, organsimul raspunde prin formare de Atc impotriva moleculei mici. Moleculele - carrier confera numai proprietatea de antigenicitate haptenei prin prezentarea acesteia catre LfB sub forma multivalenta capabila sa le provoace activarea. 'Carausii' pot fi molecule solubile circulante (ex. serum albumina) sau molecule constitutive ale
membranei unei molecule (de ex. eritrocite, granulocite sau trombocite).
In anumite imprejurari, penicilina se poate fixa pe membrana eritrocitelor cu care formeaza complexe haptenice. Atc. anti - penicilina se vor fixa pe haptena si vor inveli eritrocitele. In trecerea acestor eritrocite prin splina, Mcf. splenice vor atasa anticorpii prin intermediul R.Fc de pe membrana lor (imunoaderenta si incercand sa elimine complexele imune Atc + P (fenomenul de 'clearance') vor fagocita totodata si eritrocitele purtatoare. Prin urmare, eritrocitele vor fi distruse prematur numai prin faptul ca au devenit purtatoare de haptene (procesul a fost denumit mecanismul 'spectatorului inocent' si este implicat in liza indusa de droguri pentru mai multe categorii de celule sanguine).
A.2. Calea de patrundere: o substanta care s-a dovedit antigenica, daca a fost administrata intradermic, poate fi non-imunogenica daca este introdusa intramuscular.
A.3. Asocierea Atg. cu un adjuvant intareste imunogenicitatea in anumite conditii experimentale. Adjuvantii ar produce o captare mai mare a Atg. de catre Mcf si o eliberare mai lenta combinata cu producerea unei reactii inflamatorii la locul injectarii. Adjuvantul cel mai utilizat experimental este o emulsie de ulei mineral, de bacili Koch omorati (adjuvantul Freund complet).
A.4. Natura chimica a Atg.: proteinele sunt mai antigenice ca polizaharidele care la randul lor sunt mai antigenice decat lipidele. Macromoleculele de ADN sunt foarte slab antigenice.
A.5. 'Repertoriul' imunologic al individului reprezinta ansamblul antigenelor pe care le poate identifica un individ dintr-o specie. Specii diferite sunt programate sa recunoasca anumite Atg. Aceasta programare este controlata genetic. Ex. Atg. Rh de pe suprafata Er. umane produc un raspuns imun energic la om, dar nu produc nici un fel de raspuns la iepure si cobai.
La om, repertoriul imunologic ar insuma aproximativ 108 posibilitati calculate.
B. Specificitatea
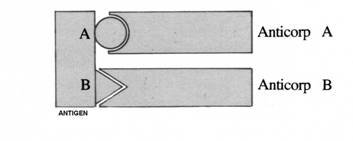
Un
Atg. este in principiul identificat de un Atc specific. Aceasta identificare
este realizata de receptorii pentru Atg.
de pe suprafata limfocitelor.
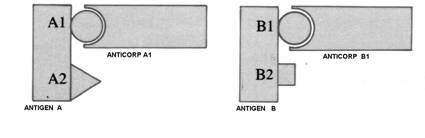
Fig. 13 Specificitatea antigenica
Fig.14. Reactii `ncrucisate
In cazuri rare Atg. diferite pot avea epitopi similari sau identici dand nastere la producerea unor Atc. 'crossreactivi'.
Legarea Atc. pe Atg. ('recunoasterea Atg.') este un proces fizic care nu difera de acela prin care se asociaza intre ele doua macromolecule chimic inrudite. Acest proces implica o potrivire reciproc complementara
intre configuratia spatiala a epitopului si cea a situsului combinativ al anticorpului ('paratopul') si o solidarizare a celor doua molecule partenere prin 4 tipuri de legaturi nespecifice: electrostatice, punti de hidrogen, legaturi hidrofobe si legaturi Van der Waals.(Fig. 13,14)
3. CLASIFICAREA ANTIGENELOR
Totalitatea antigenelor existente `n natura alcatuiesc un sistem foarte complex si divers, un adevarat univers antigenic, caruia sistemul imunitar al organismelor i se opune prin mecanisme adecvate de recunoastere, discriminare, neutralizare si eliminare, cu ajutorul efectorilor imuni, `n scopul mentinerii homeostaziei normale. Marea varietate a antigenelor face dificila o clasificare care sa corespunda tuturor cerintelor. Din ratiuni didactice, `n prezentarea diferitelor tipuri si categorii de antigene, adpotam urmatoarele criterii de clasificare:
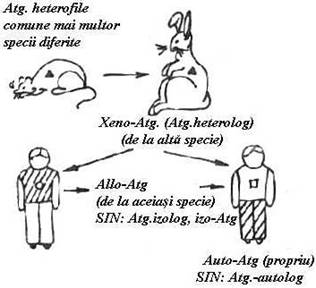
Dupa origine, Atg. pot fi clasificate in patru grupe (Fig.15):
1. Atg. heterofile - comune la mai multe specii de animale, plante si microorganisme.
In 1911 J. FORSSMAN a observat ca injectarea unor antigene de rinichi de cobai la un iepure duce la aparitia unor Atc. care vor actiona si cu Atg. rinichiului de cobai si cu Atg. ale eritrocitelor de oaie. Aceste tipuri de Atg. care induc aparitia unor Atc. care reactioneaza si cu Atg. speciei inductoare si cu Atg. altor specii au fost denumite 'heterofile'. Indiferent de sursa lor, Atg. heterofile au ca element comun o grupare heptenica similara, care determina activitatea lor imunologica comuna.
Ele sunt prezente pe eritrocitele unui mare numar de specii (oaie, capra, cal, caine, cobai) ca si la unele microorganisme, `nsa sunt absente la iepure si bou. La om sunt prezente numai la indivizii de grup sanguin A si AB. Antigenele heterofile de tip Forssman indiferent de originea lor, s-au dovedit a avea o grupare haptenica similara, care asigura reactivitatea lor imunologica comuna. Haptenele antigenelor heterofile au structura lipopolizaharidica, `nsa structura chimica a antigenului global; variaza de la o specie la alta si chiar de la un organ la altul.
Cercetarile ulterioare au demonstrat ca distributia antigenelor heterofile nu este uniforma. Astfel la unele specii ( de exemplu: oaia) sunt prezente atat `n tesuturi cat si pe eritrocite, pe cand la altele (de exemplu: cobai) se gasesc numai tesuturi.
Demonstrarea prezentei antigenelor Forssman se poate efectua prin inocularea de titrurat de rinichi de cobai de iepure. Se obtine astfel un ser ce contine anticorpi hemolizanti fata de hematiile de oaie (ser Forssman). Cercetarile au dovedit ca antigenul "rinichi de cobai" nu este identic cu antigenul "hematii de oaie". Din acest punct de vedere s-a precizat ca, un ser hemolitic antioaie este activ si pentru hematiile de bou, pe cata vreme serul Forssman este activ numai fata de hematiile de oaie. ~n consecinta se apreciaza ca hematia de oaie are o structura antigenica mai complexa, prezentand un antigen termolabil la 1000C, specific si izofil, si un antigen termostabil la 1000C, nespecific heterofil, care este de fapt haptena Forssman.
2. Atg. heterologe (xeno Atg.) = provin de la indivizi de alta specie.
3. Atg. izolage (allo Atg.) = provin de la indivizi de aceeasi specie si exprima polimorfismul genetic in cadrul unei specii. Doi indivizi ai unei specii sunt diferiti din punct de vedere antigenic.
4. Atg. autologe (auto Atg.) = Atg. exprimate de celulele proprii ale unui individ.
Dupa gradul de complexitate al moleculei de antigen se deosebesc:
- antigene complete - sunt considerate acele antigene care poseda atat suportul coloidal, cat si gruparile de specificitate (epitopii). Acest tip de antigene determina formarea de anticorpi si reactioneaza specific cu ei "in vitro". Cele mai multe din antigenele naturale sunt antigene complete;
- antigene incomplete (haptene) - sunt considerate ca fiind "fragmente" ale moleculei de antigen complet, care nu sunt capabile sa induca formarea de anticorpi specifici, atunci cand sunt inoculate singure la un organism primitor. Ele au `nsa capacitatea de a reactiona cu anticorpii, proces finalizat prin producerea unor reactii vizibile. Haptenele pot deci interactiona cu anticorpii specifici (poseda antigenitate), dar nu pot induce formarea lor (nu sunt imunogene). Haptenele sunt `n general compusi chimici cu greutate moleculara mica, ce se pot atasa de o molecula purtatoare, fiind restabilita astfel functia antigenica primara.
Dupa structura chimica se deosebesc urmatoarele tipuri
Antigene proteice - constituie grupa cea mai numeroasa de substante care `ndeplinesc de cele mai multe ori conditii de imunogenitate si antigenitate. Dupa intensitatea raspunsului imun se disting proteine cu imunogenitatea ridicata (exemplu: albuminele, glicoproteinele serice, etc) si proteine cu imunogenitate slaba (exemplu: hemoglobina si polipeptidele cu masa moleculara mica).
Tratarea termica sau chimica a unor proteine, poate modifica imunogenitatea lor. Este important de subliniat ca formaldehida aboleste activitatea biologica (patogena) a unor proteine, fara a afecta imunogenitatea lor. Aceasta proprietate este deosebit de importanta si are aplicabilitate practica la prepararea vaccinurilor. Dimpotriva tratamentul cu glutaraldehida confera moleculei (exemplu: albumina serica umana) noi determinanti antigenici constituiti probabil din nuclee piridinice, rezultate din reactia glutaraldehidei cu gruparile amino ale lizinei.
Antigene polizaharidice constituie o grupa importanta de substante `ntalnite `n natura, dar care sunt dotate cu proprietati imunogene mai putin marcante decat proteinele. Cu toate acestea, numeroase substante polizaharidice sunt active `n cadrul unor compusi complecsi. Trasatura caracteristica a celor mai multe polizaharide este structura lor regulata repetitiva, `n care unitati monozaharidice sunt legate `n lanturi lungi lineare sau ramnificate. Atat compusii polizaharidici lineari cat si cei ramnificati, au proprietati antigenice evidente, desigur cu modificarile corespunzatoare de specificitate determinate de structura. O serie de polizaharide prezente `n structura bacteriilor, au un rol `n determinarea specificitatii raspunsului imun fata de acestea. Asa este cazul compusilor glucidici ai pneumococilor, salmonelelor, colibacililor, etc. Antigenele de pe suprafata eritrocitelor, care confera specificitatea de grup sangvin (O, A, B, AB) sunt de natura glicoproteica, specifica.
Substantele glucidice au proprietati imunogene care se manifesta si prin inducerea si declansarea starilor de hipersensibilizare mediate umoral (prin anticorpi). Astfel de fenomene se realizeaza prin unele polizaharide ca atare, cat si prin haptenele lor glucidice, componente ale unor antigene. Starile de hipersensibilizare mediate celular, pot fi induse si declansate numai de catre haptene polizaharidice cuplate pe macromoleculele proteice purtatoare. Polizaharidele purificate au putere imunogena slaba, unele dintre ele (acidul hialuronic) fiind neimunogene sau imunogene doar pentru unele specii de animale (exemplu: polizaharidul de tip III din capsula pneumococului, este imunogen pentru soarece si om, dar nu si pentru cobai si iepure);
Antigene nucleice - reprezentate de catre acizii nucleici, ADN si ARN, care desi sunt macromolecule au slabe proprietati imunogene. Pot interveni `n unele procese imunologice, dar mai frecvent ca haptene, decat sub forma unor antigene complete. Pot induce formarea de anticorpi, atunci cand formeaza complexe cu unele polizaharide sau proteine. ~ncercarile de a demonstra imunogenitatea acizilor nucleici liberi, au dat rezultate contradictorii. Imunizarea cu ADN a indus raspunsuri mai constante `n urma asocierii sale cu adjuvant Freund. Dovada imunogenitatii lor este data de existenta anticorpilor anti-ADN mono sau dublu catenar, `n lupusul eritematos sistemic( uman sau murin) sau `n alte boli autoimune.
Antigenele lipidice - care includ diferite lipide singure sau `n asociere cu alte substante. Lipidele ca atare sunt lipsite de proprietati imunogene, dar pot avea calitati de haptene, mai ales daca se cupleaza cu proteine. Astfel, se pot obtine anticorpi anticolesterol, antilecitina, anticefalina, etc. Mai bine studiate sunt proprietatile antigenice ale cardiolipinei, o haptena folosita `n reactia Wasserman, reactie utilizata `n diagnosticul serologic al sifilisului la om, cardiolipina (raspandita `n tesuturile animalelor, plantelor si chiar la unele bacterii), reactioneaza pozitiv numai cu anticorpii oamenilor infectati cu Treptonema pallidum. Lipidele confera proprietati antigenice si functionale lipopolizaharidelor, endotoxinelor si altor complexe antigenice.
Dupa modul de formare, antigenele se clasifica `n naturale, artificiale si sintetice:
antigene naturale - reprezentate de diferiti constituienti naturali, `ntalniti la animale, plante si microorganisme. Pot fi substante solubile (proteine, polizaharide, acizi nucleici) sau diferite particule insolubile, corpusculare (celule, bacterii, virusuri, paraziti).
antigene artificiale - formate din macromolecule naturale cu rol de purtator, pe care se leaga `n conditii artificiale diferite haptene, care vor imprima caracterul de specificitate.
antigenele sintetice - `ntalnite `n conditii de laborator si care sunt produse prin diferite procedee de sinteza. Sunt reprezentate de polipeptide sintetizate `n mod artificial, prin polimerizarea unor derivati ai L sau/si D- aminoacizilor (anhidridele N-carboxiaminoacizilor).
Dupa capacitatea de a stimula limfocitele T sau B, antigenele se clasifica `n antigene T-independente si antigene T-dependente.
antigene T-independente (timus independente) sunt antigene capabile sa induca maturarea limfocitelor B purtatoare de receptori pentru antigen, fara a necesita interventia limfocitelor.
antigene T-dependente (timus dependente) sunt antigene care induc formarea de anticorpi de catre descendentii celulelor B, purtatoare de receptori pentru antigen, numai prin cooperare cu limfocitul T ajutator (Th), fiind necesar existenta unor stimuli aditionali.
4. PRINCIPALELE CATEGORII DE ANTIGENE
NATURALE
4.1. Antigenele bacteriene
O bacterie nu este unica de punct de vedere antigenic, ea fiind de fapt constituita dintr-un mozaic de antigene. Acestea sunt reprezentate de diferite componente chimice prezente `n structuri bacteriene. Astfel se disting:
- Antigene somatice "O". Sunt antigene extrase din corpul bacteriilor, avand localizarea preponderenta `n peretele celular. Ele sunt de natura polizaharidica, sunt termostabile si poseda `nsusiri biologice complexe:
a. sunt imunogene, asigurand specificitatea de specie la bacteriile Gram-negative si de grup la cele Gram pozitive;
b. dezvolta hipersensibilitatea imediata si de tip Sanarelli-Scwartzmann;
c. stimuleaza sistemul mononuclear fagocitar si imunitatea nespecifica;
d. manifesta activitate anticomplementara.
La bacteriile Gram negative antigenul somatic este reprezentat de endotoxine, iar la bacteriile Gram pozitive de catre acizii lipoteichoici.
- Antigene flagelare "H". Abrevierea H provine de la cuvantul hauch care `nseamna val. Sunt obtinute din flagelii (cilii) celulelor bacteriene. Din punct de vedere chimic sunt de natura proteica, iar imunologic stimuleaza mai ales limfocitele B. Antigenele flagelare au fost bine studiate la bacteriile din familia Enterobacteriaceae, `ndeosebi la cele din genul Salmonella, avand un rol important `n identificarea (tipizarea) serologica a acestor bacterii.
- Antigenele fimbriale "F". Se obtin din fimbriile bacteriilor. Prezinta specificitate corelata cu specia bacteriana, `nsa pot da reactii `ncrucisate. Mai bine studiate sunt antigenele F la E.coli si anume F4 (K88) prezente la tulpinile ce `mbolnavesc purceii si F5 (K99) `ntalnite la tulpinile patogene pentru vitei. Aceste antigene au o importanta deosebita `n mecanismul patogenic al colibacilozei, favorizand fixarea colibacililor la suprafata eritrocitelor.
- Antigenele de `nvelis "K". Sunt prezente `n structura capsulara a bacteriilor capsulogene. Din punct de vedere chimic, capsula prezinta diferente de la o specie bacteriana la alta sau chiar `n cadrul aceleasi specii. De exemplu la Streptococcus pneumonie, este de natura polizaharidica si se disting 83 de tipuri antigenice; la Bacillus antharcis are o structura polipeptidica, iar la Klebsiella pneumoniae este de natura poliglucidica si se dinsting 83 de tipuri capsulare. Antigenele capsulare au rol mai ales `n mecanismul patogenic inhiband fagocitoza.
- Antigene de suprafata "Vi". Definesc capacitatea de virulenta a bacteriilor care poseda astfel de antigene. Ele sunt situate, `n celula bacteriana, exterior antigenului somatic, `mpiedicand aglutinarea cu seruri anti-O (inaglutinabilitate). Prezenta lor se coreleaza cu virulenta tulpinilor respective si au fost descrise la unele salmonele (S.tiphy, S.hirschfeldi, S. typhimurium) dar si la alte specii bacteriene (Citrobacter). Sunt antigene termolabile, dar insensibile la alcool si formol. Pentru a elimina fenomenul de inaglutinabilitate, tulpinile testate se supun `ncalzirii.
4.2. Antigenele virale
Capacitatea imunogena a virusurilor rezida `n structura lor chimica complexa. ~n general, componentele proteice ale capsidei si pericapsidei virale, `ndeplinesc conditii de antigenitate si imunogenitate, declansand raspunsuri imune satisfacatoare, pe cand acizii nucleici au o slaba capacitate imunogena.
Unele virusuri sunt unice din punct de vedere antigenic, fiind puternic imunogene (exemple: virusul IBR/IPV, virusul bolii lui Aujeszky, virusul pseudopestei aviare, etc), pretandu-se la prepararea unor vaccinuri eficiente. Alte virusuri prezinta un accentuat fenomen de variatia antigenica, caracterizat printr-o mare labilitate antigenica si mai ales imunogenetica, care duce la aparitia de noi variante antigenice, chiar pe parcursul aceleasi epidemii (exemple: virusul febrei aftoase prezinta 7 tipuri cu urmatoarele subtipuri sau variante: A-32, O-11; C-5, SAT1-7, SAT2-3. SAT3-4 si Asia1-3; virusul pestei porcine africane prezinta 20 variante imunologic distincte; virusul exantemului veziculos al porcului are 11 variante, etc.)
Aspectele particulare sub raportul variatiei antigenice se `ntalnesc la virusurile din familia Orthomyxoviridae (gripale). Acestea posedand un genom segmentat, exista posibilitatea aparitiei unor "rearanjari" a segmentelor genomice provenite de la virusuri diferite, cat si `ntre virusurile gripale umane, cu cele equine, porcine si aviare.
La virusul anemiei infectioase se descrie asa numita "deriva antigenica" determinata de modificarea antigenitatii anvelopei virale sub actiunea anticorpilor formati `n organism, astfel, ca noii progeni aparuti `n procesul de replicare se deosebesc antigenic de susele parentale.
Variatia antigenica creaza dificultati `n obtinerea unor vaccinuri eficiente, ca urmare a faptului ca prin schimbarea periodica a profilului antigenic se evita neutralizarea prin intermediul anticorpilor produsi fata de susele parentale.
4.3. Antigenele parazitare
Parazitii prezinta de asemenea o structura antigenica complexa. Ei reusesc sa se mentina `n organism prin intermediul unor organite de fixare sau acoperirea cu substante prin care se evita raspunsul imun din partea organismului. La o serie de paraziti, cum ar fi plasmodiul malariei, tripanosomele si boreliile, ca urmare a mutatiilor genice spontane cu implicatii asupra structurii lor antigenice, pot supravietui raspunsului imun dezvoltat de gazda, fata de populatia parazitara initiala, care poseda o structura antigenica diferita de cea a mutantilor lor. Antigenele pot fi solubile sau particulate.
~n infectiile cu paraziti intracelulari, imunitatea celulara este necesara pentru a conferi rezistenta fata de agresiunea parazitara, pe cand `n infestatiile cu paraziti extracelulari (nematozi, trematozi, etc) ramane o problema deschisa daca se dezvolta sau nu imunitatea protectiva fata de acesti paraziti. Se cunoaste ca `n organismul parazitat cu diversi helminti sau alte categorii de paraziti, se dezvolta anticorpi de tip reagenic (IgE) si stari de hipersensibilizare imediata. Acesti anticorpi sunt produsi de stimulare cu antigene, fie somatice, fie produse de metabolism ale parazitilor.
~n principiu este acceptat faptul ca, parazitii care `n ciclul lor evolutiv dezvolta larve ce invadeaza tesuturile gazdei, determina un raspuns imun, iar cei cu localizare intestinala nu determina raspuns imun protectiv.
Din parazitii adulti sau diferite stadii larvare, au fost obtinute antigene cu utilizare `n reactii serologice (hemaglutinare pasiva, RFC, seroprecipitare, ELISA, imunofluorescenta, etc) sau alergice. Astfel de antigene au fost obtinute din Cystricercus celulose, Cysticercus bovis, Anaplasma sp., Trichinella spiralis, Toxoplasma gondii, Tripanosoma equioerdum, Hipoderma bovis, etc .
4.4. Antigenele de grup sanguin si Rh
Conform sistemului antigenic A,B,O, oamenii pot fi clasati `n 4 grupe disticte si anume:
grupa I sau 0, cu aglutinine dar fara aglutinogeni;
grupa II sau A, cu aglutinogen A si aglutinine beta;
grupa III sau B, cu aglutinogen B si aglutinine alfa;
grupa IV sau AB, cu aglutinogeni AB dar fara aglutinine;
Transfuzia interindividuala este posibila numai `n cadrul aceleasi grupe, cu exceptia grupei 0, care nu ridica asemenea probleme. ~n caz contrat operatia de transfuzie poate fi letala pentru receptor, deoarece aglutininenele proprii vor aglutina hematiile transfuzate, aglutinare urmata de sfarsit letal datorita obstruarii vaselor sangvine. Aglutinogenii sistemului A, B si 0 se transmit ereditar, fiind sub controlul genelor A si B cu 3 alele (A,BO), 6 genotipuri (AA, AO, BB, BO, OO, AB) si 4 fenotipuri (A, B, O, AB). Grupa O este `ntotdeauna hemozigota (OO) iar grupa IV este heterozigota (AB).
Atg. de grup sangvin sunt determinate genetic.
Exemple
1. Atg. de grup sanguin din sistemul ABO sunt produse la comanda unor gene plasate pe cromozomul 9. Ele sunt exprimate atat la suprafata eritrocitelor cat si pe alte celule tisulare. Uneori pot exista si in lichide biologice (de ex., in saliva) la indivizii denumiti 'secretori'.
2. Atg. de grup din sistemul Rh sunt produse la comanda unor gene plasate pe cromozomul 1. Ele sunt exprimate numai pe eritrocite.
Sistemul RH a fost semnalat de catre Lansteiner si Wiener (1940), care au constatat ca serul iepurilor imunizati cu eritocitele maimutelor Macaccus rhesus aglutineaza eritrocitele a 85% din oameni. Acestia au fost numiti Rh pozitivi, ceilalti fiind Rh-negativi. Folosirea de antiseruri, obtinute prin izoimunizare a dus la identificarea a 26 de antigene Rh, deci un adevarat sistem. Cel mai frecvent se `ntalnesc urmatoarele antigene: D,d, C,c, E,e,. Grupul genic ce coordoneaza sinteza antigenelor Rh se transmite ereditar dupa legile mendeliene si este responsabil de eritroblastoza fetala. Astfel o mama Rh- care poarta un fetus Rh+ (caracter mostenit de la tata), va fi imunizata de hematiile fetale. Anticorpii aparuti produc aglutinarea eritrocitelor fetale si moartea produsului de conceptie. Spre deosebire de aglutininele alfa si beta ale sistemului A, B, O `n sistemul Rh, nu exista anticorpi naturali, aparand numai dupa imunizarea mamei de catre hematiile fetale sau dupa transfuzii cu eritrocitele subiectilor D (Rh+), sau de la cei D (Rhv).
Antigenele ce reprezinta grupele sanguine sunt constituite din molecule membranare prezente la suprafata celulelor sanguine circulante, unele dintre ele fiind puternic imunogene. Toate antigenele grupelor sanguine stimuleaza producerea de anticorpi specifici. Ele se identifica `n laborator printr-o reactie Ag-Ac cu ajutorul anticorpilor specifici. Este vorba de o reactie de aglutinare, dar adesea pentru a deveni vizibila se impune uneori efectuarea unor "artificii", cum ar fi mediul macromolecular, adaugarea unor enzime, aplicarea testului Coombs.
3. Atg. leucocitare (Human Leukocyte Antigens HLA) sunt produse de gene grupate intr-o zona a bratului scurt al cromozomului 6 denumita 'complexul major de histocompatibilitate' (CMH).
4.5. Antigenele de histocompatibilitate
Antigenele de histocompatibilitate (AHC) sunt reprezentate de structuri moleculare situate la suprafata tuturor celulelor unui organism. Ele nu difera de la un organ la altul sau de un tesut la altul ci difera de un individ la altul, `n functie de linia genetica din care provine. AHC s-au dovedit a fi controlate de gene de histocompatibilitate situate pe cromozomi.
CMH contine mai multi loci care la randul lor codifica trei familii de proteine. Locii A,B,C, codifica proteinele care sunt exprimate pe suprafata tuturor celulelor nucleate si a trombocitelor. Aceste proteine (A,B, C) sunt denumite 'proteine din clasa I'. Regiunea D cuprinde locii DP, DQ si DR. Produsele acestor gene au fost denumite 'proteine din clasa a II-a'.
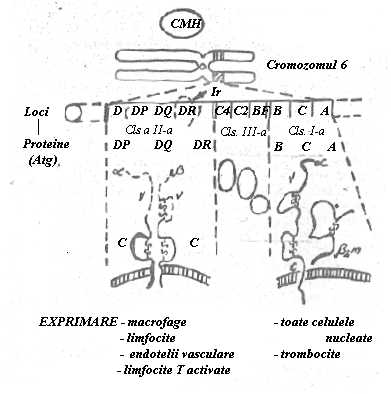
Fig.16 Complexul major de histocompatibilitate si antigenele HLA
Sunt exprimate de un numar mai restrans de celule si anume cele care sunt interesate functional in transmiterea informatiei antigenice: macrofagele care prezinta Atg. si din partenerele lor care recepteaza aceasta informatie: limfocitele B si limfocitele T activate. Proteinele din clasa I si a II-a formeaza sistemul antigenelor HLA. Pe langa genele ce codifica sistemul HLA, CMH mai cuprinde alti trei loci care controleaza producerea unor componente din sistemul complement C2, C4 si Bf. Aceste proteine foarte slab antigenice au fost grupate sub numele de proteine de clasa a III-a .(Fig.16).
4.6. Antigenele sechestrate
~nca prin cercetarile lui Burnet si ale lui Medawar si col. s-a dovedit ca exista o toleranta imuna `nnascuta fata de toate antigenele de organe, care au venit `n contact cu sistemul limfoid. Pentru structurile tisulare care raman topografic `n afara contactului cu limfocitele circulante, nu exista o toleranta imunologica `nnascuta. Asemenea structuri se gasesc `n anumite tesuturi, cum ar fi tiroida, creierul, testiculul si unele componente oculare si care `n conditii fiziologice nu sunt vehiculate `n organism.
~n cazuri patologice are loc deversarea lor `n circulatie, iar `n contact cu celulele imunocompetente, acestea le recunosc drept non-self si vor produce `mportiva lor efectori imun, avand drept consecinta inducerea de boli autoimune. Se aminteste `n acest sens tiroidita autoimuna, oftalmia simpatica, aspermia autoimuna, etc.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 6094
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved