| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Anatomia si fiziologia rinichilor
Anatomia rinichiului
Localizare. Rinichii sunt organe pereche situate retroperitoneal de o parte si de alta a coloanei vertebrale lombare. Polul superior al fiecarui rinichi corespunde vertebrei T12 iar polul inferior celei de a treia vertebre lombare - L3 [1].
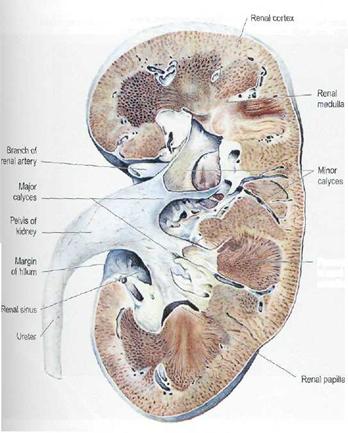
Aspect exterior. Forma rinichiului este comparata
de obicei cu cea a unui bob de fasole. Fiecarui rinichi i se disting : o fata
anterioara, usor convexa, o fata posterioara plana, un pol superior, un pol
inferior, o margine laterala convexa si o margine mediala concava unde se
gaseste hilul renal.[2] In hilul renal se gasesc artera si vena
renala, limfaticele, plexul nervos si pelvisul renal. La adult rinichiul
cantareste intre 125-
Rinichiul este alcatuit din parenchim si sistem colector.
Parenchimul renal. La nivelul parenchimului renal se delimiteaza doua zone: o zona corticala externa si o zona medulara interna.
Zona
corticala externa (cortexul renal) se
intinde de la capsula pana la bazele piramidelor Malpighi [³]. Are culoare rosiatica,
aspect granulat si o grosime de 10-
Zona medulara se intinde de la baza piramidelor Malpighi pana la papile. Fiecare rinichi are 10-18 piramide conice (piramide Malpighi) cu aspect striat. Piramidele prezinta o baza orientata spre marginea laterala si un varf - papila renala convergent in sinusul renal. Fiecare piramida Malpighi contine in jur de 500 piramide Ferrein - prelungiri radiare medulare cu varful spre exterior [²].
Sistematizarea rinichiului. Rinichiul poate fi divizat in lobi respectiv lobuli. Lobul renal reprezinta teritoriul centrat de o piramida Malpighi, impreuna cu toate piramidele Ferrein dependente de ea, limitat lateral printr-o linie ce trece prin mijlocul unei coloane Bertin. Lobulul renal reprezinta teritoriul centrat de o piramida Ferrein si apartine numai corticalei.
Sistemul colector cuprinde calicele mici, calicele mari si bazinetul. Calicele mici se insera cu o extremitate pe circumferinta papilelor renale si se unesc cu cealalta extremitate intre ele pentru a forma calicele mari. Fiecare papila proemina in interiorul unui calice mic si prezinta 15-20 perforatii ce formeaza aria cribrosa.[3] 8-10 calice mici formeaza 2-3 calice mari. Calicele mari se unesc si formeaza pelvisul renal - bazinetul- o palnie musculo-membranoasa cu baza orientata catre calicele mari si varful catre ureter.(Figura 1.1)
|
Figura 1.1. Sectiune longitudinala prin rinichi - adaptata dupa Gray`s Anatomy
Microscopie. Nefronul reprezinta unitatea structurala si functionala a parenchimului renal. Fiecare rinichi uman contine aprox 1-1,3 milioane nefroni. Nefronul este alcatuit din - corpusculul renal Malpighi
- tubul renal.
Corpusculul renal Malpighi este format la randul sau din - glomerul
- capsula Bowman
- aparatul juxtaglomerular
Tubul renal este alcatuit din - tubul proximal
- ansa Henle
- tubul distal.
Fiecare nefron se continua cu un tub colector care dreneaza in canalul colector.
Glomerulul este alcatuit din o retea de anse capilare .Capilarele sangvine se dispun in jurul axelor mezangiale alcatuite din matricea mezangiala si celulele mezangiale.¹ Reteaua capilara glomerulara este situata intre doua arteriole : arteriola aferenta si arteriola eferenta realizand o retea admirabila. Glomerulului i se descriu un pol vascular locul unde patrunde arteriola aferenta si iese arteriola eferenta, respectiv un pol urinar care se deschide in tubul proximal [³].
Capsula Bowman inveleste ghemul capilar glomerular si este formata din doua foite, foita parietala si foita viscerala ce delimiteaza camera urinara (camera de filtrare), unde este colectat filtratul glomerular. Foita viscerala a capsulei Bowman inveleste capilarele glomerulare si este alcatuita dintr-un strat de celule epiteliale numite podocite. Podocitele prezinta prelungiri citoplasmatice de trei ordine 1, 2, 3. Prelungirile de ordinul 1 corespund proceselor majore, cele de ordinul 2 proceselor secundare, iar prelungirile de ordinul 3 corespund pedicelelor. Procesele pediculate au la suprafata lor un invelis incarcat negativ, bogat in acid sialic cu rol important in mentinerea structurii si a functiei normale a barierei de filtrare. Stratul celulelor podocitare este situat pe o membrana bazala care se uneste cu membrana bazala a anselor capilare. Foita parietala a capsulei Bowman este alcatuita dintr-un epiteliu pavimentos simplu, se sprijina pe o membrana bazala, la polul vascular se rasfrange si se continua cu foita viscerala, iar la polul urinar se continua cu epiteliul tubului proximal [²].
Membrana bazala glomerulara (MBG) este cel mai important element structural al glomerulului.¹ Aceasta reprezinta singura bariera anatomica intre sange si urina si este delimitata pe de o parte de endoteliul fenestrat al capilarelor si de cealalta parte de invelisul podocitar toate cele trei structuri (straturi ) alcatuind filtrul glomerular. MBG prezinta la randul ei 3 straturi cu densitate optica diferita : un strat central dens omogen - lamina densa si doua straturi cu densitate optica joasa, mai putin dense - lamina rara interna respectiv lamina rara externa [ ³]. Ca si epiteliul podocitar, MBG are o incarcatura electrica negativa conferita de proteoglicani (heparansulfatul), care in conditii normale impiedica filtrarea albuminelor. Structura complexa a filtrului glomerular este permeabila pentru apa si molecule mici, dar retine cea mai mare parte a proteinelor, alte molecule mai mari si elemente figurate ale sangelui.
Aparatul juxtaglomerular (AJG) - reprezinta o structura specializata situata la polul vascular al corpusculului Malpighi in contact intim cu arteriola aferenta, arteriola eferenta, tubul distal si mezangiul extraglomerular [²]. Este alcatuit din : glomus, macula
densa si lacis. De asemenea aparatul juxtaglomerular se poate subimparti in doua componente : vasculara alcatuita din portiunea terminala a arteriolei aferente, portiunea initiala a arteriolei eferente si regiunea mezangiala extraglomerulara si una tubulara alcatuita din macula densa.[5]
Glomusul este alcatuit din celule mioepitelioide cu caracter de celule glandulare si contractile, localizate in peretele arteriolei aferente inaintea diviziunii ei si primeste numeroase terminatii nervoase simpatice.
Macula densa este situata la nivelul tubului contort distal in zona de contact a acestuia cu polul vascular, cu o dispozitie celulara caracteristica 'in palisada' avand rolul de a transmite celulelor mioepitelioide informatii despre volumul, presiunea si compozitia ionica a lichidului tubular.¹
Lacisul este reprezentat de mezangiu extraglomerular care se continua cu mezangiu intraglomerular. Celulele mezangiale sunt dispuse in mai multe straturi si au proprietati contractile. Contractia celulelor mezangiale ( pe care se sprijina direct capilarele glomerulare) modifica dispozitia spatiala a capilarelor si presiunea hidrostatica a capilarelor. Ele moduleaza si transmit semnale emise de celulele maculei densa catre celulele glomusului.( Figura 1.2)
Aparatul juxtaglomerular intervine in reglarea circulatiei sangvine la nivel glomerular si a echilibrului hidroelectrolitic prin baroreceptorii de la nivelul glomusului ce detecteaza scaderea presiunii arteriolare si prin chemoreceptorii de la nivelul maculei densa ce detecteaza variatiile volumului si/sau compozitiei urinii. Tubul renal incepe la polul urinar al glomerulului si este alcatuit dintr-un segment proximal, unul intermediar si altul distal.¹
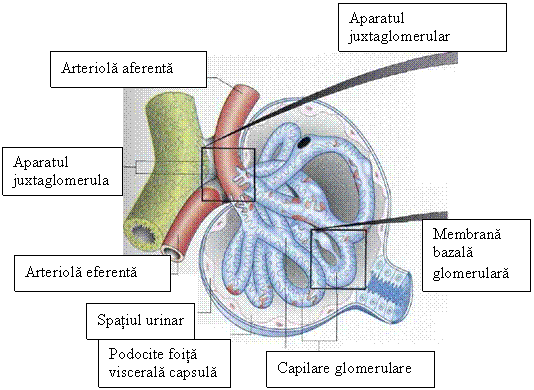
Figura 1.2. Corpuscul renal - adaptare dupa Gray s Anatomy
Tubul proximal are o lungime de 14-15mm si prezinta o portiune initiala tortuoasa - pars contorta sau pars convoluta si o portiune descendenta, rectilinie - pars recta.³ Tubul contort proximal format dintr-un singur strat de celule epiteliale cilindrice asezate pe o membrana bazala are drept functie esentiala reabsorbtia apei si a unor electroliti, in timp ce in tubul drept proximal format din epiteliu asemanator cu cel al partii contorte are loc secretia de acizi organici si baze. In continuarea portiunii drepte a tubului proximal se descrie ansa Henle cu un ram descendent subtire care coboara in medulara, se incurbeaza si revine in cortex printr-un ram ascendent larg.²Ramura descendenta subtire este acoperita de un epiteliu turtit, cu celule neregulate in timp ce ramura ascendenta groasa este formata din epiteliu cu celule cubice. Pentru segmentul subtire al ansei Henle au fost descrise 4 tipuri de celule (epiteliul de tip I, II, III, IV) cu permeabilitate diferita pentru apa. In functie de lungimea ansei Henle nefronii sunt impartiti in: nefroni lungi (20% dintre nefroni) - cu ansa Henle lunga, implicati in concentrarea urinii si nefroni scurti (80% dintre nefroni) cu ansa Henle scurta ce participa la reglarea sodiului.³
Tubul distal reprezinta segmentul nefronal dintre ansa Henle si tubul colector si este format la randul lui dintr-o portiune dreapta ascendenta - pars recta si o portiune distala
contorta - pars convoluta care prezinta la inceput zona specializata denumita macula densa descrisa mai sus. Lumenul tubului distal este marginit de un epiteliu cubic simplu care la nivelul maculei densa se modifica avand o dispozitie caracteristica 'in palisada'.
Segmentul conector - reprezinta o regiune de tranzitie intre nefronul distal si ductul colector. La acest nivel exista trei tipuri de celule: celule epiteliale ale tubului contort distal, celule proprii tubului conector si celule principale, proprii tubului colector.[¹]
Tubul colector este alcatuit dintr-un sistem de tubuli care incep in corticala, se unesc in tubi colectori si formeaza ductele papilare Bellini ce se deschid la nivelul ariei cribrosa pe suprafata papilei renale.[¹] Este alcatuit din doua tipuri de celule: principale, clare si intercalare, intunecoase.
Vascularizatia rinichiului
In conditii bazale rinichiul primeste 20% din debitul cardiac. Debitul sangvin renal este de 1200 ml/min la un debit cardiac de 5l/min. Fiecare rinichi este vascularizat de o artera renala. Arterele renale se desprind aproape in unghi drept din aorta abdominala imediat dedesubtul arterei mezenterice superioare. Cu putin inaintea hilului, fiecare artera renala se imparte intr-o ramura anterioara si una posterioara, care mai departe vor da 5 segmente (artere segmentare) : apicala, superioara, medie, inferioara si posterioara. Ramurile initiale ale arterelor segmentare dau ramuri lobare cate una pentru fiecare piramida renala. La nivelul bazei piramidelor Malpighi, artera interlobara se curbeaza si da nastere arterelor arcuate. Din acestea emerg arterele interlobulare (artere radiale corticale) dispuse intre piramidele Ferrein. Din artera radiala corticala iau nastere arteriolele aferente ale corpusculilor Malpighi. Arteriolele eferente se capilarizeaza peritubular si dreneaza ulterior intr-o vena corticala.3 Arteriolele eferente al glomerulilor juxtamedulari se ramifica si dau nastere la vasa recta descendente si ascendente. Vasa recta sunt permeabile pentru apa si sodiu si participa la mentinerea gradientului cortico-medular.
Circulatia venoasa este superpozabila celei arteriale pornind de la nivelul capilarelor peritubulare.[¹] Venele corticalei se formeaza la nivelul plexului peritubular dreneaza in venele interlobulare ce insotesc arterele cu acelasi nume. Venele medularei se formeaza la nivelul plexului capilar al medularei interne, converg in vasele drepte ascendente, care se aduna in fascicule de-a lungul vaselor descendente corespunzatoare. In regiunea subcorticala, vasele drepte ascendente dreneaza in venele arcuate ce insotesc arterele cu acelasi nume si dreneaza in venele interlobare. Acestea ajung in sinus, formeaza un trunchi cu trei ramuri anterioara, posterioara si intermediara care se unesc pentru a forma vena renala.[2]
Limfaticele rinichiului se constituie in doua retele limfatice diferite una superficiala capsulara si un sistem hilar mai profund.[5] Originea sistemului limfatic cortical consta in capilare limfatice interlobulare ce se afla in jurul arterelor cu acelasi nume si dreneaza in vasele limfatice arcuate situate la nivelul jonctiunii cortico-medulare. Acestea dreneaza in vasele limfatice interlobare care merg catre vasele limfatice hilare.[²] Sistemul limfatic subcapsular este mai putin dezvoltat si dreneaza in limfaticele subcapsulare.[¹]
Inervatia rinichiului este de natura vegetativa si provine din : ganglionii celiac, mezenteric superior, hipogastric superficial, splahnic superior, din lantul simpatic si cateva fibre vagale.[¹] Fibrele nervoase sunt situate in tesut conjunctiv lax, insotesc arteriolele renale si ramificatiile lor, se distribuie la nivelul peretilor arteriolari, la nivelul aparatului juxtaglomerular si la polul bazal al unor tubi.
Fiziologia rinichiului
Rinichiul indeplineste o serie de functii importante in organism:
1. Functia excretorie - de eliminare prin urina a produsilor de catabolism cum sunt ureea, acidul uric, creatinina, produsii finali ai catabolismului hemoglobinei, metabolitii diferitilor hormoni precum si eliminarea substantelor chimice sau a metabolitilor de tipul medicamentelor, pesticidelor aditivilor alimentari etc.
2. Functia homeostatica - de mentinere constanta a volumelor, osmolaritatii si compozitiei organismului, a echilibrului hidro-electrolitic, acido-bazic si al diferitelor substante organice.
3. Functia metabolica - de catabolizare a unor hormoni polipeptidici (insulina, glucagon, PTH, gastrina, vasopresina etc) si cea de gluconeogeneza.
4. Functia endocrino-metabolica - concretizata prin :
Secretia de renina de catre celulele mioepiteliale din arteriola aferenta, enzima proteolitica care actioneaza asupra angiotensinogenului (sintetizat de ficat) dand nastere angiotensinei I (forma inactiva) care este transformata in angiotensina II (ATII forma activa) gratie enzimei de conversie existente in plasma. La randul sau ATII stimuleaza secretia de aldosteron formand sistemul renina-angiotensina-aldosteron cu rol important in homeostazia tensionala.
Secretia de eritropoietina - hormon peptidic elaborat in interstitiu de celulele endoteliale care stimuleaza eritropoieza in maduva hematogena mentinand astfel homeostazia hematopoietica.
Hidroxilarea vitaminei D luand nastere 1,25- dihidroxivitaminaD3, forma activa a vitaminei D cu rol important in homeostazia fosfo-calcica.[6]
Functia glomerulara
Principala functie a glomerulului este formarea filtratului glomerular, prima etapa a excretiei renale. ²Etapa initiala a procesului de formare a urinii, filtrarea plasmei la nivelul capilarului glomerular are ca rezultat un fluid total lipsit de proteine - ultrafiltratul (urina primitiva) in cantitate de 120-250 l/zi.[6]Traversarea fluidului plasmatic din capilarele glomerulare catre spatiul urinar se realizeaza la nivelul 'membranei de filtrare' alcatuita din endoteliul capilar, membrana bazala glomerulara si stratul celulelor epiteliale.[3, 6] Aceasta bariera structurala si electrochimica nu lasa sa treaca proteinele sau substantele cu greutate moleculara egala sau mai mare de 70000 daltoni.[³]
Factorii ce regleaza filtrarea glomerulara sunt:
Presiunea eficace de filtrare (PEF) reprezinta forta neta ce transporta apa si solvitii prin membrana glomerulara. Expresia matematica a PEF este :
PEF = (PH - PHB) - PO unde PH - presiunea hidrostatica din capilarul glomerular (45mmHg)
PHB - presiunea hidrostatica din capsula Bowman (10mmHg)
PO - presiunea coloid - osmotica din capilare ( 25 mmHg). Valoarea normala a PEF este de 10 mmHg.[6]
Coeficientul de filtrare (Kf) al membranei glomerulare depinde de suprafata membranei capilare si de permeabilitatea acesteia. Cu ajutorul sau, rata filtrarii glomerulare (RFG) se exprima astfel : RFG = Kf x PEF . Valoarea sa normala este de 12,5 ml/min/mmHg. Valoarea normala a ratei filtrarii glomerulare este de 125 ml/min/1,73 m² suprafata corporala.[¹]
Determinantii filtrarii glomerulare sunt : presiunea hidrostatica glomerulara - factor major
fluxul plasmatic renal - cu rol secundar influentat de mecanismele de autoreglare ale presiunii arteriale, autoreglarea simpatica si rezistenta arteriolara.
Autoreglarea presiunii arteriale se realizeaza intr-o marja larga de valori presionale (90-190 mmHg) .Autoreglarea devine nula sub valoarea de 80 mmHg a presiunii arteriale, persista dupa denervarea rinichiului si dupa transplant renal, si este mediata de schimbari in rezistenta arteriolelor eferente.
Stimularea simpatica. In cadrul hipotensiunii sistemice marcate sistemul vegetativ simpatic stimulat produce vasoconstrictia arteriolelor aferente si eferente cu cresterea rezistentei vasculare renale si scaderea RFG. Hipertensiunea sistemica are ca raspuns vegetativ scaderea tonusului simpatic cu diminuarea rezistentei vasculare renale si cresterea RFG.[¹]
Reglarea procesului de ultrafiltrare glomerulara se realizeaza prin interventia substantelor vasoactive dilatatoare si constrictoare. Dintre substantele vasodilatatoare amintim : acetilcolina, bradikinina, prostaglandinele E1, E2, I2, histamina, factorul relaxant derivat din endoteliu (EDRF) si adenozin monofosfatul ciclic (AMPc).
Substantele vasoconstrictoare mai importante sunt : norepinefrina, angiotensina II, endotelinele, leucotrienele, factorul activator plachetar, factorul de crestere epidermala, lipoxinele A si B.
Functia tubulara
Dupa elaborarea urinii primitive, aceasta sufera la nivelul tubilor procese de reabsorbtie si sau de secretie rezultand urina finala.
Tubul contort proximal are ca functie esentiala reabsorbtia apei si a unor solviti. Are loc o reducere importanta a ultrafiltratului (in proportie de aproximativ 70%) . Aceasta absorbtie este izotonica legata de permeabilitatea importanta si omogena a acestui tub pentru apa, NaCl, uree etc. Glucoza este reabsorbita activ si in intregime in intregul tub proximal, cu conditia ca nivelul sangvin sa nu depaseasca 10 mmol l. Bicarbonatii sunt in intregime reabsorbiti daca nivelul plasmatic este < 27 mmol/l. In pars convoluta a tubului proximal sunt reabsorbiti Na+, K+, Clˉ, HCO3ˉ, Ca2+, HPO42+, aminoacizi, glucoza si apa. In pars recta a tubului proximal are loc in principal secretia de acizi organici si baze. Aceasta portiune este susceptibila la injuria nefrotoxicelor (metale grele si droguri).2 Ansa Henle este implicata in mecanismele de concentrare si dilutie a urinii.
Functia de concentratie a urinii este consecinta activitatii enzimatice importante de la nivelul portiunii largi a ansei Henle si portiunii initiale a tubului contort distal.[2] Rinichiul are capacitatea de a ajusta compozitia urinii eliminate in functie de necesitatile zilnice ale organismului, mentinand constanta osmolaritatea fluidelor sale. Principalul reglator al compozitiei urinare este ADH-ul. Mecanismul esential de concentrare al urinii il reprezinta multiplicarea in contracurent.[¹] In acest mecanism sunt implicate :
Ramura descendenta si ascendenta a ansei Henle sunt asimilate cu un tub in forma de U (cu fluidul care circula in directii opuse) ramurile fiind separate prin tesut interstitial medular. Fluidul izoosmotic intra in ramura descendenta subtire si este concentrat pe masura ce strabate ansa Henle. Dincolo de ansa Henle este diluat, devenind hipoton fata de plasma. Tubul in U are urmatoarele proprietati care ii permit mentinerea contracurentului multiplicator :
- ramura subtire descendenta este permeabila pentru apa si impermeabila pentru sodiu si uree; urinile tubulare devin tot mai hipertonice dinspre cortex spre medulara profunda;
- ramura ascendenta subtire este impermeabila pentru apa si uree, dar reabsoarbe NaCl dupa legile osmozei, ceea ce face ca urina sa piarda din hipertonicitate;
- ramura ascendenta larga si tubul contort distal in portiunea initiala sunt impermeabile pentru apa si uree si permeabile pentru sodiu. La acest nivel Na+, K+, Clˉ, sunt reabsorbita activ, urinile devenind hipotonice;
- segmentul terminal al tubului distal si canalul colector cortical reabsorb apa si urina devine izotonica. In prezenta ADH, devin tot mai permeabile pentru apa, dar nu si pentru uree;
- canalul colector medular in prezenta ADH este permeabil in particular pentru uree, apa dar si pentru Na. Urina finala devine hipertonica;
- vasa recta permit schimburi osmotice cu tesutul interstitial medular si prin intermediul acestuia cu diferitele segmente ale ansei Henle. Ele cedeaza apa tesutului interstitial hipertonic si primesc solviti din interstitiu. Urmarea acestor miscari ale apei si solvitilor este dezvoltarea unui dublu gradient :
Insumarea schimburilor osmotice transversale are drept rezultat dezvoltarea unui gradient longitudinal cortico-medular, ceea ce justifica termenul de multiplicare contracurent.[²]
Dilutia urinii este realizata de aceleasi structuri ca si concentrarea acesteia. In starile cu exces de apa, osmoreceptorii sesizeaza o reducere a osmolaritatii plasmatice, cu oprirea secretiei de ADH. In absenta ADH, canalele colectoare devin impermeabile la apa. Ramura ascendenta groasa a ansei Henle reabsoarbe suficient NaCl pentru a dilua fluidul luminal, chiar in absenta ADH. Lichidul ajuns in tubul contort distal are o osmolaritate de 50 mOsm. In absenta ADH, acest fluid traverseaza canalul colector si este excretat ca urina. Alt mecanism de dilutie urinara este intreruperea reciclarii ureei, prin lipsa ADH. In absenta ADH, reabsorbtia ureei este mult redusa. Lipsa reciclarii ureei scade osmolaritatea medularei interne ceea ce va duce la excretia unei urini diluate.[¹]
Intre reabsorbtia tubulara si filtrarea glomerulara exista interrelatii.
Balanta glomerulotubulara adapteaza reabsorbtia tubulara proximala de Na+ si apa la rata filtrarii glomerulare, datorita variatiilor presiunilor hidrostatice si oncotice in capilarele glomerulare si la nivelul capilarelor peritubulare. Pentru mentinerea homeostaziei Na+ si a apei, ori de cate ori apar modificari in rata de filtrare glomerulara, la nivelul tubului proximal, sodiul reabsorbit reprezinta o fractie constanta (si nu un volum constant) din cantitatea de Na+ filtrat.
Recontrolul tubuloglomerular este un mecanism prin care rata filtrarii glomerulare si fluxul plasmatic renal se autoregleaza. Originea sa se gaseste la nivelul tubului distal si al aparatului juxtaglomerular. Celulele maculei densa ale tubului distal sunt in conexiune cu peretele arteriolei aferente ce secreta renina si cu celulele lacisului, care se prelungesc prin mezangiu glomerular. Variatiile debitului urinar sau concentratiile in NaCl in tubul distal sunt susceptibile sa influenteze vasomotricitatea arteriolei aferente, si astfel, sa regleze filtrarea glomerulara a nefronului omolog.[²]
Functia endocrino-metabolica
Secretia de renina se produce in principal la nivel renal existand si structuri extrarenale secretante de renina.¹ Renina este secretata la nivelul aparatului juxtaglomerular sub forma de preprorenina. Stimulii majori ai eliberarii de renina sunt reprezentati de reducerea TA medii, hipovolemia, reducerea nivelului de catecolamine, concentratia crescuta de NaCl in macula densa. Angiotensina II isi exercita actiunile prin intermediul receptorilor AT 1 si AT 2. Ang II este un puternic vasoconstrictor, stimuleaza secretia de aldosteron si catecolamine, retentia de Na+ in tubul proximal, stimuleaza centrul setei. La nivel renal, Ang II provoaca vasoconstrictia arteriolei aferente si a arteriolei eferente, reducerea fluxului plasmatic renal, cu mentinerea ratei de filtrare glomerulara.
Aldosteronul este un hormon steroid sintetizat la nivel corticosuprarenal. Secretia este stimulata de Ang II, K+, ACTH si inhibata de hormonul natriuretic atrial. La nivel renal aldosteronul actioneaza prin stimularea Na+- K+- ATP-azei de la nivelul membranei bazolaterale, provocand reabsorbtia de Na+ , secretia de K+ si de H+ la nivelul tubului colector.
Prostaglandinele renale sunt acizi grasi nesaturati, derivati din acidul arahidonic sub actiunea ciclooxigenazei. Prostaglandinele au atat proprietati vasodilatatoare cat si proprietati vasoconstrictoare. In conditii patofiziologice (hemoragie, insuficienta cardiaca congestiva, depletie de volum) prostaglandinele vasodilatatorii (PG E2, PG I2) contracareaza efectele angiotensinei II si ale catecolaminelor. Tromboxanul A2 are efect vasoconstrictor renal cu reducerea fluxului sangvin renal si a ratei de filtrare glomerulare.[²]
Eritropoietina este un hormon glicoproteic sintetizat la nivel renal, care controleaza productia de eritrocite prin stimularea proliferarii si diferentierii celulei stem eritroide.[¹]
Productia extrarenala de eritropoietina este predominant hepatica. Sinteza este stimulata de hipoxie, determinata de anemia sau ischemia rinichiului. Productia de eritropietina este stimulata de mai multi factori : HIF-1 (hypoxia inducible factor 1), prostaglandine, vasopresina, radicali liberi de oxigen, androgeni. Antiinflamatoarele nesteroidiene blocheaza productia de eritropoietina; de asemenea unele citokine (interleukina 1, factorul de necroza tumorala α) inhiba productia de eritropoietina. Pe langa rolul esential in eritropoieza, eritropietina prezinta si efecte pleiotrope, in principal de limitare a apoptozei in leziunile de ischemie - reperfuzie la nivel renal, miocardic si cerebral.
Vitamina D este produsa in piele ca proviatmina D. Vitamina D este hidroxilata, in primul rand la nivelul ficatului rezultand 25-hidroxi-vitamina D si din nou, la nivel renal rezultand 1,25-dihidroxi-vitamina D. Acest metabolit este cel mai activ asupra absorbtiei intestinale de calciu.
Pe langa numerosii receptori pe care ii contine pentru hormonul paratiroidian si aldosteron, rinichiul participa de asemenea, la degradarea a numerosi hormoni polipeptidici (insulina, glucagon, gastrina, parathormon). Nivelul plasmatic al unora dintre acesti hormoni poate creste in cursul insuficientei renale, ca urmare a cresterii duratei lor de viata. Acest mecanism poate explica cresterea nivelului gastrinei in boala cronica de rinichi sau diminuarea necesitatilor de insulina la diabetici, atunci cand survine insuficienta renala.[2]
Bibliografie
Alexandru Ciocalteu . Tratat de Nefrologie. Editura National, 2006, 17-31
Nicolae Ursea. Tratat de Nefrologie. Editia a II a, Editura Fundatia Romana a Rinichiului. 2006: 65-240,
Adrian Covic, Maria Covic, Liviu Segall, Paul Gusbeth -Tatomir. Manual de Nefrologie. Editura Polirom, 2007: 13-31
Gray' Anatomy. The Anatomical Basis of Clinical Practice. Thirthy- ninth edition, Elsevier Limited 2005: 1273-1279
Brenner & Rector`s. The kidney- 8th edition. Saunders Elsevier, 2008: 37-38
Nicolae Ursea. Manual de Nefrologie. Editura Fundatia Romana a Rinichiului , 2001: 12-74
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 12241
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved