| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
ELEMENTELE FIGURATE ALE SINGELUI
ERITROCITELE
1.Notiuni generale de fiziologie a singelui. Singele este alcatuit
din plasma si elemente figurate: hematii (eritrocite), leucocite
si trombocite. Impreuna cu limfa, lichidul interstitial si lichidele transcelulare, singele alcatuieste
compartimentul lichidian extracelular (LEC), functional sinonim cu mediul intern. Mentinerea
2.Compozitia singelui. Hematocritul.Singele, ca sistem fizic, e compus din faza dispersata (elementele figurate) si faza dispersanta, plasma, care se separa prin centrifugare.Volumul sanguin (volemia) este alcatuit din volumul plasmatic si cel globular. Volumul globular exprimat procentual este hematocritul (Ht). Determinarea volumului sanguin se face prin metoda dilutiei. Primele determinari au fost facute la condamnati la moarte prin decapitare (Birschoff-1857). Volemia la un adult este de 5 litri, adica 65-70mL/kgc, ceea ce reprezinta 8% din greutatea corporala, din care peste 4% sint determinate de volumul plasmatic si 3% de volumul globular. Diferenta dintre sexe, la adulti este de aproximativ 1 litru in favoarea sexului masculin. Diferentele se datoreaza hormonilor sexuali, in ceea ce priveste Ht si concentratia de hemoglobina (Hb) si procentului de tesut adipos, mai slab vascularizat. Ca dovada, dupa castrare, diferentele de volemie dintre cele doua sexe practic dispar. Volumul sanguin se raporteaza si la suprafata corporala: 3,1 L/m2 la barbati si 2,5 L/m2 la femei, exprimare corecta in caz de retentii hidrosaline importante (edeme de diferite cauze) si in cazul obezilor. La copiii nou nascuti la termen, raportul volemie/greutatea corporala este de 80-100 mL/kgc, datorita volumul eritrocitar mai mare. Acest fapt se datoreaza eritropoiezei, mai puternic stimulata in timpul vietii intrauterine, prin intermediul eritropoietinei, eliberata in cantitate mai mare in conditii de hipoxie.
La prematuri, raportul este
si mai mare, de 108 mL/kgc, dar in acest caz el se datoreaza
volumului plasmatic crescut. Determinarea volemiei se face cel mai corect prin
determinarea simultana a volumului plasmatic (VP) si a volumului globular
(VG). Determinarea VP se face prin administrarea pe cale endovenoasa de coloranti (albastru Evans, denumit
si solutie T1824, sau cu Rosu de
Valorile Ht astfel obtinute dupa corectare prin inmultire cu F1 si F2, sint de 42% pentru femei si de 45% pentru barbati. La nou nascut, Ht are o valoare mai mare, de 56%, datorita numarului crescut de hematii (poliglobulie) si datorita unei usoare deshidratari specifice acestuia. Ht venos de 45% este putin mai mare fata de cel arterial, de 42%, datorita trensferurilor hidroelectrolitice de la nivelul segmentului capilar: hematia din singele venos contine mai mult clor si mai multa apa datorita fenomenului de membrana Hamburger.
Valoarea Ht variaza si in functie de organul in care este determinat: astfel, la nivel splenic, Ht este de 70% in capilarele sinusoidale splenice, care "filtreaza" eritrocitele. La nivelul capilarelor si venulelor, Ht este mai mic, datorita curgerii axiale a singelui, iar la nivel tisular, Ht este mai mic fata de Ht arterial si venos. Ht intregului organism reprezinta media valorilor Ht din toate tesuturile si organele. Astfel, Ht somatic este de 35%. Raportul dintre Ht somatic si Ht venos se numeste factor celular si este subunitar: 0,91 la adult si 0,87 la nou nascut. Ht este direct proportional cu numarul de hematii dintr-un mm3 de singe, daca volumul eritrocitar mediu (VEM) este normal (80-94μ3).
Ht creste in poliglobulii (pletora globulara), insotite si de hipervolemie. In plasmoragii (pierderi de plasma), volemia scade, dar Ht creste datorita hemoconcentratiei. Ht scade in anemiile cu volemie normala sau scazuta; in hiperhidratari (hidremii), volumul globular este normal, dar Ht scade datorita hemodilutiei. In cazul hemoragiilor acute, inaintea declansarii mecanismelor compensatorii pentru corectarea hipovolemiei prin intravazarea lichidului interstitial, Ht este normal, dar volumul globular este scazut.
3.Modificarile fiziologice ale volemiei.
3.1.Factorii endogeni.
Digestia determina o usoara si lenta crestere a volemiei, datorita absorbtiei lichidelor in intestin.
In timpul efortului fizic intens, volemia scade cu citeva sute de mL in primele 10-15 minute, datorita extravazarii de lichid in spatiul interstitial, ca urmare a cresterii numarului de capilare functionale. La subiectii antrenati, acest proces e mai putin intens. Volemia este direct proportionala cu activitatea fizica: la sportivii bine antrenati, raportul volemie/greutatea corporala este de 100 mL/kgc.
Postura. Dupa 30 de minute de ortostatism, volumul sanguin este cu 15% mai mic fata de clinostatism, datorita extravazarii lichidului din plasma la nivelul capilarelor membrelor inferioare, ca urmare a cresterii presiunii hidrostatice la acest nivel.
Repaosul la pat pe termen scurt determina cresterea volemiei cu 5%. Prelungit, (2-3 saptamini) acesta determina scaderea volumului plasmatic la adult cu pina la 500 mL.
Sarcina determina cresteri mari ale volemiei, in medie cu 20-30%, uneori cu 100%. Cresterea volemiei incepe din saptamina 10 si continua progresiv pina in saptaminile 30-34, raminind nemodificata pina la nastere. Revenirea la normal se face lent, dupa 2-8 saptamini de la nastere. Aceasta crestere se datoreaza cresterii atit a VP, cit si a VG. VP creste datorita retentiei hidrosaline din timpul sarcinii, ca urmare a secretiei crescute de aldosteron si ADH si datorita cresterii sintezei de proteine plasmatice. Cresterea VG in sarcina variaza direct proportional cu aportul de Fe exogen. Cauza cresterii de VG este descarcarea de eritropoietina, stimulata de prolactina si de hormonul lactogen placentar (somatomamotropina corionica umana,denumit si prolactina placentara - hPL).
3.2.Factori exogeni. Climatul cald induce usoara crestere a volemiei, iar cel rece o usoara scadere, datorita transferului de lichid in tesuturi (ficat, muschi). La mare altitudine, presiunea atmosferica scazuta si hipoxia prelungita determina cresterea volemiei.
4.Variatiile patologice ale volemiei.
4.1.Scaderea volemiei reprezinta hipovolemiile sau oligohemiile.
Acestea apar in: pierderi externe de singe (traumatisme, interventii chirurgicale) sau interne (ruptura de organ: ficat, splina). Scaderea volumului globular caracterizeaza anemiile. Scaderea VP se datoreaza pierderii de plasma si de lichide hidroelectrolitice in arsuri, varsaturi, diarei profuze, fistule digestive, ocluzii intestinale inalte, diureze excesive, diabet zaharat (poliurie), insuficienta a glandei corticosuprarenale, transpiratii profuze, edeme generalizate, acumularea de mari cantitati de lichid in pleura sau peritoneu. Hipovolemiile sint insotite sau nu de modificarea Ht. In hemoragiile acute, la inceput, hipovolemia este normocitemica (Ht este normal), apoi, pe masura intravazarii de lichid interstitial, Ht incepe sa scada si hipovolemia devine oligocitemica (Ht scazut). In cazul pierderilor de plasma si/sau lichide electrolitice, datorita hemoconcentratiei, hipovolemia este policitemica (Ht crescut). Daca se pierd lichide hipotone (transpiratii, varsaturi, diureze apoase), plasma devine hipertona, hematiile pierd din volum si se ratatineaza, astfel scade VG, ca urmare scade si Ht, indicind pierderi mai mari ca in realitate. In cazul pierderilor de lichide mai putin hipotone, (bila, suc pancreatic) prin varsaturi sau fistule digestive, sau in cazul evacuarilor repetate ale revarsatelor din cavitatile pleurala sau peritoneale, plasma devine usor hipotona, hematiile cresc in volum, prin urmare creste VG si astfel Ht indica pierderi mai mici ca in realitate. In hemoragiile acute datorate leziunior venoase ce nu depasesc 10% din volumul sanguin total, presiunea arteriala (PA) practic nu se modifica; in cazul leziunilor arteriale ce duc la pierderi de sub 10% din volumul sanguin, PA se modifica, dar pe termen scurt si in mica masura, datorita declansarii prompte a mecanismelor compensatorii: mobilizarea singelui din depozite, intravazarea lichidului interstitial in capilare, oligurie (scaderea diurezei) pina la anurie (absenta diurezei), cu declansarea sistemului renina-angiotensina-aldosteron (R-A-A), scaderea secretiilor exocrine, ajustarea patului vascular la volumul sanguin scazut prin vasoconstrictie selectiva si redistributia singelui spre organe vitale; se sunteaza sectorul arteriolo-venular, deci scade patul vascular.
Daca hemoragia se soldeaza cu pierderea de peste 30% din volumul sanguin, se instaleaza socul. Acesta se instaleaza in hipovolemii absolute, datorate pierderilor lichidiene si in hipovolemii functionale: decompensarea acuta a ventriculului sting (VS) in cazul infarctului miocardic acut, al tamponadei pericardice (acumulare brusca a unei cantitati mari de lichid in sacul pericardic) sau in cazul rupturii de valve cardiace, in toate aceste cazuri survenind edemul pulmonar acut (prezenta de lichid in alveolele pulmonare).
4.2. Hipervolemiile sau pletorele sanghine se pot instala datorita cresterii VP in cazul hipervolemiilor oligocitemice, cind VG este normal sau usor scazut, asa cum se intimpla in retentiile hidrosaline la pacientii cu insuficienta renala, la cei cu ciroza hepatica si la cei cu insuficienta cardiaca.
Hipervolemia policitemica se caracterizeaza prin cresterea VG, fara modificarea VP. Poliglobuliile secundare sau adaptative sint intilnite la altitudine si in cazul pacientilor cu boli respiratorii cronice.
Poliglobuliile primare. Policitemia Vera este o tulburare primara a maturarii elementelor figurate la nivelul maduvei hematogene, in care valoarea Ht creste foarte mult.
Hipervolemia normocitemica se datoreaza cresterii atit a VP, cit si a VG, cu Ht normal: hipertiroidie, transfuzie de singe integral. Hipervolemiile acute se datoreaza perfuziei masive de diferite solutii: singe, plasma, masa eritrocitara, solutii cristaloide.
5.Repartitia volumului sanguin este inegala, cea mai mare parte, 85% gasindu-se in sistemul de joasa presiune, adica in rezervorul de capacitate, reprezentat de cord, venele mari, capilarele din marea circulatie, mica circulatie, iar restul de 15% sint repartizate in aorta si artere, ce reprezinta rezervorul de presiune. In conditii fiziologice si in repaos, jumatate din volumul sanguin total se afla efectiv in circulatie, ca volum sanguin circulant, in timp ce restul circula de10-12 ori mai lent sau stagneaza in sectoare ale sistemului venos, ca volum sanguin de rezerva. In ficat exista 0,5-1L de singe, in splina -0,3-0,5 L si, de asemenea, la nivelul plexurilor subpapilare. La aceste nivele, se pot acumula cantitati relativ mari de singe, fara modificarea presiunii intravasculare.
6.Controlul volemiei. Mecanismele de baza ale controlului volemiei sint aceleasi care controleaza presiunea arteriala (PA). In cazul cresterii volemiei, creste debitul cardiac, creste PA, creste presiunea in artera renala, creste filtratul glomerular, creste diureza si astfel se corecteaza volemia.
Natriureza si diureza variaza direct proportional cu valoarea PA. Pentru valori mici ale volemiei, mecanismul invers de retentie hidrosalina are la baza sistemul R-A-A. Intervin reflexul baro-receptor arterial si cel de intindere a receptorilor din zonele de joasa presiune ("stretch receptors"), denumit "reflex de volum"astfel: cresterea volemiei duce la cresterea PA, cresterea presiunii in artera pulmonara si alte zone intratoracice de joasa presiune, care excita baroreceptorii si receptorii de intindere; ca urmare, va fi inhibat sistemul nervos vegetativ simpatic, si astfel are loc vasodilatatia la nivelul arteriolelor renale si va creste diureza.
Hipervolemia stimuleaza si eliberarea de atriopeptina (hormonul natriuretic atrial), ca urmare vor creste natriureza si diureza de 3 pina la 10 ori, mecanismul nefiind, insa, de lunga durata. Concomitent, atriopeptina inhiba eliberarea de aldosteron. In cazul hipovolemiei, aldosteronul favorizeaza reabsorbtia de Na si apa in segmentele distale ale tubilor renali, ca urmare volemia creste cu 10-20% in primele doua zile dupa cresterea eliberarii de aldosteron. Pe masura ce creste PA, datorita cresterii volemiei, se instaleaza fenomenul "aldosteron escape", care consta in cresserea diurezei si natriurezei in ciuda aldosteronului eliberat in cantitate mai mare.
Volemia revine in citeva zile la
valori cu 5-10% mai mari ca normal, dupa fenomenul "aldosteron escape". In
mentinerea volemiei, intervin eliberarea de ADH si mecanismul setei, ca si mecanismul "apetitului
pentru sare". Acesta din urma intervine in hipovolemia din boala
Mentinerea normala a volemiei se datoreaza si schimburilor lichidiene de la nivelul capatului arterial al capilarului, interstitiului si capatului venos al capilarului, guvernate de un joc presional denumit "fortele Starling". Presiunea hidrostatica la capatul arterial este de 32 mmHg, cea coloid-osmotica de 28 mmHg, presiunea coloid-osmotica din interstitiu este de 5 mmHg, iar cea hidrostatica a lichidului interstitial este negativa, datorita suctiunii limfatice. Structurile solide din interstitiu dezvolta o presiune de sens contrar celei din lichid, iar turgorul cutanat (elasticitatea pielii) exercita o presiune de 2 mmHg. Suma algebrica a fortelor de la capatul arterial al capilarului este egala cu o presiune efectivaade filtrare de citiva mmHg, al carei efect este extravazarea de lichid din capilar in interstitiu: 0,3% din plasma capilara. La capatul venos, predomina forta de intravazare, datorita scaderii presiunii hidrostatice la 15 mmHg; reintra in capilar 9/10 din totalul lichidului extravazat, iar 1/10 ia calea limfei.
7.Proprietatile singelui.
7.1.Culoarea singelui se datoreaza Fe din hemoglobina (Hb), care reflecta razele spectrale rosii. Oxihemoglobina are culoarea rosu-aprins, Hb deoxigenata (redusa) are culoare rosu-inchis. Cianoza (culoarea albastruie a tegumentelor si mucoaselor) se datoreaza cresterii cantitatii de Hb redusa la 6-7% sau mai mult. In zonele cu staza sanguina sau circulatie incetinita, culoarea singelui este inchisa. Carboxihemoglobina (in care monoxidul de carbon inlocuieste oxigenul) are culoarea rosu-aprins, iar methemoglobina (al carei hem contine Fe+++) are culoare rosu bruna, cresterea concentratiei sale ducind la aparitia "cianozei brune".
7.2.Temperatura singelui creste paralel cu activitatea metabolica tisulara. La nivel hepatic, temperatura este de 41C. Cea mai scazuta temperatura este cea cutanata, datorita proceselor de termoliza de laacest nivel. Un factor esential pentru termoreglare este caldura specifica mare a singelui.
7.2.Densitatea singelui este de 1050-1070. Cea a plasmei este 1030, iar a hematiilor este 1090. Densitatea singelui creste in policitemii, iar a plasmei creste in pierderi hidro-electrolitice (pe cale digestiva), in arsuri, in hiperproteinemii patologice. Densitatea singelui scade in anemii, iar a plasmei scade in retentii hidrosaline(edeme) de cauza renala, hepatica, sau in casexie.
7.3.Presiunea osmotica (
7.3.Viscozitatea (V) singelui este un element al rezistentei intravasculare la curgerea singelui, pentru ca singele are un regim de curgere neturbulent. Ea se poate calcula din ecuatia Hagen- Poiseuille, in care fluxul sanguin Q = ΔP x r4 /8ŋL. Viscozitatea depinde de factori plasmatici (globuline alfa 1, alfa 2, beta si gamma si fibrinogen) si de factori eritrocitari. In afibrinogenemii, V scade foarte mult, iar in hiperfibrinogenemii, creste foarte mult. V variaza direct proportional cu Ht.
In hiperviscozitate, scade mult viteza de circulatie a singelui in microcirculatia hepatica, cerebrala hepato-splenica si exista tendinta la agregare intraarteriolara a hematiilor, ceea ce duce la ischemie si staza in acele teritorii, urmate de semne neurologice de focar ce sugereaza un accident vascular cerebral ischemic si semne de retinopatie. Hiperviscozitatea sanguina are consecinte foarte grave: infarcte fara tromboza, gangrena localizata fara ocluzie arteriala.
Hiperviscozitatea prin cresterea Ht este intilnita in Policitemia Vera.
Hiperviscozitatea este provocata si de cresterea agregarii eritrocitare in diferite situatii: arsuri, intoxicatii, embolii grasoase, transfuzii masive de singe.
Hiperviscozitatea prin scaderea temperaturii ambiante este intilnita in hipotermii spontane sau provocate.
Hiperiscozitatea prin scaderea deformabilitatii eritrocitare se intilneste in diferite hemoglobinopatii si alterari ale metabolismului eritrocitar.
Hipoviscozitatea se intilneste in anemii, hemodilutii, hipoproteinemii, hipertermii.
8.Functiile singelui.
8.1.Functia respiratorie se datoreaza transportului gazelor respiratorii.
8.2.Functia nutritiva este datorata transportului de substante energogenetice, plastice de la nivelul tubului digestiv la nivel tisular si de la organele de depozit la alte tesuturi.
8.3.Functia excretoare este datorata transportului catabolitilor la organele de eliminare (plamini, rinichi, piele, intestin, ficat, glande sudoripare).
8.4.Functia de control al echilibrului hidroelectrolitic este datorata schimburilor dintre plasma, lichidul interstitial si celule, care asigura izoionia (echilibrul ionic), izotonia (mentinerea presiunii osmotice a plasmei) si izohidria (mentinerea echilibrului acido-bazic). Pentru realizarea acestei functii, singele asigura transportul a numerosi hormoni implicati.
8.5.Functia de termoreglare este asigurata prin transportul de caldura de la organe cu metabolism intens (ficatul) la suprafata corpului, astfel realizindu-se termoliza. O importanta aparte in aceasta functie o detine caldura specifica mare a singelui, datorita apei, ce reprezinta 90% din compozitia singelui.
8.6.Functia de hemostaza, realizata prin trombocite si o serie de factori plasmatici indispensabili hemostazei si fibrinolizei,
8.7.Functia de aparare impotriva diversilor agenti patogeni si cea de eliminare a elementelor celulare imbatrinite, realizata prin cooperarea unor mecanisme complexe imune, specifice si nespecifice, ce implica elemente figurate ale singelui, elemente tisulare si proteine diverse, cu origine plasmatica sau tisulara.
8.8.Functia de coordonare si reglare, alaturi de SNC, asigurata de transportul hormonilor si a mediatorilor chimici, substante biologic active, toate implicate in mentinerea homeostaziei.
9.Hematopoieza este definita drept proliferarea celulelor progenitoare (precursoare), al caror numar este mentinut constant de celulele stem si diferentierea lor in toate componentele celulare ale singelui. Sediul hematopoiezei depinde de virsta individului si de starea fiziologica sau patologica a acestuia.
Eritropoieza este procesul prin care sint generate hematiile adulte.
In ontogeneza, etapele acesteia sint:
9.I.Etapa embriofetala, alcatuita din 3 subetape:
9.I.1.Etapa mezoblastica, in care eritropoieza este extraembrionara si intravasculara; astfel, in insulele sanguine mezodermale ale sacului vitelin, se observa in saptaminile 3-12 de sarcina hemangioblasti, celule mezenchimale sosite prin migrare din "linia primitiva" a blastomerului primitiv.Incepind cu a 22-zi, insule sanguine asemanatoare, dar mai putin numeroase se observa si in tesutul mezoblastic al embrionului. Insulele extraembrionare si intraembrionare vor fuziona, alcatuind un sistem vascular unic. In aceasta etapa, se sintetizeaza hemoglobina (Hb) Gower I, ce contine 2 lanturi ζ (zeta) si 2 lanturi ε (epsilon), care se va transforma rapid in Hb Portland, cu formula lanturilor globinice ζ 2 γ (gamma) 2 si apoi Hb Gower II, cu 2 lanturi α (alfa) si doua lanturi ε (epsilon). La fat se sintetizeaza Hb F, (fetala) cu 2 lanturi α si 2 lanturi γ.
9.I.2.Etapa hepatosplenica se caracterizeaza prin migrarea celulelor pluripotente la acest nivel, unde exista conditii superioare pentru hematopoieza. Ea ocupa intervalul cuprins intre saptamina 6 si luna a 5-a desarcina, spre sfirsitul caruia hematopoieza scade la acest nivel,dar se mentine in grad redus pina la 2 saptamini dupa nastere. La nivel splenic, hematopoieza se prelungeste pina in luna 8 de sarcina.
9.I.3.Etapa medulara incepe sa fie activa din luna a 5 a vietii, iar incepind cu luna 7, devine principalul sediu al hematopoiezei.
9.II.A doua mare etapa a hematopoiezei incepe dupa nastere. La nou-nascut si la copil, hematopoieza se desfasoara la nivelul tuturor cavitatilor epifizare si diafizare, si anume la nivelul diafizelor oaselor lungi si cavitatilor spongioase din oasele late si scurte. La adult, sediul hematopoiezei se limiteaza la nivelul oaselor scheletului axial -vertebre,coaste, stern, bazin, oase craniene,epifizele proximale ale femurului si humerusului. Maduva rosie este activa, functionala, iar cea galbena apare prin inlocuirea celulelor specializate cu adipocite. La 20 de ani, inlocuirea este completa, realizindu-se centripet, dintre extremitati spre trunchi. Adipocitele medulare difera mult de cele din tesutul adipos propriuzis: ele sint celule adventiceale, reticulare si macrofage transformate prin incarcarea cu grasime si detin si alte roluri, in afara celui de sustinere.
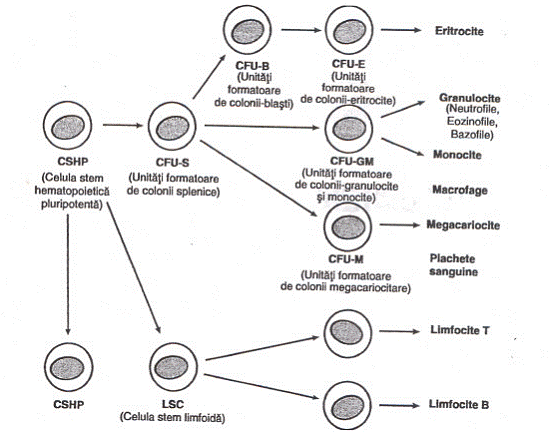
10.Populatia celulara a maduvei hematogene este alcatuita din: celule stem pluripotente, care sint cele mai timpurii. Sint desemnate prin prescurtarea CFU-S (colony forming units-spleen); aceste celule formeaza colonii pure si mixte de celule precursoare la nivel splenic, la soareci iradiati. Morfologic, aceste celule sint asemanatoare limfocitelor mici, dar sint incomplete din punct de vedere imunologic. La om, exista CFU-blast, celule pluripotente, asemanataoare celulelor CFU-S. Din aceste celule primordiale, se dezvolta 2 linii celulare:
Celule stem multipotente limfoide, din care deriva limfocitele T si B si
Celule stem multipotente mieloide, din care deriva precursorii pentru seria elementelor rosii (hematii), granulocite, monocite si megakariocite.
Maduva hematogena se caracterizeaza prin particularitati de microclimat;
stroma sa celulara contine celule endoteliale, fibroblasti si celule adventiceale, macrofage centrale, denumite "nurse cells", (celule doica) ce asigura Fe necesar sintezei Hb de catre eritroblasti si celule adipoase specifice.
Matricea extracelulara contine glicoproteine ce actioneaza ca situsuri de recunoastere si de adeziune pentru celulele stem hematopoietice, precum: fibronectina, laminina, colagenul, proteoglicanii.
Factorii de crestere hematopoietici sint hormoni ce apartin microclimatului medular si care detin si functii extramedulare. Ei controleaza multiplicarea si diferentierea celulelor stem in celule mature; intervin si in functionarea celulelor mature in anumite situatii (leucocite in infectii); Sint produsi de diverse celule medulare si extramedulare, precum limfocite, monocite, macrofage,celule endoteliale, fibroblasti.
11.Clasificarea factorilor de crestere hematopoietici.
11.1. Factorii nespecifici ce actioneaza pe celulele stem pluripotente si multipotente, denumiti multi- CSF(factors), cum este IL3, implicata in maturarea granulocitelor, monocitelor, eozinofilelor, mastocitelor,eritrocitelor si megakariocitelor.
11. 2. GM-CSF, factori implicati in maturarea granulocitelor si monocitelor si 11.3.Factori de crestere specifici, ce actioneaza pe o anumita linie celulara, mai exact asupra celulelor orientate, "comise" spre o anumita linie celulara. Astfel,exista G(granulocit)-CSF, M(onocit)-CSF, Eo(sinofil)-CSF, Eritropoietina, trombopoietina, precum si numeroase limfokine si monokine. Acesti factori detin numeroase functii hematopoietice, realizind o adevarata retea de efecte interactive, cum sint raspunsul imun la infectii si apararea antitumorala.
Interleukina1 (IL1) (denumita si factor endogen pirogen datorita efectului de hipertermie indus prin actiunea sa la nivel hipotalamic) este o glicoproteina produsa de macrofage in principal, dar si de endotelii, astrocite, fibroblasti si limfocite T, care detine o serie de functii, cum ar fi cea de stimulare a celulelor stromale ale maduvei hematogene, pentru sinteza si eliberarea de CSF.
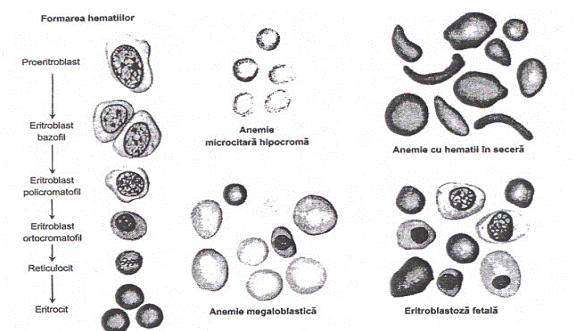
12.Eritrogeneza sau eritropoieza parcurge citeva etape: proeritroblast
-eritroblast bazofil - eritroblast policromatofil - eritroblast acidofil (ortocromatofil =
normoblast) - reticulocit (R) - hematia adulta. Pe parcursul acestor
etape, au loc importante modificari ale nucleului si citoplasmei: nucleul
dispare treptat, astfel incit este complet expulzat la eritroblastul acidofil,
apoi fagocitat. Pe parcursul maturizarii, scade numarul de ribozomi
si mitocondrii, precum si aparatul Golgi si sistemul de
microtubuli. Proeritroblastul are deja Hb, concentratia acsteia crescind
progresiv, pina ajunge la a reprezenta 1/3 din masa hematiei. Hematia
adulta contine de 10 ori mai multa Hb comparativ cu proeritroblastul. Sinteza de Hb in cadrul ciclului
celular este mai importanta in faza G2 si inceputul fazei
Eritropoieza este ineficienta cind reticulocitele sint blocate intre celulele endoteliale si ulterior fagocitate de macrofage. Comparativ cu reticulocitul, eritrocitul adult pierde organitele prin autofagie si expulzie, pierde capacitatea proprie de locomotie, contine mai multa Hb si pierde antigenele din sistemul HLA, ce exista pe membrana tuturor precursorilor, inclusiv la reticulocitele mai virstnice, detinind in schimb propriile sisteme antigenice, cu cele doua mai importante, AOB si Rh.
Multiplicarea precursorilor eritrocitari este de tip homoplastic, adica o celula produce doua celule fiice identice cu ea si intre ele, ceea ce asigura stocul celulelor stem, deci aceasta multiplicare are loc in cazul celulelor nediferentiate sau slab diferentiate.
Al doilea tip de multiplicare este de tip heteroplastic, sau maturizanta, in care celulele fiice difera de celula mama. Este prezenta pina la etapa de eritroblast policromatofil, care nu se mai poate divide. Marcarea ADN cu Fe59 si timidina tritiata a aratat ca proliferarea celulelor eritroide este asincrona, viteza fazelor ciclului celular difera de la un stadiu la altul. Relativ stabile par fazele S si M, cea mai variabila fiind G1. Numarul mitozelor in seria rosie este de 4: un proeritroblast formeaza 16 eritrocite.
In maduva hematogena exista un tip particular de eritroblasti cu diferite virste,denumiti sideroblasti, care contin granule de Fe neheminic, denumite siderozomi. Numarul granulelor scade progresiv, pe masura ce se acumuleaza Hb. Eritrocitele nu contin siderozomi. Timpul de tranzit medular al eritroblastilor este de 4-7 zile. Eritrocitul adult este o celula anucleata, de forma discoidala, desi sint denumite globule rosii si reprezinta faza dispersata a singelui. In singele periferic, exista 25 X 1012; la barbati exista 5 milioane de hematii/mm3 de singe, iar la femei - 4,5 milioane/mm3 singe. Aceste diferente, datorate hormonilor sexuali, nu exista pina la pubertate si dispar dupa climacterium.
13.Structura functionala a hematiei. Hematia are forma de disc biconcav, cu diametrul mediu de 7,2-8,3μ, grosimea scade in partea sa centrala, (1μ), fata de periferie, (2,2μ), ceea ce confera hematiei maxim de suprafata la un minim de volum. Volumul hematiei este de 80-90μ3. Deformabilitatea hematiei este o proprietate esentiala pentru functia ei.Viscozitatea interna a hematiei (1-2 centipoise) depinde de cantitatea si de calitatea Hb. Membrana hematiei este importanta pentru aceasta functie, mai ales prin componentele sale lecitina si lizolecitina. Se adauga si rolul factorilor citoscheletali, si anume prezenta pe versantul intern al membranei a actinei, ankirinei, spectrinei fosforilate, care depinde de proteinkinazele AMPc-dependente.
Defosforilarea spectrinei depinde de actiunea fosfatazelor, inhibate de 2,3-DPG (difosfoglicerat);calmodulina si glicoforinele prezente in citoplasma hematiei,ca si acidul sialic prezent la nivel membranar, sint implicati in deformabilitatea hematiei, proprietate care este principalul consumator de ATP, acesta fiind sintetizat 90% prin glicoliza anaeroba si 10% doar prin oxidarea glucozei.
Forma eritrocitului depinde de virsta si de cantitatea de ATP existenta. In singele conservat, ATP scade cu 50%, eritrocitele devin crenelate, apoi sferocite. Daca eritrocitele sint incubate cu inhibitori de glicoliza anaeroba (NaFl), apar modificari similare. Eritrocitele mai au proprietatea de a se dispune in fisicuri, agregarea eritrocitara fiind numita si simpexa hematiilor, fenomen reversibil, ce consta in circulatia hematiilor sub forma de trenuri, rulouri. Aceasta proprietate depinde de numarul hematiilor si de macromoleculele plasmatice circulante, precum albumine,globuline, fibrinogen, dextrani. Lungimea rulourilor de hematii influenteaza viteza de sedimentare a hematiilor (VSH). Rezistenta globulara a hematiilor la agresiuni mecanice,chimice,osmotice este cercetata prin rezistenta la hemoliza, hematiile fiind expuse la medii progresiv mai hipotone.
VSH exploreaza stabilitatea in plasma a hematiilor.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4053
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved