| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
FACTORII IMPLICATI IN IMUNITATEA NATURALA
Una din primele conditii ale realizarii unei infectii este trecerea barierelor anatomice, boala fiind rezultatul multiplicarii agentului etiologic in organism, fara a fi distrus de factorii umorali, celulari ai acestuia.
Factorii imunitatii naturale la organismele superioare se pot clasifica dupa mai multe criterii:
a) localizarea lor in organism, dupa care pot fi impartiti in: interni si externi;
b) felul in care participa la determinismul starii de imunitate, unii fiind factori predominant pasivi, iar altii activi;
c) nivelul lor de organizare biologica sau biochimica: unii sunt compusi chimici in marea lor majoritate prezenti in umori (factorii umorali), iar altii diferite celule raspandite in aproape toate organele si tesuturile care actioneaza prin insasi activarea lor biologica (factorii celulari);
d) complexitatea structurii sau functiei constituiente a factorului de rezistenta, deosebindu-se unii factori cu un grad redus de complexitate (factorii mentionati anterior), altii fiind complecsi, reprezentati de caracteristici constitutionale, constante morfologice sau fiziologice organismului, cum sunt: structura genetica, presiunea sanguina, temperatura normala, etc.
e) natura factorului in raport cu normalul si patologicul (hipertermia, inflamatia, productia de interferoni). Acestia din urma actioneaza dupa ce 'non-selful' a reusit sa se instaleze in organism si pot avea doua tipuri de consecinte, posibile pentru organism: unul salutar, in cazul cand 'non-selful' este facut inofensiv, altul nociv, cand mentinerea non-selfului duce la o amplificare a procesului patologic agravand evolutia bolii.
Luand in considerare criteriile de clasificare amintite factorii si mecanismele imunitatii naturale pot fi sistematizate conform tabelului nr.1:
Tabel nr. 1
|
Barierele naturale |
externe |
pielea mucoasele capcanele anatomice (cornetii nazali) |
|
interne |
peretii vaselor tesuturile dense membranele celulare |
|
Mecanis-mele de potentare a rolului barierelor |
regenerarea si exfolierea epiteliilor |
||||||
|
miscarea cililor epiteliilor ciliate |
|||||||
|
actiunea secretiilor glandelor |
mecanica |
spalare includere in mucus urmata de eliminare |
|||||
|
chimica |
acidul undecilenic (piele) acidul clorhidric (sucul gastric) factorii umorali diversi prezenti in lacrimi, lapte etc. |
||||||
|
antagonismul microbian intre flora normala autohtona si cea patogena autohtona sau alohtona |
|||||||
|
Factorii umorali |
lizozimul complementul (C) proteine de faza acuta (ex. PRC=proteina C reactiva) polipeptidele bazice beta-lizina opsoninele anticorpii naturali diversi interferonii (IFN) |
||||||
|
Factori celulari |
microfagele |
leucocite polimorfonucleare in general si mai ales cele neutrofile |
|||||
|
macrofagele |
mobile |
histiocitul monocitul |
|||||
|
fixe |
celulele Kuppfer macrofagele osteoclastele celule nevroglice |
splenice peritoneale alveolare din ganglioni limfatici din endoteliile vasculare |
|||||
|
Mecanisme complexe |
fiziologice |
temperatra normala circulatia sanguina si limfatica presiunea sanguina |
|||||
|
fiziopatologice |
hipertermia inflamatia elaborarea de interferoni |
||||||
|
Factori constitutionali si genetici cu caracter de: |
specie rasa linie individ |
||||||
In continuare vom descrie modul de actiune a unor factori enumerati in tabelul anterior.
1. Barierele naturale
a. Pielea si mucoasele se opun patrunderii germenilor patogeni in organism printr-o protectie mecanica functionala si chimica.
Protectia mecanica si functionala este determinata de integritatea lor morfofunctionala (cu exceptia virusurilor, leptospirelor, brucelelor, ankilostomelor care pot patrunde chiar prin pielea intacta), prin capcane anatomice si mobilitatea ciliara, care indeparteaza continu secretiile mucoaselor, incarcate cu diferite particule straine si microorganisme, de pe suprafata acestora.
Protectia chimica la nivelul pielii si mucoaselor se realizeaza prin reactia acida a secretiilor acestora (acidul undecilenic la piele, acidul clorhidric la mucoasa gastrica, acidul lactic la mucoasa vaginala, etc) si prin prezenta in continutul lor a unor substante cu rol bactericid cum ar fi: lizozimul, mucoproteinele si altele.
Lizozimul este o enzima mucolitica care produce liza bacteriilor Gram (+) si a carei actiune la nivelul mucoaselor este potentata de IgA secretorie.
Mucoproteinele sunt mai active contra diferitilor virusi iar acidul clorhidric are un efect bactericid foarte important la anumite pasari de prada (vulturul plesuv, pitulicea) care consuma adesea cadavre in descompunere. El permite distrugerea tuturor bacteriilor in stomac, permitand si mentinerea sterilitatii din duoden si jejun la anumite specii.
b) Antagonismul microbian intre flora normala autohtona si cea patogena alohtona se manifesta prin efectul de bariera (secretia unor bacteriocine active fata de alte bacterii patogene, competitia nutritionala si de ocupare a spatiului vital prin viteza de multiplicare, etc.).
Flora bacteriana saprofita actioneaza asupra agentilor patogeni in doua moduri:
competitie pentru elementele nutritive esentiale;
secretia unor substante bactericide;
Astfel, la nivelul intestinului se gasesc bacterii saprofite care secreta substante cu rol bactericid. De exemplu:
Escherichia coli secreta substante numite colicine care se insera la nivelul membranei bacteriei patogene si modifica potentialul energetic , determinand moartea acesteia;
Bacteriile saprofite de la nivelul epiteliului vaginal metabolizeaza glicogenul (produs de celulele epiteliale vaginale) si il transforma in acid lactic. Acidul lactic scade pH-ul vaginal creandu-se un mediu nefavorabil dezvoltarii bacteriilor patogene.
c) Spatiul extracelular prin gelul de mucopolizaharide bogat in acidul hialuronic se opune prin vascozitatea sa la mobilitatea bacteriilor cu exceptia celor producatoare de hialuronidaza.
d) Tensiunea locala de oxigen actioneaza prin nivelul ei ridicat sau scazut. Nivelul ridicat in pulmon permite, de exemplu, dezvoltarea bacilului tuberculozei (a bacteriilor aerobe) in timp ce diminuarea oxigenului din tesuturi atrage dupa sine germinarea sporilor de germeni anaerobi (Clostridium) dupa interventii chirurgicale.
2. Factorii umorali
Factorii umorali sunt reprezentati de: complement (alexina), lizozimul, interferonii, polipeptidele bazice, opsonine nespecifice, beta - lizine, proteine C reactive si anticorpii naturali diversi.
Complementul reprezinta un sisterm seric complex de 26 de substante proteice din plasma (C1-C26) denumite 'componente' care se activeaza succesiv una pe cealalta intr-o ordine data ('activare in cascada').(Fig.1)
Acestea se gasesc intr-o concentratie relativ constanta in serul normal. Cantitati mai mari de complement se gasesc in serul de cobai, iepure si nurca. Componentele complementului sunt sintetizate atat de macrofage si monocite cat si de catre celulele hepatice (cu exceptia C1 care ar fi sintetizata la nivelul epiteliului gastrointestinal si urogenital).
Elementul central consta in activarea fractiunii C3. Enzimele activatoare ('convertazele') sunt inscrise cu linie continua. Inchiderea buclei autocatalitice de activare a lui C3 pe cale alternativa este marcata cu linie intrerupta. Pentru simplificarea schemei s-a omis marcarea inactivatorilor si a inhibitorilor sistemului C' si alte detalii ale activarii finale.
Proteinele sistemului complement reprezinta 3 - 4% din totalul proteinelor plasmei (cca. 300 mg/ 100 ml).
Activarea complementului este declansata prin doua mecanisme ('cai'):
v calea clasica amorsata de complexe moleculare rezultate din cuplarea Atc. cu Atg. ('complexe imune').
v calea alternativa care este daclansata in afara complexului Atc/Atg la care participa factorii:
agregate moleculare de Atc;
polizaharidele ;
bacterii Gram pozitive sau Gram negative;
paraziti (ex. Tripanozoma);
endotoxine bacteriene;
levuri (ex. Candida albicans).
Calea clasica de activare a complementului.
Moleculele de imunoglobulina dupa ce au fixat antigenul sufera modificari conformationale care descopera situsul de legare cu C' - situat in zona Fc. Odata descoperit situsul de legare pentru C', leaga fragmentul C1q din complexul proteic C1 al C'. Acesta are forma unui buchet de lalele.(Fig.)-de introdus Extremitatea globuloasa se leaga la situsul pentru complement de pe molecula de Ig, iar tija (in prezenta Ca2+ )activeaza fragmentele C1s si C1r . Dupa activare, C1s si C1r s sunt convertite in enzime proteolitice numite "convertaze" (Fig.4)
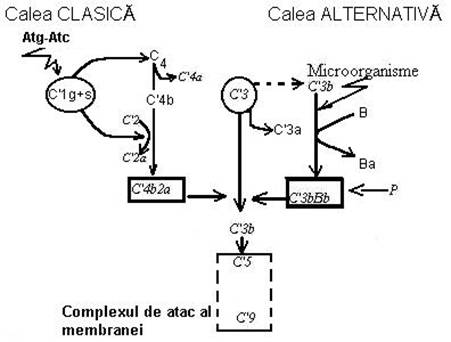
Fig.4. Modelul de activare enzimatica in 'cascada' a complementului
Ambele cai produc enzime specifice 'convertaze' care vor actiona asupra componentei a 3-a a sistemului (C3) rupand-o in doua fragmente C3a si C3b. C3a este un agent inductor de inflamatie, iar C3b se depune pe bacterii sau pe alte celule - tinta, pe care le elimina prin doua mecanisme: imunoaderenta si liza. O serie de fagocite (granulocite, neutrofile, macrofagele si celulele dendritice) exprima pe suprafata receptori specifici pentru C3b (denumiti CR1). C3b mijloceste pe aceasta cale atasarea celulelor - tinta pe fagocite ('imunoaderenta') si inglobarea lor ulterioara. Aceasta invelire cu C' a tintelor care faciliteaza fagocitoza a fost denumita 'opsonizare'. C3b poate media, totodata, continuarea activarii in cascada a sistemului care se va finaliza prin asamblarea unui complex enzimatic C5,6,7,8,9 pe suprafata celulei - tinta. Acest complex a fost denumit 'unitate de atac' a membranei, deoarece, provoaca solutii de continuitate care duc la liza celulei atacate. Mecanismul este foarte eficient deoarece o singura molecula de C'3 - convertaza poate activa sute de molecule de C'3 pe suprafata celulara. Este de subliniat faptul ca C'3 nativ din plasma este o molecula instabila. Ea genereaza continuu si aparent spontan C'3b cu o rata foarte scazuta. Aceasta autoactivare poarta riscul formarii convertazei caii
alternative si de aici pe acela al declansarii secventelor inscrise mai sus (activarea autocatalitica a lui C'3). Organismul este protejat impotriva acestei activitati potential devastatoare prin mai multi inactivatori plasmatici si celulari care actioneaza in diverse puncte ale cascadei, incepand cu dezasamblarea convertazei.
Evaluarea in clinica a C' poate fi facuta prin dozari radioimune ale componentelor sau prin aprecierea globala a activitatii sistemului, care are ca principiu evaluarea capacitatii serului de cercetat de a liza 50% dintr-o suspensie standard, de eritrocite de oaie, invelite cu anticorpi specifici. Aceasta determinare a 'complementului hemolitic' (CH50) apreciaza global activitatea caii clasice de activare si serveste ca test de triaj pentru detectarea unor anomalii mai pronuntate ale sistemului. Complementul prezinta in mare, urmatoarele actiuni:
provoaca liza antigenelor dupa fixarea pe complexele antigen- anticorp;
activeaza fagocitoza;
are actiune flagogena;
activeaza chimiotaxia;
mareste imunoaderenta;
produce conglutinare.
Lizozimul (muraminidaza) este un ferment glucidolitic (FLEMING, 1922), prezent in lichidele organice ale diverselor specii de animale, in special la mamifere si pasari si care are o actiune dezintegranta asupra celulei bacteriene. Modul de actiune consta in liza componentelor mucopeptidice (mureina) ale peretelui celulei bacteriene si consecutiv, a distrugerii acestora.
Activitatea optima a lizozimului se desfasoara la pH 7,2 si temperatura pana la 50C, in conditii de anaerobioza si aerobioza.
Raspandirea lizozimului in organism este mare, prezenta lui fiind semnalata in secretiile care umecteaza mucoasele conjunctivala, bucala, nazala si vaginala, deasemenea, se gaseste in umori ca in lichidul pleural, plasma sanguina si intr-o serie de organe, stomac, ficat, amigdale, intestin, precum si in albusul de ou. Lipseste din urina, fecale, si lichidul cerebro-spinal.
Activitatea lizozimului este mai marcata asupra bacteriilor Gram pozitive saprofite (Micrococcus, Sarcina, Staphylococcus, Bacillus) si mai putin marcata asupra Gram - negativilor.
Interferonii (IFN) reprezinta o familie de glicoproteine moleculare provenite din mai multe surse:
IFN alfa, produsi de leucocite ('IFN leucocitari');
IFN beta, produsi de fibroblasti si celulele epiteliale ('IFN fibroblastici');
IFN gamma, produsi de limfocitele T ('IFN imuni').(Fig.5)
Dupa infestarea cu un virus celulele parazitate produc si secreta IFN. Acesta se ataseaza pe receptori specifici exprimati pe membrana celulelor invecinate carora le induce o stare de rezistenta antivirala, care stanjeneste diseminarea virusurilor. Moleculele de interferon, se leaga la receptorul pentru interferon (RIFN) de pe celulele invecinate celulei infectate. Complexul IFN - RIFN este endocitar si interferonul actioneaza prin doua mecanisme pentru a impiedica infectarea acestor celule:
activarea unei gene care blocheaza sinteza proteinelor virale;
activarea unei enzime (endoribonucleaza) care fragmenteaza ARN viral.
Productia IFN apare foarte repede dupa ce s-a produs infectia. Se presupune ca celula reactioneaza fata de acizii nucleici virali prin derepresia genelor pentru IFN.
Speciile cele mai eficiente in apararea antivirala sunt IFN alfa, IFN beta. IFN gama joaca un rol important in reglarea sistemului imun: activeaza macrofagele, limfocitele B si T, celulele NK si indirect, regleaza productia de anticorpi. IFN prezinta specificitatea de specie (au activitate maxima asupra celulelor speciei care i-a produs). In prezent, IFN, care pot fi produsi in vitro prin inginerie genetica, sunt utilizati in tratamentul mai multor afectiuni neoplazice sau inflamatorii.
Polipeptidele bazice actioneaza asupra bacteriilor Gram pozitive, mecanismul fiind asemanator cu a detergentiolor cationici fara a actiona asupra lui Listeria si Pneumococcus.
Anticorpii naturali (opsonine nespecifice)
Se numesc anticorpi naturali, componentele serice care exista in serul indivizilor normali in afara tuturor imunizarilor specifice de origine infectioasa sau vaccinala. In realitate, acesti anticorpi naturali sunt produsi dupa acelasi proces ca si anticorpii imuni rezultati dintr-o imunizare specifica. Totusi trebuie mentionata existenta lor in cadrul imunitatii nespecifice din moment ce prezenta lor in umori este naturala, deci neconsecutiva unei imunizari voluntare sau postinfectioase.
Plecand de la anticorpii naturali, transmisi eventual, de la mama la nou - nascut pe cale transplacentara sau colostrala si care dispar in prima perioada de viata extrauterina, acestia sunt produsi ulterior de acelasi individ in numar si in concentratie crescuta, pentru a atinge un maxim la sfarsitul perioadei de crestere. Sinteza lor este datorata resorbtiei intestinale a antigenelor florei bacteriene normale sau alimentelor ingerate (de origine vegetala in principal). Animalele axenice, crescute in boxe sterile si private de flora intestinala nu produc acesti anticorpi decat foarte putin si apar numai ca un raspuns la antigenele de origine alimentara. Acesti anticorpi naturali sunt IgM produse de limfocitele B purtatoare a markerului CD5.
Numarul crescut de Atg. existenti in lumea vie, sta la baza cuplarii nespecifice a anticorpilor naturali din serurile normale cu bacteriile, globulele rosii, celulele animale sau vegetale ce urmeaza a fi captate prin endocitoza. Fagocitoza majoritatii Atg. necesita in prealabil, opsonizarea (sensibilizarea) lor prin intermediul Atc. naturali si a complementului (C').
Opsonizarea consta in legarea opsoninelor de Atg. solubile sau corpusculare, reactie care permite, aderenta acestora de suprafata celulelor fagocitare purtatoare de receptori Fc sau C' cu rol majoritar in inducerea fagocitozei (imunoaderenta).
Atc. naturali provoaca, deasemenea, aglutinarea Atg. bacteriene in suspensie, motiv pentru care mai sunt cunoscuti si ca aglutinine naturale. Dar puterea aglutinanta a serurilor normale se poate exercita si asupra hematiilor. Sunt calificate drept heterohemoaglutinine naturale, anticorpii serului unei specii care aglutineaza hematiile unei alte specii. De exemplu, serul de gaina contine numeroase heterohemoaglutinine pentru globulele rosii ale unor diverse specii de animale.
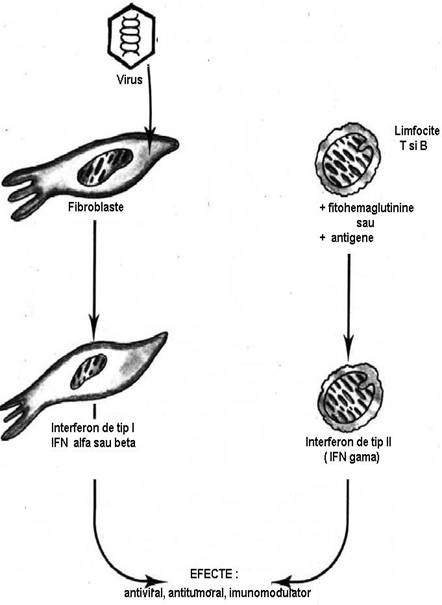
Fig.5. Tipurile de interferoni
In cazul in care globulele rosii ale anumitor indivizi sunt aglutinate de catre serul altor indivizi din aceeasi specie, se vorbeste de izohemoaglutinine. Ele au fost in mod particular demonstrate de serul uman (izohemoaglutinine contra antigenelor de grup A si B). Serul a numeroase specii de animale domestice nu contine izohemoaglutinine naturale cu exceptia celui de la caine de grup sangvin DEA - 7 (care contine izohemoaglutinine anti DEA - 7 naturale) si a celui de la pisica de grupa B care contine izohemoaglutinine anti - A.
Incrucisarile antigenice sunt deasemenea, frecvente intre bacterii si explica de ce anticorpii naturali indreptati impotriva bacteriilor banale din flora digestiva pot juca un rol benefic in mod natural si contra bacteriilor patogene inrudite antigenic.
Beta - lizinele
Este recunoscut faptul ca plachetele sangvine constituie sursa unei mici proteine bazice (6 Kd) care este eliberata atunci cand se produce agregarea lor in cursul coagularii. Aceasta proteina este stocata in granulele plachetelor sangvine. Ea s-a denumit 'B - lizina de origine plachetara' pentru a o distinge de o alta proteina diferita, care poseda aceleasi activitati, izolata din lizozomii leucocitari si cunoscuta sub numele de 'B - lizina neplachetara'. Probarea originii plachetare a beta- lizinei rezulta din constatarea ca serul, obtinut prin coagularea plasmei, nu contine beta - lizine.
Beta - lizinele sunt in egala masura gasite in saliva si in lacrimi unde prezenta lor este asigurata printr- o sinteza a celulelor secretoare ale glandelor salivare si canalului lacrimal.
Efectul lor asupra bacteriilor sensibile consta intr-o liza asupra peretelui acestora, dupa contactul cu serul activ, liza bacteriilor Gram pozitive are loc in cateva minute. Aceasta eficacitate bacteriolitica remarcabila a lizozimului si beta-lizinelor, capabila sa neutralizeze milioane de bacterii saprofite care vin in contact continuu cu tesuturile organismului la nivelul ranilor, face parte din mecanismele naturale de defensiva nespecifica.
Proteina C reactiva (PCR) isi trage numele de la faptul ca reactioneaza, printre altele, cu proteina C a pneumococului. PCR se ataseaza pe unele bacterii sau fungi si promoveaza invelirea acestora cu moleculele de C'. Acoperirea Atg. cu C' ('opsonizarea') produce facilitarea fagocitarii acestora. (Fig.7)
3. Factorii celulari
In aceasta grupa de factori sunt
incadrate toate celulele din sistemul fagocitar mononuclear si polimorfonuclear
(polimorfonucleare
neutrofile), microfage (monocite), macrofagele mobile si fixe care se gasesc
raspandite in toate organele si tesuturile, precum si celulele NK.(Fig.6)
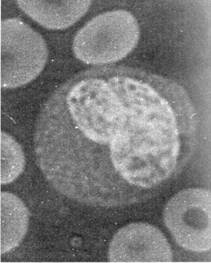
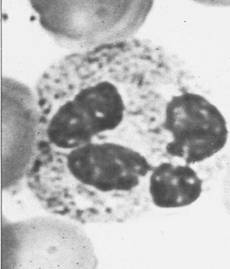
a b
Fig 6: a. Polimorfonucleare neutrofile; b. Monocite
(microscopie optica).
1. Fagocitoza si pinocitoza (gr. fagien = a manca, Mecinikov, 1905) sunt mecanisme cunoscute in cele mai fine detalii. Termenul de fagocitoza este utilizat pentru particule cu o talie mai mare si o structura de regula corpusculara (o celula bacteriana sau animala sau un fragment din acestea) iar cel de pinocitoza pentru materialele fagocitabile, al caror nivel de organizare este molecular (macromolecule proteice sau complexe, endotoxine bacteriene etc). Ambele fenomene au aceeasi dinamica, ele desfasurandu-se in patru faze :
a) faza de apropiere care presupune congestie, marginatie leucocitara si deplasarea prin pseudopode;
b) faza de fixare a materialului fagocitabil, pe fagocit, realizata pe fondul chimiotactismului determinat de complementaritatile de natura electrostatica si stereochimica dintre receptorii memebranei fagocitului si gruparile chimice de la periferia particulei fagocitabile; faza de fixare este mediata si de interventia unor factori umorali din categoria opsoninelor sau anticorpilor naturali;
Fig.7. Modele de cooperare `ntre factorii imuni
umorali [i celulari `n aprarea nespecific
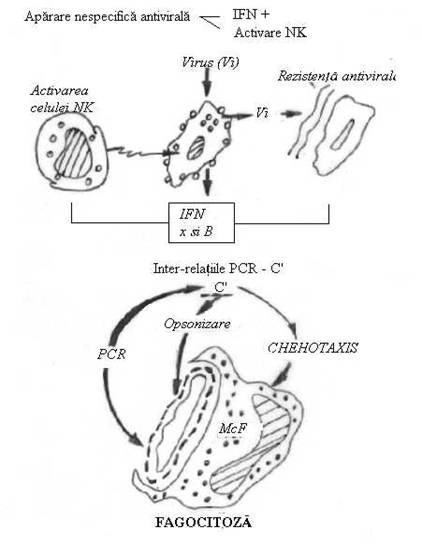
c) faza de inglobare, initiata prin formarea unei invaginari a membranei care se adanceste treptat, pana cand se inchide deasupra moleculei sau particulei, incluzand-o intr-o vacuola intracitoplasmatica, ce poarta numele de fagozom;
d) faza de digestie a matarialului fago(pino)citat, prin enzime de tip lizozomal, debuteaza prin apropierea si apoi contopirea fago(pino)zomului cu un lizozom. La nivelul acestuia se produce degradarea materialului fago(pino)citat prin procese enzimatice in care predomina reactiile oxidative si cele de hidroliza. Majoritatea enzimelor decelate in cursul digestiei intrafagocitare sunt de tipul oxidazelor, al mieloperoxidazei si a hidrolazelor acide. (Fig. 8)
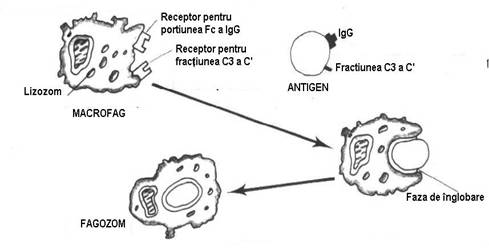
Fig. 8. Prezentarea schematica a fagocitozei
In faza de digestie se poate constata o destul de mare diversitate in comportarea materialului fagocitat in functie de starea imunologica in care este implicat, procesul de fago(pino)citoza:
digestia completa constand in degradarea materialului fago(pino)citat pana la produsi finali, este un fenomen constant, in cazul leucocitelor polimorfonucleare si frecvent in cazul macrofagelor; metabolizarea totala a particulelor ingerate este un mecanism nespecific caracteristic imunitatii naturale;
digestia partiala a non-selfului in proportie, ajungand pana la 90% , dar cu pastrarea la suprafata macrofagului a aproximativ 10% din non-self si purtarea acestuia la exteriorul celulei, timp de cateva saptamani; este un mecanism care intervine in imunitatea dobandita si in fenomenele de alergie. Acest tip de fagocitoza poate fi numita si fagocitoza imuna. Ea este un atribut exclusiv al macrofagelor iar non-selful fagocitat este antigenic. In fixarea antigenului pe suprafata macrofagului intervin si anticorpii citofili. Prezenta acestor anticorpi este confirmata si prin evidentierea receptorilor Fc la nivelul membranei macrofagelor. Elementele antigenice pastrate pe suprafata celulei sunt ulterior oferite limfocitelor, dupa unul din modelele care vor fi descrise la capitolul de imunogeneza. Caracterele diferentiale intre fagocitoza normala si cea imuna sunt rezumate in tabelul urmator:
Tabelul 2
|
Caracterul distinctiv |
Fago(pino)citoza ca mecanism al imunitatii naturale |
Fago(pino)citoza ca mecanism al imunitatii dobandite |
|
Celule implicate |
Microfagele polimorfonucleare si macrofagele |
Numai macrofagele |
|
Categoria de non-self |
Toate categoriile |
Numai antigenele |
|
Factorii care faciliteaza fixarea pe celula |
Numai opsoninele (anticorpii naturali) |
si anticorpii citofili fixati pe receptorii Fc ai macrofagului |
|
Tipul de digestie |
Totala |
Partiala |
|
Soarta materialului nedegradat |
Este eliminat |
Este transmis limfocitelor |
persistenta nealterata si mentinerea viabilitatii unor bacterii patogene; ex. clasic este cel al bacililor tuberculozei, care nu numai ca nu sunt digerati, dar raman vii, se multiplica, determina transformarea fagocitelor in celule gigante, iar acestea devin un mijloc de vehiculare a germenilor in organism;
digestia unor tipuri de 'non-self' pe care fagocitele organismelor neimunizate in mod normal nu le digera. In organismul animalelor cu imunitate antituberculoasa apar macrofagele capabile sa digere bacilii tuberculozei; aceste fagocite au prin urmare o comportare de sens contrar cu cele mentionate anterior. Noua insusire dobandita de aceste macrofage, numite acum imunofagocite, poate fi considerata ca un mecanism al imunitatii dobandite, cu toate ca gradul de specificitate al acestei transformari calitative celulare este redus. Imunofagocitele obtinute cu bacilii tuberculozei manifesta aceleasi proprietati si fata de brucele, salmonele sau listerii, germeni caracterizati si ei prin rezistenta la digestia intrafagocitara.
Pinocitoza se deosebeste de fenomenul de fagocitoza prin cateva caractere diferentiale (MESROBEANU si BONA, 1968) :
a) In timp ce inductorul de pinocitoza este solubil cel al fagocitozei este intotdeauna o particula solida.
b) Inductorul de pinocitoza trebuie sa posede o anumita incarcatura electrica, pe cand corpusculul fagocitat poate fi neutru.
c) Pinocitoza, spre deosebire de fagocitoza nu implica chemotactism.
d) Aspectul citologic al celor doua fenomene difera: in fagocitoza apar modificari ale membranei celulare sub forma de pseudopode, in pinocitoza apar invaginari, vezicule sau canalicule ale suprafetei celulare.
Incorporarea particulelor virale, printr-un fenomen similar pinocitozei se numeste viropexie.
Celulele NK ('natural killer cells') recunosc modificari de suprafata care apar la celulele infestate cu virusuri. Celulele NK sunt activate de IFN produs de acele celule pe care le ucid printr-un mecanism de citotoxicitate directa. Sunt asemanatoare cu limfocitele, dar au talia mai mare si granulatiile azurofile in citoplasma ('large granular lymphocytes')- LGL. Dupa infectia virala, celulele infestate sunt induse sa exprime pe suprafata o serie de structuri glicoproteice macromoleculare care ar fi 'recunoscute' de celulele NK cu ajutorul unor receptori. Dupa alipirea la celula-tinta are loc eliberarea mai multor proteine active din granulatiile celulelor NK. Cea mai importanta dintre aceste substante este 'perforina' (citolizina) care-si trage numele de la faptul ca produce pori in membrana celulei-tinta, intocmai ca si componenta a 9-a a C', cu care de altfel, are unele analogii structurale.
Perforinele se ataseaza pe membrana celulei tinta si in prezenta ionilor de Ca2+ vor polimeriza si vor forma canale care strabat membrana tintei de la exterior spre interior. Se creeaza o diferenta de presiune intre interiorul si exteriorul celulei ceea ce face ca prin aceste canale sa intre apa si ioni de Na+ , celula tinta fiind in cele din urma lizata.
Celulele NK au pe suprafata membranei un proteoglican numit CONDROITIN SULFAT A, care protejeaza celulele NK de activitatea perforinelor. In concluzie, celula NK indeplineste urmatorul rol:
A. celula NK ucide:
celulele infectate cu virus;
celulele tumorale
are rol in supravegherea hematopoezei deoarece distruge celulele "defecte" rezultate din hematopoeza;
B. celula NK are rol in reglarea raspunsului imun prin secretia de citokine (interleukina 2 si interferon gamma) care intervin in cooperarile intercelulare.
4. Modificarea imunitatii naturale
Imunitatea naturala nespecifica poate fi modificata prin mecanisme complexe fiziologice, fiziopatologice si de factorii constitutionali si genetici.
Temperatura normala circulatia sanguina si limfatica si presiunea sanguina contribuie la activitatea normala a imunitatii umorale si celulare nespecifice.
Temperatura scazuta, diminuarea circulatiei sanguine si limfatice duc la scaderea indicelui opsono-citofagic (a fagocitozei), la lipsa de oxigenare si nutritia tesutului.
Hipertermia sau reactia febrila are un rol benefic in apararea nespecifica naturala, prin activarea circulatiei periferice si centrale, cresterea catabolismului celular si a oxido-reducerilor celulare in urma unor activitati intense a hormonilor tiroidieni si steroizi. La acestea trebuie adaugat si contributia reactiei febrile la mobilizarea celulelor implicate in procesul de fagocitoza si a activarii factorilor serici ai reactivitatii nespecifice (lizozimul, complementul, interferonii, etc). Hipertermia poate actiona si direct asupra unor virusuri 'reci' care nu suporta temperaturi mai mari de 38,5C. Principalele substante piretogene (lipopolizaharidele bacteriene, amidele, endotoxinele, interleukina 1. etc) actioneaza asupra centrului de termoreglare din hipotalamus ducand la hipertermie.
Reactia inflamatorie prin efectele ei, mecanismele si caracterele histologice precum si evolutiile ulterioare poate influenta imunitatea naturala nespecifica.
Manifestarea mai aparenta sau mai putin aparenta a unui proces infectios, semnifica puterea de aparare a organismului respectiv.
Inflamatia reprezinta o reactie locala a organsimului fata de invazia microorganismelor in tesuturi. Ea limiteaza diseminarea germenilor si reduce considerabil mortalitatea prin septicemie.
Clinic, procesul inflamator local se caracterizeaza prin congestie (rubor), edem (tumor), caldura (calor), durere (dolor) si tulburari functionale (functiolesa).
Microorgansimele patrunse intr-un tesut, produc la poarta de intrare o alterare initiala atat prin prezenta lor fizica cat si prin produsele lor metabolice. In aceasta alterare initiala, (incetinirea circulatiei, vasodilatatiei, cresterea permeabilitatii vasculare, diapedeza leucocitelor etc), un rol important joaca histaminele care determina aceste modificari. Granulatiile din mastocite, bazofile, eozinofile, neutrofile (celule specializate care insotesc inflamatia) contin in majoritate histamina si alti principii activi: heparina, serotonina, bradichinina, leucotriene, tromboxani, prostaglandine etc.
In continuare, exudatul inflamator se imbogateste in diversi principii activi (leucotoxine si factorul de initiere a leucocitozei -LPF), care isi au originea in celulele care participa la inflamatie (limfocite, plasmocite, histiocite).
Sistemul nervos intervine deasemenea in mecanismele de vasodilatatie prin reflexul de axon pus in evidenta de stimularea capatului periferic a unui nerv sectionat, care determina o vasodilatatie in teritoriul inervat de acesta.
Fibrele simpatice stimuleaza franarea reactiei inflamatorii, iar simpatectomia intensifica vasodilatatia.
Evolutia ulterioara a inflamatiei in functiei de agresivitatea agentului patogen duce la constituirea focarului inflamator si la localizarea infectiei.
Cantitatea crescuta de acid lactic din focarul inflamator, consecinta gliucolizei anaerobe, este susceptibila de exercitarea unei actiuni nocive printr-un pH scazut, asupra diversilor agenti infectiosi (ex. bacilul tuberculozei).
Factorii care modifica imunitatea naturala nespecifica sunt:
a) factorii diminuanti: neintegritatea barierelor anatomice, anastezicile, iradiatiile, imunodepresoarele si unele boli ca: silicoza, diabetul, obezitatea, etilismul cronic, etc.
b) factorii favorizanti: endotoxinele bacteriene, micobacteriile (M. avium, M. kansasii), BCG, Corynebacterium parvum, poliadjuvantul din peptidoglicani si adjuvantul complet Freund.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4296
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved