| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Imunologie
Antigen Presenting Cells (APC; celulele prezentatoare de antigen) sunt celule cu rol esential in generarea unor raspunsuri imune umorale si celulare fata de antigenele timodependente. Au trei functii importante din punct de vedere imunologic:
sunt responsabile direct de declansarea raspunsului imun;
au rol esential in intretinerea (in timp) a raspunsului imun;
participa la controlul raspunsului imun.
APC sunt implicate intr-un fenomen de cooperare celulara cu doua categorii de limfocite:
I. Limfocite cu rol imunoreglator, clasificate in doua subvarietati:
a. T-helper cu rol in amplificarea raspunsului imun;
b. T-supressor cu rol in inhibare raspunsului imun.
II. Limfocite cu rol efector (care participa la realizarea efectiva a raspunsului imun):
a. Limfocite B pentru raspunsul imun umoral;
b. Limfocite T citotoxice pentru raspunsul imun celular.
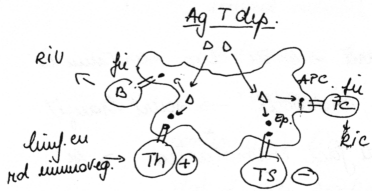 (fig. 1)
(fig. 1)
Legenda pentru figura 1: Antigenele timodependente sunt preluate de APC, apoi internalizate si prelucrate. In urma acestei prelucrari rezulta fragmente antigenice (epitopi) care vor fi expuse pe membrana APC in complex cu molecule MHC (n. ed. MHC tip II; Major Histocompatibility Complex type II). Sub aceasta forma epitopii sunt <prezentati> ambelor subvarietati de limfocite cu rol imunoreglator (Th, Ts), apoi epitopii sunt prezentati unei singure subvarietati de limfocite cu rol efector, fie B, fie T (n. ed. nu sunt prezentati ambelor subvarietati).
Celulele APC au urmatoarele caracteristici:
a) dintre toate celulele participante la cooperare, APC sunt primele si singurele celule ce vin in contact cu antigenele (Ag) native;
b) sunt singurele capabile sa prelucreze Ag native generand fragmente antigenice (epitopi); doar aceste fragmente sunt recunoscute de limfocite (si in special de limfocitele T);
c) APC sunt substratul morfologic al cooperarii celulare.
Clasificarea APC:
I. APC profesionale:
macrofage;
celule dendritice;
limfocite B (in anumite conditii pot functiona ca APC).
II. APC ocazionale:
PMN (dintre acestea sunt mai importante neutrofilele);
Celulele endoteliale;
Celulele epiteliale.
I.1 APC profesionale - macrofagele
Macrofagele sunt celule de origine medulara; maduva (n. ed. osoasa hematopoietica) elimina continuu in circulatia periferica elemente (n.ed. figurate sanguine) tinere, nematurizate (monocite). Dupa un scurt tranzit (24.36 h), acestea trec in tesuturi, fiind atrase de anumiti factori (chemotactici). In tesuturi se vor maturiza sub influenta unor (n. ed. altii?) factori cum ar fi IFN g (n. ed. gamainterferon), transformandu-se treptat in macrofage. Macrofagele pot indeplini functia de APC (trecand prin anumite transformari).
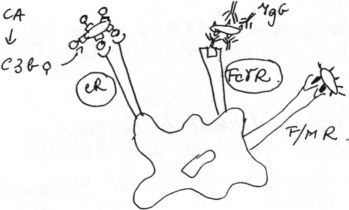 (fig.
2)
(fig.
2)
Transformarea macrofagelor in APC a fost impartita (n. ed. didactic) in 5 etape (n. ed notate A.E in cursul de fata).
A. Captarea antigenelor:
A.1 Antigenele corpusculare captate de macrofage pot fi impartite in doua categorii:
i) Antigene corpusculare opsonizate (adica <invelite> de factori imunologici solubili); opsonizarea favorizeaza net captarea Ag si endocitarea lor. Opsoninele sunt reprezentate de:
I) C3b - prezent in mod continuu in lichidul interstitial, in cantitate mica; generarea C3b se face prin functie continua a caii alternative de activare a complementului; C3b are o foarte pronuntata tendinta de fixare a oricaror suprafete membranare non-self (n. ed. adica C3b, considerat nespecific, se leaga specific si preferential de structuri non-self.
Macrofagele au receptori capabili sa lege C3b; acesti receptori se vor nota cu CR (Complement Receptor).
II) Ig (mai ales IgG) - prezente in lichidul interstitial in concentratii mai mari. Ig se ataseaza pe membrana bacteriana datorita recunoasterii unor antigene de suprafata exprimate pe respectiva membrana. Portiunea Ig care se leaga la determinantul antigenic a fost notata Fab (Fragment antigen-binding). Fragmentul Fc (Fragment constant) al Ig astfel legate este liber in lichidul interstitial. Captarea are loc rapid prin legarea Fc la receptorii FcgR ai macrofagelor (n. ed. spre deosebire de complement, Ig sunt specifice in ceea ce priveste Ag legat prin structura Fab, care este complementara cu structura antigenica recunoscuta; pentru doua antigene diferite, Fab corespunzator nu poate fi acelasi; Fc este acelasi indiferent de antigenul recunoscut de Fab).
ii) Antigene corpusculare neopsonizate - captarea lor este mai putin eficienta. Macrofagele au receptori care se leaga la oligozaharide ale peretelui bacterian - radicalii fucozil, manozil - de unde si notatia F/MR.
A.2 Captarea este nespecifica pentru ca macrofagele nu au contact direct cu determinantii antigenici de suprafata, contactul facandu-se (n.ed. exclusiv) prin C3b, Fc si radicali F/M (n.ed. care sunt structuri constante);
B. Endocitarea antigenelor corpusculare captate este un proces energodependent (<activ>); endocitarea presupune modificarea conformatiei membranare cu formare de pseudopode si vezicule de endocitare.
C. Prelucrarea/procesarea antigenelor endocitate. In lizozomi, sub actiunea proteazelor si hidrolazelor are loc o proteoliza partiala in urma careia rezulta fragmente (epitopi). Ulterior sunt selectati epitopii imunodominanti (cei mai imunogeni).
D. O parte din epitopii selectionati sunt conservati in macrofag si servesc la intretinerea in timp a raspunsului imun. Cantitati mici sunt eliberate si mentin stimularea antigenica.
E. Cea mai mare parte din epitopii selectati se cupleaza in macrofag cu molecule MHC I sau MHC II, rezultand complexe MHC I (sau II) - epitop care vor fi expuse pe membrana macrofagului. Primele celule care vin la cooperare sunt limfocitele T (mai exact subsetul Th). Limfocitele Th pot recunoaste epitopii doar in complexe MHC II - epitop. Capacitatea de prezentare este deci conditionata de capacitatea macrofagelor de a sintetiza molecule de tip MHC II. De exemplu:
70 % din macrofagele peritoneale sunt capabile sa sintetizeze MHC II;
50 % din macrofagele splenice sunt capabile sa elaboreze MHC II;
maxim 25 % din macrofagele hepatice sunt capabile sa exprime MHC II.
Macrofagele care elaboreaza MHC II sunt implicate in generarea unor raspunsuri imune. Cele care nu au aceasta capacitate nu participa la generarea unor raspunsuri imune dar sunt implicate doar in reactiile inflamatorii locale.
Etapele limitante ale procesului de transformare macrofag-APC sunt A si E:
rata captarii depinde de numarul de receptori CR, FcgR functionali;
prezentarea depinde de capacitatea de sinteza a MHC II.
Functia de ansamblu a macrofagelor in raspunsul imun: capteaza antigenele si le transporta catre ganglionii locoregionali unde le prezinta limfocitelor Th aflate in aria timodependenta a ganglionilor (paracortexul ganglionar).
I.2 Celulele dendritice (n. ed. in cadrul sectiunii I.2 notatia va fi CD)
.sunt celule de origine medulara care in conditii de repaus emit prelungiri citoplasmatice (pseudopode). Ansamblul pseudopodelor are urmatoarele caracteristici:
pseudopodele sunt foarte numeroase;
sunt foarte fine, subtiri, efilate;
sunt foarte lungi (lungimea minima este de 5.6 ori diametrul celular).
Aceste caracteristici dau celulei un aspect similar celor dendritice din SNC. In ceea ce priveste functia APC, despre celulele dendritice se poate afirma ca:
a. in general exprima receptori pentru opsonine, deci pot capta antigene;
b. in general pot sintetiza MHC II.
Celulele dendritice (CD) sunt o clasa foarte heterogena, cu sase subtipuri, grupate in trei subunitati mai importante (n. ed. notate I.III in tabelul de mai jos).
Grupa/Subtip CD
|
I. CD intratisulare (nelimfoide) 1. cel. Langerhans |
CR |
FcgR |
MHC II |
|
2. CD interstitiale | |||
|
II. CD limfoide 3. IDC | |||
|
4. CD foliculare* | |||
|
III. CD intracirculatorii 5. CD sanguine | |||
|
6. CD valuroase |
* CD foliculare functioneaza ca filtre locale pentru antigene.
Grupa I
1. CD din subtipul 1 se gasesc:
in submucoasa bucala;
tractul digestiv;
colul uterin.
Functia lor de ansamblu este de a capta antigenele si a le transporta in ganglionii locoregionali, in paracortexul acestora prezentandu-le limfocitelor Th.
2. CD interstitiale au morfologie similara celulelor Langerhans. Sunt foarte larg distribuite (se gasesc in toate spatiile interstitiale).
Grupa II este formata din celule localizate in organele limfoide secundare.
3. CD interdigitate (engl. IDC) au aspect tipic dendritic si sunt distribuite in ariile timodependente din ganglionii limfatici si splina:
paracortex ganglionar;
pars interna a splinei (strat limfoid periarteriolar intern).
4. CD foliculare au aspect tipic dendritic si sunt distribuite in foliculii limfoizi din organele limfoide secundare (foliculi limfoizi= aglomerari de limfocite B), adica in ariile bursodependente din organele limfoide:
cortexul ganglionar;
pars externa a splinei.
CD intracirculatorii
5. CD sanguine - ca morfologie sunt ovalare (in circulatie). Sunt intalnite rar.
6. CD valuroase (engl. veiled) sunt aplatizate si ovalare. Pe lateral prezinta foarte multe franjuri si ondulatii. Sunt prezente numai in circulatia limfatica, mai ales in limfaticele aferente. Fenotipic sunt CR (-), FcgR (-), MHC II (-).
Se considera ca toate aceste celule dendritice (1.6) nu reprezinta entitati morfologice distincte, ci sunt principalele etape ale traficului celulelor dendritice in organism. Toate CD au aceeasi origine (n.ed maduva osoasa hematogena) si in functie de tesutul pe care il parcurg isi modifica caracterele morfologice si fenotipice.
Traficul si filiatia celulelor dendritice in organism se desfasoara in patru sectoare: maduva, sectorul tisular, sectorul limfoid si sectorul vascular.
CDS I. Calea intratisulara:
a. in tesutul cutanat devin celule Langerhans: CR(+), FcgR(+); se pot activa, caz in care transporta antigenele spre ganglionii locoregionali prin limfaticele aferente, unde devin CD valuroase (CDV); in paracortexul ganglionar aceste CDV devin IDC, care sintetizeaza in cantitati mari MHC II prin care prezinta antigenele catre limfocitele T helper (3000.4000 Th: 1 IDC) inclavate printre prelungirile IDC.
b. in spatiile interstitiale din organele limfatice devin CD interstitiale, care exprima CR si FcgR; prin activare migreaza si vor transporta antigenele pe doua cai:
i. catre ganglionii locoregionali - CDV- IDC (in maniera descrisa mai sus la litera a.);
ii. migreaza antidromic - in circulatie redevin CDS, trec in splina devenind IDC in PALS interna; IDC coopereaza cu limfocite T.
II. Traficul limfoid presupune trecerea directa:
i. in ganglioni apoi in cortexul ganglionar, unde are loc transformarea in CDF;
ii. prin artera lienala in PALS externa cu transformare in CDF.
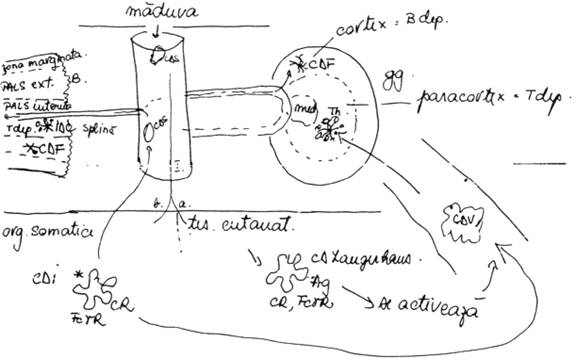
(fig. 3)
I.3 Limfocitele B pot functiona ca APC deoarece exprima receptori de captare a antigenelor (CR, FcgR, BCR) (n. ed. aceasta este formularea data in curs si trebuie sa-i ramanem fideli, chiar daca este in mod clar un sofism; e.g. daca eu am o undita nu inseamna ca sunt pescar). Limfocitele B capteaza doar antigenele solubile. In stadiul de limfoblast, ele elaboreaza cantitati crescute de molecule MHC II.
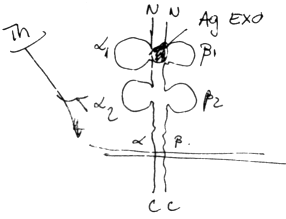
Moleculele prezentatoare de antigen (MHC)
MHC I sunt distribuite ubicuitar, cu exceptia hematiilor. Ca structura sunt heterodimeri ab. Lantul a are o portiune transmembranara pe cand b este strict extracelular. Aceste lanturi prezinta organizare multidomeniala datorita unor legaturi disulfurice intracatenare. Lantul a are trei domenii a a iar lantul b are o singura bucla:
domeniile a a sunt constituite din secvente hipervariabile (difera de la un tip celular la altul) si de aceea sunt numite domenii polimorfe ale MHC I;
domeniul a are o structura primara constanta fata de tipul celulei (n.ed independenta de tipul celulei pe a carei membrana este exprimata molecula MHC I analizata);
domeniul b are o structura primara constanta la toti indivizii unei specii date
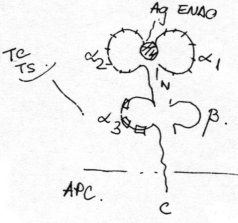 (fig.
4)
(fig.
4)
Functii: Domeniile a a sunt situate simetric si se asociaza formand o cavitate de dimensiuni reduse in care este inclavat un antigen (= antigen endogen) = situs combinativ pentru antigene. Domeniul a se comporta ca un antigen self foarte imunogen, fiind implicat in declansarea unor reactii de respingere a grefelor de organ/ tesut.
Gena MHC I este situata pe autozomii 6.
MHC II sunt distribuite doar pe membranele celulelor ce participa la realizarea raspunsului imun: APC, limfocite B si limfocite T. Ca structura sunt heterodimeri ab, ambele lanturi fiind transmembranare. Fiecare lant prezinta cate doua domenii (<bucle>) iar pozitionarea lor simetric fata in fata da moleculei un aspect Ig-like.
Domeniile aminoterminale a b sunt domeniile polimorfe ale moleculei MHC II (variaza mult de la o clona celulara la alta). Domeniile carboxiterminale a b au structuri primare constante (mai ales a ) si in consecinta se numesc domenii monomorfe.
Domeniile polimorfe se asociaza formand cavitati in care este inclavat pentru prezentare un antigen exogen= situs prezentator pentru antigen.
Domeniile monomorfe (mai ales a ) au rol in declansarea unor reactii de respingere a grefelor si in cooperarea APC-Th.
+i genele MHC II sunt situate pe perechea 6 de autozomi.
Mecanismele prezentarii selective a antigenelor pe moleculele MHC: desi ambele categorii de molecule MHC sunt sintetizate in acelasi subsector celular (RER), ele prezinta diferentiat antigenele pentru ca se cupleaza cu acestea in subsectoare celulare diferite.
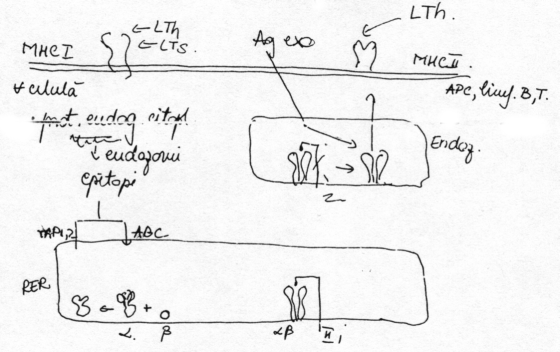
In cazul MHC I se sintetizeaza initial lantul b apoi a. Prin cuplarea acestora rezulta molecula MHC I si are loc modificarea conformationala a lantului a, cu deschiderea larga a situsului combinativ pentru antigen. Astfel, moleculele MHC I pot fixa diverse antigene: proteine endogene intracitoplasmatice - autoantigene sau neoantigene sau fragmente rezultate din prelucrarea unor microorganisme cu habitat sau dezvoltare intracelulara (n.ed. in acest al treilea caz, nu se poate afirma ca respectivele proteine au provenienta endogena, nefiind biosintetizate pe o matrita ARN provenita din transcrierea unei secvente ADN proprii). Prin prelucrarea respectivelor polipeptide in citoplasma, sub influenta proteozomilor (n. ed. lizozomi?) rezulta epitopi care prin anumite sisteme transportoare (vezi figura mai sus: TAP 1,2= transportor asociat prezentarii de antigen; ABC= casetele de legare a antigenului) ajung in RER (reticul endoplasmic rugos). Incarcarea moleculelor MHC I cu epitopi implica o noua modificare conformationala, MHC I desprinzandu-se de pe perete si migrand pe membrana.
In cazul MHC II se sintetizeaza pe langa lanturile a si b si un lant II inhibitor (IIi) care <astupa> (blocheaza) situsul de prezentare a antigenului. Lantul IIi trece printr-un dublu clivaj proteolitic:
indepartarea capatului aminoterminal elibereaza situsul combinativ pentru antigen, care poate fixa diverse antigene (e.g. antigene exogene din procesele de fagocitare in endozomi)
al 2-lea clivaj indeparteaza complet lantul IIi iar moleculele MHC II devin neancorate pe peretii endozomali, trecand pe suprafata membranara, unde pot fi recunoscute de limfocitele Th.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2387
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved