| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Unitatea functionala a sistemului nervos este reflexul realizat prin neuron. Un neuron este format din corp (soma) sau pericarion si prelungiri denumite dendrite si axoni (cilindracsi). Axonii sunt prelungirile prin care excitatia pleaca, iar dendritele, acelea care vin la som. Forma corpului celular poate fi stelata, rotunda, piramidala, fuziforma, ovalara, piriforma etc.
Dupa numarul prelungirilor pe care le prezinta si dupa felul in care pornesc acestea din corpul celulei nervoase neuronii se clasifica in neuroni unipolari, pseudounipolari, bipolari si multipolari. Neuronii unipolari nu prezinta decat axonul, polul receptiv fiind difuz, excitatiile fiind receptionate prin toata suprafata pericarionului. Astfel de neuroni sunt rari si intra in structura retinei (celule amacrine).
Neuronii pseudounipolari sunt caracterizati prin aceea ca din corpul lor pleaca o singura prelungire, dar dupa un traiect scurt se bifurca in doua ramuri ce reprezinta dendrita si axonul; se gasesc in ganglionii spinali. Neuronii bipolari prezinta un axon si o singura dendrita care pleaca de obicei din puncte opuse.
Astfel de neuroni se gasesc in retina, mucoasa olfactiva, in ganglionii lui Corti si Scarpa. Neuronii multipolari prezinta un axon si numeroase dendrite care pornesc de pe toata suprafata pericarionului. Majoritatea neuronilor de acest tip se gasesc in toate segmentele sistemului nervos: celulele piramidale din cortexul cerebral, celulele radicaluare motorii din coarnele anterioare ale maduvei spinarii, celulele din ganglionii vegetativi extranevraxiali.
Din punct de vedere functional neuronii pot fi motori, senzitivi si de asociatie. Neuronii motori sau efectori, de obicei sunt mari, multipolari, cu axonul lung ce se termina in organele efectoare (muschi, glande). Asa sunt celulele piramidale din scoarta cerebrala, motoneuronii din coarnele anterioare medulare, celule Purkinje din scoarta cerebeloasa. Neuronii motori pot somatomotori si visceromotori.
Neuronii senzitivi sau receptori au proprietatea de a primi excitatiile receptionate de celulele senzoriale din mediul extern sau intern. Dendritele acestor neuroni senzitivi se distribuie la diferiti receptori externi sau interni. Exemple de astfel de neuroni sunt neuronii pseudounipolari senzitivi, somatici si vegetativi din ganglionul spinal, neuronii senzitivi din coarnele posterioare ale maduvei sau neuronii din ganglionii Corti si Scarpa etc. Neuronii de asociatie (intercalari), fac legatura intre neuronul senzitiv si cel motor. Sunt de dimensiuni mici, multipolari si se gasesc in toate etajele sistemului nervos.
Atat prelungiri care aduc impulsuri la soma neuronala, cat si cele care le transmit la aceasta formeaza fibrele nervoase. Ele pot avea un diametru mai mare sau mai mic, pot fi mielinizate sau nemielinizate si conduc impulsul cu o viteza in functie de aceste particularitati. Fibrele nervoase intra in componenta nervilor si pe baza caracteristicilor mentionate au fost impartite in diferite categorii (vezi fig. 1).
|
Tipul de fibre |
Diametrul Fibrei m |
Grosimea |
Viteza de conducere m/s |
Originea si functia |
|
|
A |
a |
groase |
Proprioceptori, terminatii anulo-spirale (I a), organul tendinos Golgi (Ib) |
||
|
b |
Subtiri |
Fibre pentru contact si presiune |
|||
|
g |
Subtiri |
Fibre g - eferente ale fusului muscular |
|||
|
d |
Subtiri |
Receptori termici si nociceptivi (III) |
|||
|
B |
Mai subtiri |
Fibre preganglionare vegetative |
|||
|
C |
s |
Foarte subtiri |
Fibre eferente postganglionare simpatice |
||
|
d |
|
Foarte subtiri |
Fibre eferente din radacina doprsala, pentru durere (IV) |
||
Figura 1 - Clasificare fibrelor nervoase. In paranteza e trecuta
clasificarea
numerica pentru fibrele senzoriale
Proprietatile specifice neuronului sunt excitabilitatea si conductibilitatea, proprietati generale ale tuturor tesuturilor vii.
Excitabilitatea
Excitabilitatea este proprietatea celulelor de a raspunde la un stimul. Este o proprietate generala a materiei vii. Nervii, care sunt pachete de axoni si dendrite, sunt dotati in consecinta cu excitabilitate. Excitabilitatea a fost studiata cu ajutorul stimulilor electrici care pot fi usor dozati ca intensitate si timp. In acest scop se folosesc curentii continui (galvanici) sau alternativi. Excitabilitatea a fost caracterizata prin determinarea unor raporturi intre stimul si raspuns. Cei mai importanti parametri ai excitabilitatii astfel studiate sunt: reobaza, timpul util, cronaxia, climaliza si labilitatea.
Reobaza (curent de baza, de prag sau liminar) este intesitatea minima de curent care produce un raspuns. Prin reobaza se stabileste pragul excitabilitatii sau intensitatea curentului liminar (fig. 2).
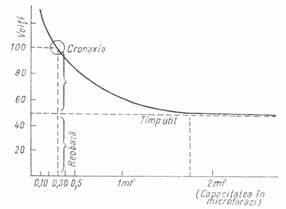
Figura 2 - Curba excitabilitatii in functie de timp
Ea poate fi, ca si alti parametri, urmarita pe un preparat neuromuscular izolat, pe un nerv izolat sau pe organismul intreg, raspunsul fiind apreciat dupa contractia musculara. In acest din urma scop se stimuleaza pe suprafata cutanata puncte situate in apropierea unui camp motor. O valoare mica a reobazei indica o excitabilitate crescuta si invers. Reobaza nervilor care inerveaza flexorii (muschii albi) este mai mica decat a extensorilor (muschii rosii), primii avand deci o excitabilitate mai mare.
Timpul util este timpul minim necesar ca un curent de intensitatea reobazei sa provoace un raspuns. El prezinta insa variatii mari (2-20 ms) la modificari mici de curent si, in consecinta, are o valoare informativa redusa.
Cronaxia este timpul minim necesar pentru ca un curent dublu fata de reobaza sa produca un raspuns din partea tesutului, respectiv a nervului. Pentru caracterizarea excitabilitatii, determinarea cronaxiei este mai valoroasa decat a reobazei si a timpului util, pentru ca valoarea ei variaza putin la modificari mai mari de curent asa cum rezulta din curba excitabilitatii in functie de timp, stabilita de Lapicque, care a si introdus aceasta notiune. Valoarea cronaxiei este de 10-30 de ori mai mica decat a timpului util, variind intre 0,1-0,6 ms.
Nervii muschilor albi, ca si acestia din urma, au o cronaxie mai mica si, in consecinta, o excitabilitate mai mare decat cei rosii. Acelasi lucru rezulta si din reobaza mai mica pe care o au primii. In trasmiterea sinaptica s-a atribuit o mare importanta egalitatii dintre cronaxia nervilor si a muschilor corespunzatori. Lapicque a presupus ca transmiterea excitatiei de la nerv la muschi ar fi conditionata de acest izocronism. Cronaxia lor are intr-adevar o valoare apropiata. Daca diferenta dintre ele are o valoare dubla (heterocronism), transmiterea impulsului nervos nu mai are loc. Se presupune ca oboseala musculara s-ar putea datora si acestui heterocronism.
Climaliza este alt parametru al excitabilitatii si exprima raportul dintre bruschetea cu care se modifica intensitatea curentului si excitatie. Pentru a se obtine un raspuns la o anumita valoare a stimulului, de exemplu a reobazei intensitatea curentului prag trebuie atinsa cu o anumita viteza. Daca ea creste incet, nervul se acomodeaza si raspunsul nu mai are loc. Excitarea liminara a nervului se obtine deci, cu o anumita bruschete minima. La acelasi curent, daca intensitatea liminara este atinsa mai repede raspunsul apare mai prompt. Timpul de climaliza (timpul de crestere a curentului pentru obtinerea excitatiei de prag) este de 2-3 ori mai scurt decat cronaxia.
Perioada refractara consta din inexcitabilitatea nervului dupa excitatie, caracterizand si alte tesuturi. Ea a fost studiata insa, mai tardiv pe nerv. Perioada refractara se imparte in absoluta si relativa. In prima faza a perioadei refractere (0,5 ms), nervul nu mai raspunde la excitatie, iar in a doua faza (2 ms), raspunde doar la un stimul mai puternic, nervul avand deci un prag de excitabilitate mai mare. Faza de inexcitabilitate absoluta a nervului corespunde cu durata potentialului de varf. Cea mai scurta perioada refractara o au nervii oculomotori. Faza de inexcitabilitate relativa este urmata de o faza de hiperexcitabilitatea.
Labilitatea este in functie de perioada refractara si exprima numarul maxim de stimul care pot fi insusiti de un tesut, respectiv de nerv. Daca frecventa stimulilor este mai mare decat labilitatea, o serie de stimuli sunt pierduti. Optimul unei excitatii consta din stimuli de frecventa labilitatii. Labilitatea diferitilor axoni variaza de la 100 la nivelul placii motorii pana la 1000 la neuronul cortical. Cea mai mare labilitate (1500) o au celulele Renshaw din maduva spinarii.
Parabioza reprezinta o modalitate particulara de raspuns a neuronilor atunci cand asupra acestora actioneaza stimuli care depasesc capacitatea lor functionala. Excitantii puternici pot determina, mai intai, o stare de excitatie, urmand apoi o depolarizare intensa, modificand in felul acesta excitabilitatea nervului care poate deveni hipoexcitabil, chiar inexcitabil. Parabioza se desfasoara in mai multe faze (hiperexcitabilitate, egalizare, paradaxola, inhibitie completa) si este considerata ca un mecanism de aparare a neuronilor fata de actiunea unor excitanti foarte puternici, ea fiind de fapt o inhibitie de protectie.
Procesul parabiotic este reversibil, dupa incetarea excitantului excitabilitatea nervoasa revine la normal, parcurgand in sens invers fazele prin care a trecut.
Conductibilitatea
Conductibilitatea nervilor consta in proprietatea lor de a transmite un impuls generat intr-un anumit punct. Impulsul este transmis in ambele directii, asa cum reiese din inregistrarea curentilor de actiune. Conducerea bilaterala este astfel una din proprietatile fibrei nervoase; dar in organismul intreg impulsul nervos este transmis intr-un singur sens datorita polarizarii sale la nivelul sinapselor din sistemul nervos central.
Conducerea unilaterala in sensul unui arc reflex este denumita ortodromica, spre deosebire de conducerea in sens invers, adica antridromica. Transmiterea liniara a excitatiei se datoreste depolarizarii zonei excitate. Transmiterea retrograda este impiedicata de perioada refractara care se instaleaza dupa excitatie. Daca se plaseaza doi electrozi pe o fibra nervoasa excitata, ca si pe alt tesut muscular liniar, se inregistreaza un curent de acaiune bifazic.
Viteza de transmitere a excitatiei este in functie de dimensiunile fibrei si de mielinizarea nervului. Este mai mare in fibrele mai groase si in cele mielinizate (de 50 de ori). Dupa viteza de transmitere a excitatiei, ca si dupa grosimea lor, nervii sunt impartiti in diferite categorii (fig. 1).
Viteza mare din fibra mielinizata se datoreste transmisterii saltatorii a impulsului de la o strangulatie Ranvier la alta, singurele zone excitabile in aceste conditii. De la o strangulatie Ranvier la alta, stimulul este transmis prin mediu, care este un bun conductor datorita compozitiei electrolitice cu proprietati de conducere.
Stimulul sare intervalul dintre cele doua strangulatii, mielina fiind un izolator relativ (fig. 3a). Se activeaza astfel, nodulul lui Ranvier. In axonii amielinici, excitatia este transmisa din aproape in aproape, prin depolarizarea membranei prin curenti locali Hermann (fig. 3b), ceea ce explica viteza mai lenta de raspandire.
Conducerea influxului dealungul unui axon integru se face izolat (legea conducerii izolate). Prin lezarea axonului excitatia este transmisa unui axon vecin.
Degenerescenta walleriana, descrisa de Waller in secolul trecut, consta in degenerarea nervului izolat prin sectiune de soma, deci a segmentului distal (anterograd). Suturand capatul perific al nervului cu cel central, el se regenereaza datorita functiei trofice a tecii fibroase care il inconjoara. Aceasta regenerare se poate produce si daca segmentul distal este suturat cu un alt nerv, ceea ce demonstreaza plasticitatea sistemului nervos central.
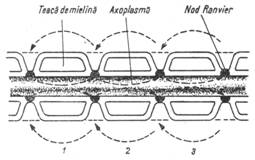
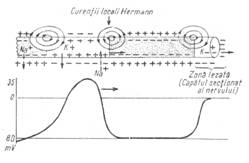
Figura 3a -
Conducerea saltatoare Figura 3b - Curentii
Hermann
de-a lungul unui axom mielinizat
In sistemul nervos central degenerescenta axonului sectionat are loc in sens bilateral. De exemplu, sectionand traiectul piramidal la nivel medular, degenerescenta se produce atat periferic (anterograd), cat si central (retrograd). Urmarirea acestei degenerescente constituie o metoda histologica utilizata pentru precizarea diferitelor conexiuni.
Potentialul de repaus si cel de actiune
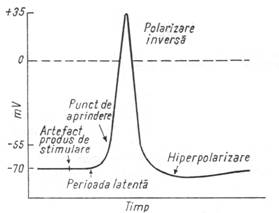
Excitatia si propagarea stimulului dealungul nervilor sunt
insotite de fenomene electrice. Plasand doi electrozi pe suprafata
unei fibre nervoase, in repaus, nu se remarca nici o deosebire de
potential. Daca insa, unul patrunde in citoplasma, se
constata ca interiorul axonului are un potential negativ de 70
mv. Acesta este generat in repaus de distributia inegala a ionilor de
Na si K, datorita pompelor active care polarizeaza membrana.
Prin stimulare, aceasta este depolarizata datorita unui influx de
ioni de Na. Potentialul de actiune dispare rapid, membrana fiind
repolarizata (fig. 4).
Figura 4 - Potentialul de actiune
Potentialul de repaus este restabilit initial prin efluxul ionilor de potasiu si influxul celor de clor. Cheltuiala de energie necesara proceselor de polarizare si depolarizare este furnizata de oxidarea tisulara din mitocondrii, prin metabolizarea unei importante cantitati de glucoza (tesutul nervos este cel mai sensibil la hipoglicemie).
Pentru a obtine o excitatie care se propaga, intensitatea stimulului trebuie sa aiba o valoare minima (pragul excitabilitatii), care variaza de la un tip de fibra la altul. Fibra nervoasa este excitata dupa legea tot sau nimic. Excitantii subliminari produc modificari, nemanifestate insa, coborand sau ridicand pragul excitabilitatii, in functie de efectul lor de depolarizare sau de hiperpolarizare. Daca depolarizarea atinge un anumit nivel critic (10-30 mV), ia nastere un potential de actiune care se propaga sub forma de excitatie.
Electrotonusul. Stimulii electrici subliminari modifica excitabilitatea in mod diferit la catod si la anod. La catod se produce o acumulare de sarcini pozitive, intracelulare care reduc polarizarea de repaus. Aceasta depolarizare creste progresiv cu intensitatea stimulului, pana cand diferenta de potential este redusa cu 15 mV, moment in care are loc procesul de activare cu aparitia potentialului de actiune. La anod, o redistributie ionica inversa creste valoarea potentialului de repaus prin hiperpolarizare. Intreruperea curentului poate declansa un curent de actiune. Aceste modificari de polarizare produse de stimuli sub prag poarta denumirea de electrotonus, cele de la catod fiind denumite catelectrotonus, iar cele de la anod, anelectrotonus.
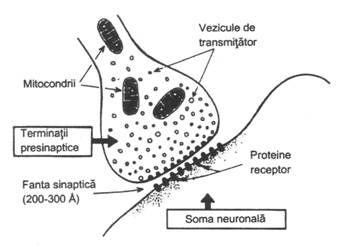
Impulsul nervos este transmis de la un neuron la altul cu ajutorul mediatorilor chimici, dintre care cel mai cunoscut este acetilcolina. Actiunea ei dureaza putin, datorita colinesterazei aflate la acest nivel si care o inactiveaza rapid. Efectul stimularii poate fi prelungit impiedicand actiunea acestei enzime cu ajutorul unor substante farmacodinamice numite anticolinesteraze (ex. ezerina). Alaturi de acetilcolina, ca mediator chimic sinaptic pot interveni si alte subtante ca: acidul glutamic, noradrenalina, serotonina, histamina.
Transmiterea chimica la nivelul sinapsei explica intarzierea de 0,7 ms pe care o prezinta, ca si marea fatigabilitate (oboseala), asa cum rezulta din epuizarea transmiterii daca excitatia are o frecventa de 40-50 impulsuri/s. Sinapsele sunt foarte sensibile la hipoxie prin afectarea ATP-ului, generator de energie pentru sinteza mediatorului. Impulsul sinaptic este descarcat repetitiv, ceea ce se explica prin eliberarea treptata a mediatorului. Mediatorul chimic este eliberat sub impulsul potentialului de actiune al axonului, care contine la capatul sinaptic vezicule de acetilcolina. Acetilcolina sau ceilalti mediatori chimici sunt eliberati in fanta sinaptica in cca. 0,6 ms de la stimulare, producand o depolarizare a somei neuronale postsinaptice (fig. 5). Cand depolarizarea atinge nivelul critic, impulsul este transmis spre periferie prin axonul postsinaptic.
Figura 5 - Sinapsa neuro-neuronala
Potentialul postsinaptic excitator. Daca stimulul presinaptic este slab, in soma motoneuronului se produce o depolarizare, care apare ca un potential de actiune redus, cu durata de cca. 20 ms. Acesta nu este urmat de o activare axonala, ramanand localizat. El este denumit potential postsinaptic excitator (PPSE) si creste proportional cu intensitatea stimulului. PPSE este produs printr-un influx ionic in zona neuronala, provocat de depolarizarea membranei sinaptice; este monofazic si proportional cu intensitatea impulsurilor.
Cand polarizarea somei neuronale este redusa la un nivel critic de 60 mV, apare o depolarizare brusca. Acest nivel critic provoaca un potential de actiune care se propaga dealungul axonului. Potentialul intracelular ajunge astfel, de la -70 mV la +20 mV, fiind deci de 90 mV fata de nivelul initial. Spre deosebire de PPSE, potentialul de actiune se supune legii tot sau nimic, nefiind neproportional cu intensitatea excitatiei.
Potentialul postsinaptic inhibitor. Anumiti stimului sinaptici in loc sa produca o depolarizare a somei neuronale, produc hiperpolarizare pana la -80 mV. Acest potential poarta denumirea de potential postsinaptic inhibitor (PPSI). El realizeaza o imagine in oglinda fata de PPSE.
Soma neuronala este astfel inhibata, stimularea necesitand o excitatie mai puternica. PPSI poate fi produs de exemplu prin stimularea unui nerv antagonist ipsilateral; este generat probabil de un transfer ionic prin porii somei neuronale, cu dimensiuni de cca. 5 suficiente pentru a permite trecerea pe K+ si Cl-, dar nu de Na+. K+ Paraseste astfel neuronul, concomitent cu influx de Cl-, care ii suplimenteaza sarcini negative.
Hiperpolarizarea este produsa de mediatori chimici inhibitori cu o compozitie putin cunoscuta. Se atribuie un rol important in acest sens acidului gammaaminobutiric (GABA). Unii cercetatorii sustin ca pe suprafata somei neuronale exista zone inhibitoare astfel incat inhibitia ar fi determinata de locul de aplicare a stimulului.
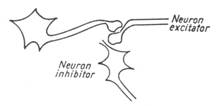
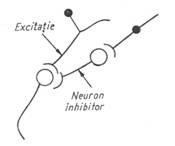
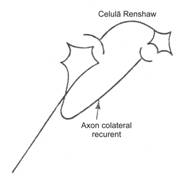
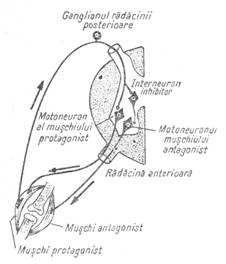
Inhibitia neuronala produsa prin hiperpolarizare sau prin alte mecanisme este tot atat de importanta ca si excitatia. Prin ea, raspunsul motor este modelat si coordonat, realizand o corespondenta intre activitatea muschilor antagonisti. S-au descris trei tipuri de inhibitie neuronala: directa, presinaptica si recurenta (fig. 6).
a) b)
c)
Figura 6 - a) Inhibitie directa; b) Inhibitie presinaptica; c) Inhibitie de tip Renshaw
Inhibitia directa consta din intercalarea unui neuron inhibitor care, prin mediator inhibitor, actioneaza asupra unui neuron stimulat convergent.
Inhibitia presinaptica diminua descarcarea mediatorilor la nivelul sinapsei (cantitatea fiind proportionala cu intensitatea potentialului de actiune), asa cum rezulta din scaderea PPSE. Ea este produsa de neuroni care actioneaza pe terminatiile celor excitatori sau pe axon.
Inhibitia recurenta este mediata de celulele Renshaw, care sunt intercalate intr-o ramificatie axonala recurenta si motoneuron. Stimulul se intoarce astfel la motoneuron, pe care il inhiba, producand o hiperpolarizare.
Numarul neuronilor care intra in alcatuirea organelor nervoase centrale este foarte mare. Impulsurile nervoase care apar la nivelul receptorilor sunt transmise neuronilor din centri nervosi. La nivelul sinapselor centrale impulsurile vor trece la neuronii motori, fie direct, fie indirect prin intermediul neuronilor intercalari. Centrii nervosi, existenti in diferite etaje nervoase, nu sunt izolati, ci intre ei exista numeroase legaturi. Datorita diferitelor procese de transmitere sinaptica, centrii nervosi confera impulsului un caracter diferentiat si nuantat. Transmiterea excitatiei prin centrii nervosi se caracterizeaza printr-o serie de particularitati.
Conducerea unilaterala a impulsului. Dupa cum s-a vazut, in fibra nervoasa impulsul este transmis in ambele sensuri. In centrii nervosi, transmiterea este insa unilaterala, fapt explicat prin mecanismul de eliberare a acetilcolinei la nivelul butonului terminal unde se depolarizeaza portiunea de membrana subiacenta.
Intarzierea sinaptica. Perioada latenta a unui reflex este mai mare decat cea justificata de timpul necesar pentru stimularea receptorului, conducerea prin caile aferente si eferente si raspunsul organului efector. Perioada latenta mai lunga este explicata prin intarzierea centrala sinaptica, adica prin timpul necesar ca un stimul sa fie transmis de la terminatia axonala la corpul neuronal. La nivelul fiecarei sinapsei impulsul intarzie 0,5-0,7 ms. Cu cat centrul nervos este alcatuit dintr-un numar mai mare de neuroni, cu atat este mai mare intarzierea centrala sinaptica. In consecinta, dupa durata perioadei latente se poate aprecia cu aproximatie si numarul neuronilor care formeaza centrul unui reflex.
b) a)
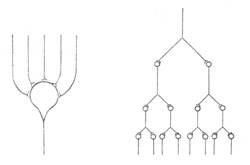
Convergenta si
divergenta. La suprafata fiecarui neuron
se termina un mare numar de prelungiri axonale. Fenomenul poarta
denumirea de convergenta. In acelasi timp axonul unui singur
neuron vine in contact prin terminatiile sale cu un mare numar de corpuri neuronale
printr-un fenomen denumit divergenta (fig. 7).
Figura 7 - a) Convergenta; b) Divergenta
Se presupune ca fiecare neuron primeste aferente axonale de la cca. 100 de neuroni, sau mai multi si trimite terminatii axonale la cca. 100 de neuroni. Posibilitatile de interconexiune neuronala sunt deci practic infinite.
Postdescarcarea. Prin postdescarcare se intelege faptul ca, numarul impulsurilor din nervii eferenti este mai mare decat al celor din fibrele aferente. In consecinta, prin trecerea prin centrii nervosi, stimulii sunt multiplicati. Acest proces poate fi explicat prin fenomenul divergentei, in sensul ca un stimul unic al nervului aferent este descarcat, prin numeroase ramificatii axonale, intr-un numar mai mare de neuroni motori eferenti. Multiplicarea stimulului la nivelul centrului nervos este importanta pentru realizarea contractiei tetanice, care este contractia fiziologica a muschiului striat.
Recrutarea. Prin recrutare se intelege cresterea numarului de unitati motoare stimulate in legatura cu cresterea intensitatii stimulului.
Inductia
simultana si succesiva. Centrii nervosi sunt sediul unor procese
de coordonare care sincronizeaza activitatea muschilor
antagonisti. Coordonarea este realizata prin inductie, care
poate fi simultana sau succesiva. De exemplu, concomitent cu
stimularea neuronilor care inerveaza extensorii unui membru se produce
relaxarea flexorilor, care sunt antagonisti. Fenomenul se numeste
inductie negativa simultana. Acesta este un proces de coordonare
locala, probabil prin inhibitie neuronala directa. Astfel,
de exemplu, in decursul reflexului osteotendinos, impulsurile aferente sunt
transmise neuronilor motori ai extensorilor, in timp ce este stimulat si
un neuron inhibitor intercalar care inhiba motoneuronii flexorilor (fig.
8).
Figura 8 - Conexiuni centrale probabile in coordonarea
muschilor antagonisti
Dupa ce motoneuronii extensorilor au fost stimulati ei sunt inhibati probabil prin hiperpolarizare. Fenomenul se numeste inductie negativa succesiva. In acelasi timp, flexorii, anterior inhibati, sunt stimulati, tot prin inductie succesiva, dar pozitiva. Prin aceasta interferenta dintre excitatia si inhibitia din centrii nervosi sunt coordonate extensia si flexia unui membru. Procese de inductie succesiva survin si la nivelul altor centri nervosi care aferenteaza musculatura antagonista, ca de exemplu in centrii nervosi din trunchiul cerebral, care comanda miscari de masticatie, coordonand coborarea si ridicarea mandibulei.
Iradierea. Iradierea este fenomenul prin care cresterea intensitatii stimulului activeaza o zona progresiv, mai mare de neuroni centrali.
Sistemul nervos indeplineste rolul de reglare, coordonare si integrare a functiilor tuturor organelor. El asigura in acelasi timp unitatea organismului cu mediul inconjurator, realizand unitatea organism-mediu si coordoneaza activitatea organelor interne.
Sistemul nervos central este
in permanenta informat prin stimulii receptionati din afara
si din interiorul organismului, ceea ce face posibila integrarea
organismului in mediul inconjurator si mentinerea
- compartimentul senzitiv, unde sosesc informatiile culese de la nivelul receptorilor;
- compartimentul motor, care transmite comenzile la efectori.
Deci fiecare organ nervos are doua functii fundamentale: functia senzitiva si functia motorie. La nivelul emisferelor cerebrale mai apare si functia psihica. Separarea functiilor sistemului nervos este doar cu caracter didactic; in relitate nu exista activitate senzitiva fara manifestari motorii si viceversa, iar starile psihice deriva din integrarea primelor doua.
Functiile fundamentale ale sistemului nervos sunt functia reflexa si functia de conducere.
Functia reflexa.
Reflexul reprezinta mecanismul fundamental de activitate a sistemului nervos. El reprezinta reactia de raspuns a centrilor nervosi la stimularea unei zone receptoare. Termenul a fost introdus in urma cu 300 de ani de catre matematicianul si filosoful francez Rene Descartes. Raspunsul reflex poate fi excitator sau inhibitor.
La realizarea unui reflex participa cinci componente anatomice: receptorul, calea aferenta, centri, calea eferenta si efectorul. Ele alcatuiesc impreuna arcul reflex.
Receptorul este de obicei o celula sau un grup de celule diferentiate si specializate in celule senzoriale (gustative, auditive, vizuale, vestibulare). Alti receptori sunt corpusculii senzitivi alcatuiti din celule, fibre conjunctive si formatiuni nervoase dendritice (receptorii tegumentari si proprioceptorii). Uneori rolul de receptor il indeplinesc chiar terminatiile butonate ale dendritelor (receptorul olfativ, receptorii durerosi).
La nivelul receptorului are loc transformarea energiei excitantului in influx nervos. Fiecare receptor este specializat in transformarea unei anumite forme de energie din mediu (excitanti sau stimuli) in informatie nervoasa specifica (influxul nervos sau potentialul de actiune) cu amplitudine proportionala cu intensitatea excitantului. In acelasi timp, fiecare receptor poate fi stimulat de orice forma de energie daca depaseste cu mult intensitatea normala; astfel celulele vizuale pot fi excitate si de energii mecanice mari (o lovitura cu pumnul in ochi provoaca senzatii vizuale rudimentare).
Receptorii se pot clasifica dupa mai multe criterii.
Dupa localizare:
exteroceptori, situati la nivelul pielii si organelor de simt, care culeg din mediul extern stimuli termici, durerosi, tactili, olfactivi, vizuali, auditivi;
- proprioceptorii, situati la nivelul muschilor, tendoanelor, articulatiilor, periostului si ligamentelor;
- interoceptori (visceroceptori) care culeg stimulii de la nivelul arborelui vascular si al organelor interne (viscere).
Dupa specializare:
- mecanoreceptori, sensibili la excitantii mecanici;
- chemoreceptori, sensibili la actiunea substantelor chimice;
- osmoreceptori, sensibili la variatiile presiunii osmotice;
- termoreceptori, sensibili la variatiile de temperatura;
- voloreceptori, sensibili la variatiile de volum;
- fotoreceptori, sensibili la energiile luminoase;
- algoreceptori, sensibili la stimulii nocivi, durerosi.
Calea aferenta. Receptorii vin in contact sinaptic cu terminatiile dendritice ale neuronilor senzitivi din ganglionii spinali sau de pe traiectul unor nervi cranieni. Variatiile de potential de receptor produc depolarizari pasive in terminatia dendritica care atingand pragul critic descarca potential de actiune dupa legea 'tot sau nimic' ce se propaga celulipet (aferent). Informarea corecta= a centrilor privind variatiile energiei excitantului se face prin modulare de frecventa (modularea in amplitudine nu este posibila din cauza legii tot sau nimic). Ca urmare un potential de receptor de amplitudine redusa determina numai cateva potentiale de actiune pe secunda, in timp ce potentiale de receptor mai ample induc zeci sau sute de potentiale de actiune pe secunda. Cea mai simpla cale aferenta este reprezentata de neuronul senzitiv spinal si prelungirile sale.
Centrul. In cazul unui arc reflex elementar format din doi neuroni (unul senzitiv, celalalt motor) centrul reflex este reprezentat chiar de sinapsa dintre axonul neuronului senzitiv si corpul neuronului motor (exemplu reflexul miotatic). Prin centrul unui reflex se intelege totalitatea structurilor din sistemul nervos central care participa la actul reflex respectiv. Spre exemplu, centrii reflexelor respiratori se afla in bulb, in punte precum si in hipotalamus si in scoarta cerebrala.
Complexitatea si intinderea unui centru este in functie de complexitatea actului reflex pe care il efectueaza. Centrul nervos care primeste informatia o prelucreaza, o compara cu date din memorie, elaboreaza decizii si controleaza permanent modul de executie efectuand corectarile necesare. La nivelul sinapselor din centrii reflecsi, transmiterea se face din nou prin modulare in amplitudine deoarece potentialele postsinaptice nu mai respecta legea 'tot sau nimic'.
Calea eferenta este reprezentata de axonii neuronilor efectori (motori sau secretori). Cea mai simpla cale eferenta se gaseste la reflexele monosinaptice cand este formata din axonul motoneuronului . In cazul sitemului nervos vegetativ, calea efrenta este formata dintr-un lant de doi neuroni motori: un neuron preganglionar situat in coarnele laterale ale maduvei spinarii sau intr-un nucleu vegetativ din trunchiul cerebral si un neuron postganglionar situat in ganglionii vegetativi periferici. De-a lungul cailor eferente informatia circula spre efectori din nou prin modulatie de frecventa.
Efectorii. Principalii efectori sunt muschiul striat, muschiul neted si glandele exocrine. In functie de fracventa potentialelor de actiune sosite pe axon, la nivelul placii motorii de exemplu, se vor sparge un anumit numar de vezicule cu acetilcolina, care va determina potentiale postsinaptice de amplitudini diferite in functie de numarul de molecule de acetilcolina eliberate. Ca urmare, pe membrana fibrei striate apar zeci sau sute de potentiale de actiune pe secunda, producand contractii de amplitudine si forte corespunzatoare comenzii centrale.
Controlul indeplinirii comenzii. Arcul reflex cu cele cinci componente al sale reprezinta un model incomplet al desfasurarii activitatii reflexe. In ultimele decenii s-a descoperit existenta unor noi circuite nervoase ce leaga centrii de organele receptoare. Prin intermediul acestora, centrii nervosi pot regla pragul de excitabilitate al receptorilor si implicit intensitatea stimulilor aferenti. Un asemenea control se exercita asupra efectorilor musculari de catre centrii motori extrapiramidali si cerebrali.
In acelasi timp de la nivelul efectorilor porneste spre centrii un circuit recurent care ii informeaza asupra modului indeplinirii comenzii (feed-back). Mecanismul de feed-back are caracter universal ce se intalneste in toate sistemele autoreglate. Comanda si controlul exercitate de centrii nervosi sunt de natura reflexa. In acest sens, centrii nervosi nu sunt numai senzitivi sau numai motori, ci ei reprezinta centrii de integrare senzitivo-motorie. Raspunsul reflex poate surveni imediat dupa actiunea stimulului sau poate intarzia minute, zile sau ani; in aceste cazuri informatia ce intra in centrii este depozitata sub forma de memorie si va fi actualizata in momentul elaborarii raspunsului.
In raport cu efectorul asupra caruia se exercita preponderent comanda, sistemul nervos poate fi impartit in:
- sistem nervos al vietii de relatie (somatic) care controleaza efectorul muscular striat voluntar;
- sistem nervos al vietii vegetative (de nutritie) care controleazan efectorul muscular neted si glandular, coordonand activitatea organelor interne (viscere). Este format din sistem nervos simpatic (care are ca mediator chimic noradrenalina si adrenalina) si sistem nervos parasimpatic (a carei activitate este mediata chimic de acetilcolina).
Activitatea sistemului nervos somatic si a celui vegetativ nu se desfasoara separat ci intr-o unitate perfecta; hipotalamusul, sistemul limbic si scoarta cerebrala integreaza si coordoneaza activitatea viscerelor, glandelor endocrine si cea somatica. In felul acesta se realizeaza atat unitatea organismului in ansamblu cat si unitatea organism-mediu.
Functia de conducere.
Excitatiile culese la nivelul suprafetelor receptoare sunt conduse spre centrii, iar comenzile elaborate aici sunt conduse la periferie la organele efectoare. Intreg sistemul nervos apare ca un ansamblu imens de circuite functionale, bidirectionale prin care informatiile circula necontenit.
Viteza de conducere a influxului nervos depinde de structura si grosimea fibrelor nervoase. Fibrele cu diametru mare conduc mai repede decat cele inguste, iar pentru aceeasi grosime fibrele mielinice conduc cu viteze mai mari decat cele amielinice. Viteza cea mai mare o au fibrele cailor proprioceptive (120 m/s) iar viteza cea mai redusa o au fibrele amielinice ca transmit stimulii durerosi (0,5 m/s). Conducerea excitatiei de la periferie spre centru se numeste conducere afenta, iar a comenzilor de la centru la periferie se numeste conducere eferenta.
Functia de conducere si functia reflexa sunt strans corelate, separarea lor se face numai in scopuri didactice.
Atat sistemul nervos somatic cat si cel vegetativ au o componenta centrala (centrii nervosi) situata in sistemul nervos central (maduva spinarii si encefalul) si o componenta periferica (ganglioni si trunchiuri nervoase), ce constituie sistemul nervos periferic.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5013
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved