| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
PROPRIETATILE MIOCARDULUI
1. Automatismul si ritmicitatea-functia cronotropa;
2. Excitabilitatea-functia batmotropa;
3. Conductibilitatea-functia dromotropa;
4. Contractilitatea-functia inotropa;
5. Tonicitatea-functie tonotropa.
1.Automatismul si Ritmicitatea - Functia cronotropa
Reprezinta proprietatea miocardului de a genera spontan si ritmic stimuli care ii produc contractia.
Geneza stimulilor - 2 teorii:-Teoria miogena si Teoria neurogena.
Teoria miogena, argument: la organismele superioare sunt furnizate argumente pentru teoria miogena conform careia elementele musculare, miocardice sunt cele care genereaza stimuli:
- inima puiului de gaina incepe sa bata la scurt timp dupa incubare, inainte de aparitia elementelor nervoase;
- in culturile de celule se evidentiaza autodepolarizarea celulelor,,P'';
- inima de broasca izolata (scoasa din torace), fara legaturi nervoase continuua sa bata.
Suportul morfologic al automatismului cardiac este tesutul excitoconductor, format din noduli de automatism si structuri de legatura intre acestia (cai de conducere specifice).
In structura nodulilor de automatism se gasesc celulele pacemaker (P), cu particularitati structurale si functionale care permit generarea unui potential de actiune (PA):
in membrana lor sunt putine canale rapide de Na+, in schimb sunt bine reprezentate canalele lente de Na+ si Ca 2+;
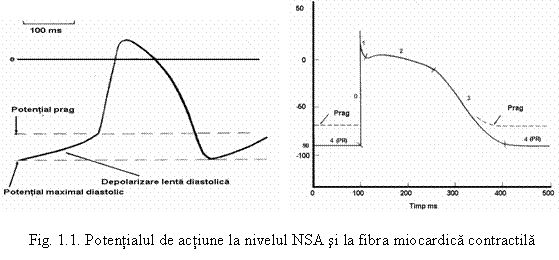
au un potential de repaus - PR- inconstant care se numeste potential diastolic maximal - PDM;
in momentul atingerii PDM se deschid canalele lente de Ca 2+ si Na+ , acesti ioni incep sa intre in celula determinand o depolarizare lenta diastolica (numita si prepotential), care atunci cand atinge un anumit prag = potentialul prag declanseaza potentialul de actiune si celula se autodepolarizeaza
la celulele nodulului sino-atrial - NSA, potentialul de repaus este inconstant, panta de depolarizare (0) este lenta, potentialul de actiune are amplitudine mai mica decat a altor celule P sau cel de la fibra miocardica adulta, nu are spike, varful fiind rotunjit, nu prezinta faza de platou caracteristica fibrei miocardice de tip adult, durata PA este mai mica, ~120 ms decat la fibra miocardica adulta, unde are o valoare de 260-280 ms. Durata PA creste treptat de la NSA, la nodulul atrio-ventricular - NAV ~ 150 ms, la reteaua Purkinje ~ 300 - 310 ms (acesta durata mare explica de ce repolarizarea la miocardul ventricular se fac in sens invers depolarizarii, dinspre epicard spre endocard).
Frecventa de generare a stimulilor la nivelul nodulilor de automatism depinde de:
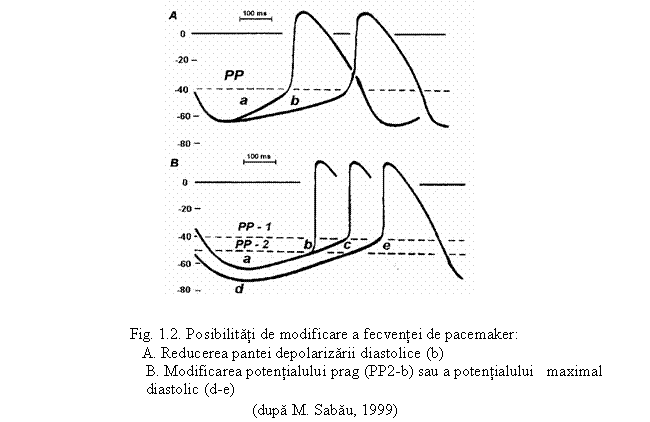
● Valoarea potentialului diastolic maximal, sau potentialul de repaus Cu cat valoarea PDM va fi mai indepartata de cea a potentialului prag cu atat durata pantei prepotentialului va fi mai mare, inregistrand un efect cronotrop negativ - pe unitatea de timp celula se va depolariza de un numar mai mic de ori, deci frecventa va fi mai mica.
Cand PDM este apropiat de valoarea potentialului de repaus, durata pantei autodepolarizarii fiind mai mica, frecventa creste - efect cronotrop pozitiv.
● Valoarea potentialului prag - cu cat valoarea PP se apropie de PDM se va inregistra un efect cronotrop pozitiv; cand valoarea PP se indeparteaza de PDM -efect cronotrop negativ.
● Rapiditatea, bruschetea pantei de autodepolarizare lenta diastolica (prepotentialul)
La NSA viteza prepotentialului este estimata la 0,1V/s. Ea scade la Reteaua Purkinje, ajungamd la 0,03 V/s. Astfel se poate explica de ce NSA descarca stimuli cu frecventa intre 60-100/minut atunci cand este influenta vagala, iar scos de sub influenta vagului ν=120-150 st/min.
NAV, cu o panta a prepotentialului mai lenta descarca stimuli cu 40-60/min, iar focarul tertiar de automatism (facsiculul His+Reteaua Purkinje cu o ν=25-35 st/min.
Elementul care ierarhizeaza centrii de automatism e reprezentat de frecventa de descarcare a stimulilor, dependenta de caracteristicile pantei prepotentialului.
Pacemakerii se pot clasifica in:
Pacemakeri naturali-centrele de automatism;
Pacemakerii ocazionali-focarele ectopice;
Pacemakerii artificiali-stimularea artificiala a cordului se poate face transtoracic sau endocavitar.
Stimularea transtoracica (de scurta durata) se face cu 2 electrozi cu suprafata mare plasati pe toracice- unul in zona socului apexian si cel de-l 2-lea, simetric pe hemitoracele drept.
Stimularea endocavitara - pe o durata lunga, se realizeaza cu ajutorul unei sonde electrod introdusa pe cale venoasa, pozitionata la nivelul AD sau VD, pusa in legatura cu un generatoar de impulsuri.
Pacemakeri artificiali pot fi: asincroni - lucreaza tot timpul la aceiasi parametrii. Prezinta doua mari dezavantaje- nu permit adaptarea activitatii cardiace la necesitatile variabile, de exemplu la efort fizic, iar activitatea lor poate interfera cu activitatea spontana a cordului care se poate relua ulterior.
- Pacemakeri la cerere sau a demand - lucreaza la aceiasi parametri tot timpul, dar intra in actiune doar la oprirea activitatii cardiace pe o perioada de 800-1000ms.
- Pacemakeri stand-by, cu sincronizare ventriculara sau atriala, care permit adaptarea activitatii cardiace la diverse necesitati. Sunt folositi mai ales in blocurile atrio-ventriculare complete.
Cum recunoastem prezenta unor pacemakeri pe EKG la un pacient?
- Pe traseu apare artefactul de stimulare, inaintea complexului QRS
- Daca pacemakerul e plasat la nivelul VD, aspectul QRS este asemanator cu cel intalnit in blocul de ramura stanga.
Factorii care influenteaza functia cronotropa:
- SN Vegetativ parasimpatic exercita un efect cronotrop negativ deoarece prin acetilcolina mareste permeabilitatea membranara pentru ionul de K+, care va iesi din celula si va determina o hiperpolarizare a membranei, prelungind durata pantei prepotentialului.
- SNV simpatic exercita un efect cronotrop pozitiv. Epinefrina mareste permeabilitatea membranara pentru Ca2+ si Na+ care vor intra mai rapid in celula micsorand durata pantei prepotentialului si accelerand viteza de depolarizare.
- Tempereatura - cresterea determina un efect cronotrop pozitiv usor observabil in starile febrile cand se instaleaza tahicardia.
- Modificarea concentratiei plasmatice a unor ioni:
- Cresterea concentratiei K+ extracelular peste 6,5 mEq/1 are efecte deprimante asupra functiei cronotrope; la valori ale potasemiei peste 8,5 mEq/1, cordul se opreste in diastola - inhibitio potassica;
- Hipokalemia moderata are efect cronotrop pozitiv
- Factorii mecanici-distensia fibrelor poate crea conditii ca pacemakerii ocazionali sa se manifeste
Ritmurile reprezinta succesiunea regulata a ciclurilor cardiace. Ritmul normal al inimii este ritmul imprimat de NSA (pricipalul focar de automatism) si se numeste ritm sinusal
Ritmul sinusal - caracteristici ECG:
sa existe unda P, care trebuie sa preceada QRS si sa fie pozitiva in cel putin doua dintre derivatiile standard
distantele P - P = R-R si egale de la un ciclu la altul
▪ intervalul Pq = 0,12 - 0,21 sec.;
▪ frecventa cardiaca: 60 - 100/min;
Daca frecventa este sub 60 batai/min, dar sunt pastrate celelate caracteristici, vorbim de bradicardie sinusala, iar daca frecventa depaseste 100 batai /min de tahicardie sinusala.
Ritmuri patologice
Ritmul nodal, generat in nodulul atrio-ventricular. Este de 3 feluri, in functie de locul de emitere a stimulilor: supranodal, mezo(medio)nodal si infranodal;
- F = 40-60/min.
- unda P, cand apare,este negativa deoarece activarea ei se face retrograd;
- in cazul ritmului supranodal, unda p este inaintea QRS si este negativa, iar intervalul pq va avea durata sub limita inferioara fiziologica.
- unda p absenta - ritm medionodal.
- in cazul ritmului infranodal, unda p este dupa QRS si este negativa.
Ritmul idioventricular, generat de fasciculul His si reteaua Purkinje
- frecventa 25-35/min;
- lipsa undelor P in fata complexelor PQR;
- complexe QRS largite, deformate datorita conducerii stimulilor pe cai nespecifice.
In conditiile unui blocaj atrio-ventricular complet avem unde P care nu au legatura cu complexele ORS. (atriile se contracta dupa alt ritm fata de ritmul V).
2. Excitabilitatea - functia batmotropa
Este proprietatea anumitor tesuturi de a raspunde specific la stimuli.
Reprezinta o treapta superioara a iritabilitatii - proprietate a intregii materii vii de a raspunde la stimuli.
Este caracteristica sistemului nervos, muscular si endocrin.
La nivelul miocardului, excitabilitatea are urmatoarele caracteristici:
respecta legea ,,tot sau nimic'': la un stimul cu intensitatea cel putin liminara, miocardul raspunde cu amplitudine maxima. Oricat creste intensitatea stimulului, amplitudinea raspunsului ramane aceeasi. La un stimul cu intensitatea subliminara miocardul nu raspunde. Miocardul se supune acestei legi spre deoarece d.p.d.v functional se comporta ca un sincitiu. La muschiul scheletic aceasta lege nu este respectata, pragul de excitatie a diferitelor unitati motorii fiind diferit. Pe masura ce creste intensitatea stimulului, la muschiul scheletic se recruteaza in raspunsul contractil noi unitati motorii.
OBS Fenomenul in trepte al lui Bowditeh
Amplitudinea raspunsului miocardic e dependenta de conditiile metabolice. La reluarea activitatii sale dupa o perioada de oprire, amplitudinea poate fi mai redusa, dar ea reprezinta tot amplitudinea maxima pe care o poate da miocardul in acele conditii metabolice. Reluandu-si activitatea si irigarea cordului imbunatatindu-se, amplitudionea creste si in acest caz, reprezinta tot maximul, dar in conditii metabolice diferite, mai favorabile contractiei.
Miocardul respecta legea Marey - legea ,,inexcitabilitatii periodice''- cordul este inexcitabil in sistola; la sfarsitul sistolei sau inceputul diastolei poate raspunde la stimuli cu intensitati mari, dupa o perioada de latenta crescuta; progresand in diastola, revine la normoexcitabilitate.
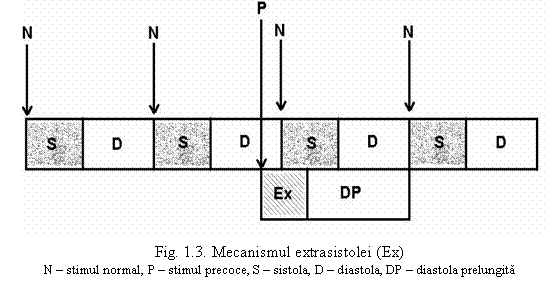
In timpul depolarizarii si primei faze a repolarizarii, miocardul se afla in perioada refractara absoluta; un stimul aplicat acum nu este urmat de raspuns, indiferent de intensitatea lui. Spre finalul repolarizarii si mai ales in perioada postpotentialelor se afla in faza refractara relativa (faza vulnerabila) si poate raspunde la stimuli cu intensitate mare. Un stimul venit dintr-un focar ectopic, determina in acesta faza aparitia unei contractii precoce = extrasistola. Ca orice contractie (sau sistola) si extrasistola are o perioada refractara absoluta sau inexcitabila. Din acest motiv, urmatorul stimul fiziologic, venit din focarul de automatism va gasi cordul in faza refractara absoluta a extrasistolei, deci in perioada inexcitabila si nu va fi urmat de un nou raspuns. Urmeaza deci o pauza sau o diastola prelungita
Urmatorul stimul normal, fiziologic (pornit din pacemaker-ul cardiac) gaseste inima in aceasta pauza dupa extrasistola (diastola prelungita) deci in faza excitabila si astfel poate raspunde la excitant. In patologia umana si chiar la indivizii normali, se intalnesc frecvent extrasistole ca manifestare clinica ce traduce o perturbare a excitabilitatii miocardice, cu aparitia de focare ectopice generatoare de extrasistole.
▪ Respecta legea conservarii perioadelor fiziologice a lui Engelmann: un stimul cu intensitatea cel putin liminara, aplicat in afara perioadei refractare va determina un raspuns prematur reprezentand extrasistola. Dupa extrasistola poate sa apara o diastola prelungita datorata lipsei de raspuns a miocardului la stimulul care vine de la focarul fiziologic de automatism, lipsa de raspuns datorata perioadei refractare a extrasistolei. Prin urmare, chiar si in cazul aparitiei unei extrasistole, perioadele fiziologice ale activitatii cardiace, sistola si diastola se conserva.
▪ Contractia miocardului este o secusa - miocardul nu se tetanizeaza. Secusa reprezinta contractia produsa de un singur stimul. Secusei i se pot delimita perioadele de: latenta, contractie si relaxare.
Insumarea secuselor=TETANOSUL.
Aparitia platoului la nivelul PA mareste durata acestuia, aducand-o in apropierea duratei fenomenului contractil. Prin urmare la miocard, durata fenomenului electric, adica a PA este foarte apropiata de a fenomenului mecanic, adica contractia, spre deosebire de muschiul scheletic, la care PA dureaza doar 2-4 ms, iar contractia, in medie zeci de ms pana la sute de ms.
Marirea perioadei refractare face ca miocardul sa poata raspunde la un alt stimul abia dupa terminarea (aproape) a fenomenului contractil. De aceea, contractile nu se pot insuma.
Spre deosebire de miocard, la muschiul scheletic, raspunsurile contractile sunt in majoritate de tip tetanos, insumarea contractiilor este posibila datorita diferentei mari intre durata fenomenului electric si a celui contractil. De aceea muschiul este capabil sa raspunda la mai multi stimuli electrici, raspunsurile mecanice corespunzatoare lor insumandu-se.
Excitabilitatea depinde de:
- Nivelul (valoarea) potentialului de repaus si a potentialului prag. Cu cat cele doua valori sunt mai aproapiate cu atat excitabilitatea structurii respective va fi mai mare. Este valabila si situatia inversa.
Factorii care influenteaza excitabilitatea
▪ SN vegetativ parasimpatic are efect batmotrop negativ deoarece mareste permeabilitatea membranei pentru K, produce astfel hiperpolarizare care este urmata de hipoexcitabilitate;
▪ SN vegetativ simpatic prin marirea permeabilitatii membranei pentru Na si Ca determina o hipopolarizare a membranei urmata de hiperexcitabilitate - efect batmotrop pozitiv;
▪ Temperatura crescuta - determina hiperexcitabilitate;
▪ Factori mecanici - distensia fibrelor-efect batmotrop pozitiv;
▪ Factori metabolici- hipoxia modificand pragul de excitabilitate conduce la hiperexcitabilitate;
- dereglari endocrine-hipertiroidismul determina hiperexcitabilitate;
- hiperfunctia medulosuprarenalei prin cresterea concentratiei catcolaminelor induce hiperexcitabilitate;
- Hipertiroidismul creste excitabilitatea cardiaca, pana la instalarea fibrilatiei atriale.
pH-ul acid are de asemenea un efect de crestere a excitabilitatii miocardului
Tulburari de excitabilitate: aritmii
A. Tranzitorii - de scurta durata
- Extrasistole/Tahicardii paroxistice. Extrasistolele pot aparea datorita unor focare ectopice din musculatura atriala, NAV, miocardul ventricular.
Pot fi: - monomorfe (acelasi aspect)=unifocare;
- polimorfe-plurifocale;
- nesistematizate cand nu exista o relatie fixa intre extrasistola si contractiile ritmului de baza; sau sistematizate - exista o relatie fixa.
- Tahicardiile paroxistice sunt fenomene care apar pe durata variabila, in timpul lor frecventa ajunge la 160-200 contractii/min. Ele pot fi supraventriculare sau ventriculare.
B. De lunga durata - Flutterul si Fibrilatia atriala
- Fibrilatia ventriculara - tulburare foarte grava, in care eficienta hemodinamica a ventriculilor este compromisa si daca nu se intervine cu defibrilare, pacientul moare.
3. CONDUCTIBILITATEA - FUNCTIA DROMOTROPA
Reprezinta proprietatea miocardului de a prelua stimuli de la focarul de automatism si de a-i conduce in toata masa sa. Conducerea se face:
- specific prin componentele sistemului excito-conductor, cu viteza de conducere mare de 1-4 m/sec.
- nespecific, conducere ephaptica, se realizeaza din aproape in aproape, prin fibrele miocardului de aceea viteaza este mica 0,1-0,3 m/s.
Conductibilitatea depinde de mai multi factori:
▪ factorul geometric - raportul care se stabileste intre portiunea membranara depolarizata si cea inca in repaus dintr-un anumit teritoriu.Atunci cand valoarea raportului este mai mare si conducerea stimulilor se realizeaza mai usor si reciproca este si ea valabila.
▪ numarul si calitatea discurilor intercalare;
▪ parametrii potentialului de actiune - durata PA la nivelul miocardului adult este de 260-280 ms iar la nivelul retelei Purkinje creste la 310 ms. Avand perioada refractara mare, celulele retelei Purkinje se comporta ca un baraj, stopeaza trecerea stimulilor emisi de focarele ectopice spre miocardul propriu-zis. Ele se mai numesc si ,,celule poarta''.
▪ pragul de excitabilitate -atunci cand pragul de excitabilitate este scazut (are o excitabilitate mai mare) conducerea stimulului se realizeaza mai usor.
La nivel atrial conducerea stimulilor se face atat pe cai specifice cat si nespecifice. Conducerea specifica se face prin tracturile internodale, cu viteza de 0,7-1,2 m/s, iar cea nespecifica prin musculatura atriala, din aproape in aproape, cu viteze mult mai mici (cel mult 0,2 m/s. )
La nivelul NAV, datorita structurii neomogene a acestuia, conducerea stimulilor are o intarziere de pana la 0,1 s. Acesta intarziere este data de prezenta la acest nivel a elementelor musculare atriale si a fibrelor de jonctiune care au diametrul redus, opunand o rezistenta mai mare la trecerea stimulilor
- Zona superioara, atrio-nodala in afara celulelor nodale sunt prezente elemente musculare atriale si fibre de jonctiune care au diametrul redus, opunand o rezistenta mai mare la trecerea stimulilor; de aceea la nivelul acestei zone stimulul intarzie cu pana la 0,04 s.
- In zona nodala, fiind prezente mai multe cellule nodale,viteza de conducere a stimulilor este mai mare intarzierea fiind de pana la 0,02 s.
- In zona nodo-hisiana- pe langa celule nodale sunt prezente din nou fibre de jonctiune si elemente de origine ale fasciculului His; intarzierea ajunge pana la 0,04 s.
La nivel VENTRICULAR, conducerea specifica se face prin faciculul His, care dupa o scurta portiune iniatiala da doua ramuri: ramura dreapata mai lunga care urmareste subendorcardic septul interventricular, ajunge la apex si se ramifica formand reteaua Purkinje si ramura stanga care are un traiect mai scurt, da o ramura anterioara si una posterioara. Acestea, la nivel suebdocardic vor genera reteaua Purkinje. La nivelul fasciculului His, viteza de conducere este de 2-3 m/s, iar in reteua Purkinje de 2-3-4 m/s.
Conducerea nespecifica se face prin fibrele miocardice ventriculare, din apropae in aproape,cu viteze intre 0,3-0,7 m/s.
Factorii care influenteaza dromotropismul:
▪ SNV parasimpatic- prin nervul efector cardiac - vag si mediatorul acestuia care este acetilcolina - are un efect dromotrop negativ. Cei 2 nervi vagi au distributie diferita:
- Vagul drept se distribuie predominant la nivelul NSA. Prin stimularea lui se obtin in special efecte cronotrop negative (scade frecventa cardiaca)
- Vagul stang se distribuie predominant la NAV iar stimularea lui conduce in special la efecte dromotrop negative (incetinirea conducerii)
Prin acetilcolina care actioneaza pe receptorii M2, parasimpaticul produce cresterea permeabilitatii membranare pentru ionul de K ceea ce duce la o hiperpolarizare membranara. Conducerea stimulului in acest caz e mai dificila.
▪ SNV simpatic - nervii cardiaci determina efecte drmotrop pozitive prin mediatorii sai: adrenalina(la nivelul suprarenalei mai multa adrenalina) si noradrenalina 85-89%, care prin marirea permeabilitatii membranare pentru Na+ produc o depolarizare de fond a membranei (in acest caz membrana este hipopolarizata, deci hiperexcitabila) ceea ce faciliteaza conducerea stimulilor.
▪ Factorii mecanici - distensia fibrelor - densitatea sarcinilor scade si mimeaza o hipopolarizare, deci efect dromotrop negativ .
▪ Factorii metabolici- hormonii tiroidieni influenteaza polarizarea membranara si implicit conducerea stimulilor. In hipertiroidism, conducerea este accelerata.
▪ pH scazut si cresterea temperaturii
Tulburarile dromotropismului: sunt determinate fie de incetinirea conducerii, fie de accelerarea ei
1. Tulburari de conducere caracterizate prin incetinirea si chiar sistarea conducerii stimulului= blocuri
▪ BLOCUL SINOATRIAL - se instaleaza o perioada de,,liniste electrica"(asemanatoare cu ligatura I Stannius)
▪ BLOCURILE ATRIOVENTRICULARE sunt de trei grade:
- de gradul I prin alungirea constanta a segmentului PQ (vezi corelarea cu frecventa cardiaca);
- de gradul II - cu 3 variante vezi LP;
- de gradul III - blocul atrioventricular complet. Intre unda P si complexul qRS nu e o relatie constanta. Atriile se contracta in ritm sinusal, iar ventriculii in ritm idioventricular. Blocul atrioventricular complet poate aparea si daca sunt blocate toate ramurile fasciculului His (deci nu trebuie sa fie neaparat la nivelul NAV).
Aparitia blocului atrioventricular complet impune implantarea de pace-makeri cardiaci artificiali, deoarece frecventa de 28-35 st/min nu e suficienta. Este asemanator ligaturii III a lui Stannius.
▪ BLOCURILE DE RAMURA
- blocul de ramura dreapta - stimulii vor parcurge mai intai ramura stanga producand depolarizarea pe cai specifice a ventriculului stang si apoi din aproape in aproape se va depolariza si ventriculul drept. Depolarizarea ventriculara va dura mai mult, deci va creste durata complexului ORS; undele date de ventriculul drept vor fi largite, deformate - aspect rr'in V1 V2, iar vectorii nu vor mai avea orientare normala. Axul ORS va fi deviat la dreapta.
- blocul de ramura stanga - similar, de data aceasta modificarile vor fi la nivelul undelor ventriculului stang - aspect RR' in V5V6, iar axul electric va fi deviat la stanga.
Se mai pot inregistra leziuni la nivelul ramificatiei ramurei stangi a Fasciculului His si astfel sa avem HEMIBLOC ANTERIOR=BLOC FASCICULAR ANTERIOR prin lezarea ramurei anterioare a ramurei stangi si HEMIBLOC POSTERIOR=BLOC FASCICULAR POSTERIOR prin lezarea ramurei posterioare.
2. Conducerile accelerate al caror suport este reprezentat de fascicule aberante de conducere - Palladino-Kent, James si Mahaim.
▪ Prezenta fasciculului Palladino-Kent determina aparitia sindromului Wolff - Parkinson-White - WPW caracterizat prin scurtarea intervalului PQ pe seama segmentului PQ si aparitia undelor delta ( ) la nivelul complexului QRS, motiv pentru care complexele sunt deformate si au o durata mai mare;
▪ Fasciculul James duce la aparitia sindromului Lown-Ganong-Levine - LGL caracterizat de scurtarea segmentului PQ, complexele QRS fiind normale;
▪ Fasciculul Mahaim apare unda delta, segmentul PQ este normal .
4. CONTRACTILITATEA functia inotropa
Este proprietatea tesututlui muscular de a raspunde specific la stimulii veniti din focarele de automatism printr-o contractie. La miocard cresterea tensiunii in interiorul fibrelor musculare este urmata intotdeauna de scurtarea fibrelor
Muschiul striat cardiac are asemanari morfologice, functionale atat cu muschiul neted, cat si cu muschiul striat scheletic:
- asemanarai cu muschiul neted:
- nu are unitati motori
- PA, prin durata si amplitudine este asemanatoar;
- sarcolema contine canale de Ca2+ voltage-gated
- are posibilitatea sa emita stimuli care sa-i genereze contractia;
- activitatea este sub influenta SNV.
- asemanari cu muschiul scheletic:
- forta de contractie asemanatoare cu cea dezvoltata de muschiul scheletic;
- viteza contractiei;
- unele carcateristici structurale (ex:numarul mare de mitocondrii)
Miocardul este alcatuit din fibre cu diametrul de 10-20μ si lungimi de 50-120μ. Acestea sunt formate din: sarcolema, sarcoplasma si nucleu cu 1-2 nucleoli.
In sarcoplasma, se gasesc organite specifice - miofibrile in numar de 300-600 de miofibrile/fibra.
Miofibrilele sunt constituite in principal din proteine contractile organizate in miofilamente de actina si miozina.
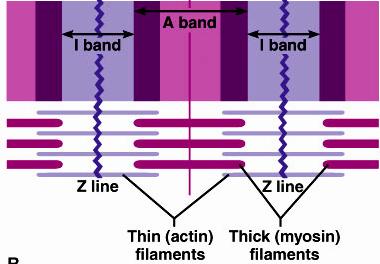
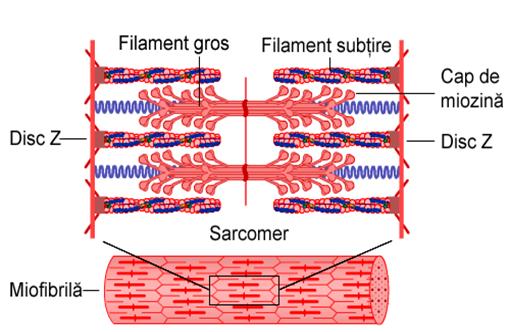
Unitatea morfo-functionala a miofibrilei este SARCOMERUL sau ,,Casuta Krause''.Un sarcomer e delimitat de doua membrane Z. La membranele Z adera miofilamentele subtiri de actina care patrund partial spre interiorul sarcomerului printre filamentele groase de miozina. In timpul contractiei, filamentele groase "trag" filamentele subtiri spre centrul sarcomerului. Astfel, sarcomerul este definit ca cea mai mica unitate contractila a fibrei musculare.
Datorita acestei orientari a miofilamentelor se observa o alternanta intre partile mai intunecate - DISCUL ANIZOTROP (discul intunecat) cu parti mai clare - DISCUL IZOTROP (discul clar).
Dimensiunile sarcomerului sunt in stare relaxata, in medie de 2-3 μ. Lungimea optima a sarcomerului este de 2-2,2μ Aceasta lungime corespunde la o presiune intracavitara de 10 mmHg.
- o lungime < 2μ presupune suprapunerea miofilamentelor in regiunea centrala, prin urmare se reduce numarul de punti actomiozinice care poate fi format. Forta dezvoltata va fi mai redusa.
- o lungime >2,5 μ presupune o distantare a miofilamentelor de actina de cele de miozina, ceea ce duce la scaderea numarului de punti actomiozinice formate, iar contractia va avea o forta mai mica.
Figura 1.
Structura sarcomerului. Banda A este discul intunecat format din miofilamente
groase de miozina intrepatrunse
cu cele de actina Banda I este discul clar format din miofilamente subtiri
de actina. Sarcomerul este delimitat de 2 membrane Z
La jonctiunea dintre benzile A si I, sarcolemma patrunde in interiorul sarcoplasmei formand o retea de tuburi transversale - tuburile in T care au in interior lichid extracelular. Tuburile in T ajung pana in vecinatatea reticulului sarcoplasmic cu care formeaza o unitate functionala numita diada. La muschiul scheletic formeaza triade.

Modificat de dr. Florin Romanescu dupa Sheffield, Getbodysmart.com, 2004
In structura fibrei miocardice se gasesc mai multe tipuri de proteine:
- proteine contractile - actina si miozina;
- proteine reglatoare (modulatoare) - tropomiozina, troponina, alfa, beta actinina si distrofina;
Distrofina este o proteina cu rol in coordonarea contractiei intre fibrele musculare prin legaturile pe care le face intre citoscheletul a 2 celule (fibre) vecine. Deficitul de distrofina este incriminat in aparitia cardiomiopatiei hipertrofice.
La nivelul citoscheletului s-au evidentiat doua proteine gigant: titina si nebulina. Nebulina nu e prezenta in miocard si muschiul neted.Titina formeaza o structura elastica care leaga filamentele de miozina de membrana Z.
Proteinele contractile:
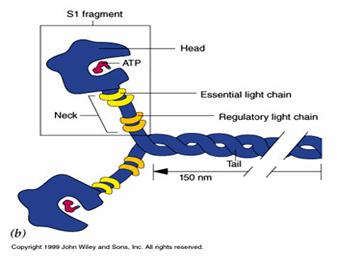
▪ MIOZINA - proteina complexa cu greutate moleculara mare (GM) - 480000. Este un hexamer format din 6 lanturi polipeptidice cu caracteristici diferite:
- 2 lanturi grele care au la un capat o portiune globulara - heavy meromiosin (meromiozina grea - HMM). Portiunea globulara are 2 capete legate de restul lantului prin 2 brate de articulare (gat) Fiecare dintre capete are un domeniu (loc, sit) motor pentru legarea ATP-ului si un domeniu pentru legarea de actina. Situl de legare pentru actina se afla pe partea opusa celui de legare a ATP. Capetele de miozina scindeaza ATP (au activitate ATPazica). Deoarece se leaga de filamentele subtiri in timpul contractiei, capetele de miozina mai sunt numite punti transversale (cross-bridges
- 4 lanturi usoare - light meromiosin (meromiozina usoara) care se dispun:
in apropierea portiunii globulare a HMM , in jurul gatului
▪ 2 la extremitatea opusa, unde se rasucesc in -helix formand "coada" miozinei.
Fiecare molecula de miozina are 2 regiuni flexibile, numite "balamale", una la baza capului si alta intre gat si coada. Acestea permit miscarea capetelor de miozina. In timpul contractiei, capetele se leaga de filamentele subtiri si le "trag" spre interiorul sarcomerului.
Cozile moleculelor de miozina, orientate spre centrul sarcomerului, formeaza miezul miofilamentului gros, capetele, impreuna cu gatul proeminand in afara, spre filamentele subtiri

Figura 2. Molecula de miozina. Portiunea cuprinsa intre sectiunea transversala si cele 2 notatii P este formata din 2 lanturi usoare rasucite in alfa helix. Portiunea globulara de Heavy meromiozina are 2 capete legate de lanturile usoare prin brate de articulare numite si "gaturi" De-a lungul fiecarui gat se gasesc alte 2 lanturi usoare, unul reglator si altul esential. Pe capete se gasesc cele 2 domenii de legare ale moleculei: un "buzunar pentru ATP sau ADP" si un loc de fixare pe actina. Capul si gatul sunt cunoscute ca subfragmentul S1, iar cele 2 lanturi usoare rasucite in alfa helix ca subfragmentul S2.

▪ ACTINA formata din monomeri globulari (actina G) care prin polimerizare formeaza doua lanturi rasucite in dublu-helix de Actina F - fibrilara. Miofilamentele de actina se intind de la discul Z pana catre centrul sarcomerului, suprapunandu-se partial cu miofilamentele groase.
Fiecare molecula de actina G are un sit activ, de legare pentru capetele de miozina. In repaus, siturile active sunt acoperite de o proteina filamentoasa, Tropomiozina. Filamentele de actina sunt ancorate la discul Z prin intermediul -Actininei.
Figura 4.
Miofilamentul de actina
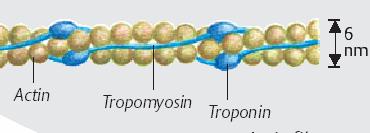
Proteine modulatoare
▪ TROPOMIOZINA. Formeaza bare alungite de-a lungul actinei F, de care se leaga prin intermediul troponinei. Fiecarei molecule de tropomiozina ii este atasata o molecula de Troponina. Moleculele de Tropomiozina se unesc cap-coada si formeaza cate 2 lanturi ce se rasucesc in jurul filamentelor de actina F. O singura molecula de Tropomiozina acopera 7 situri active de pe actina.
Rol: mascheaza locusurile active de pe molecula de actina impiedicand formarea puntilor acto-miozinice, adica legarea miozinei. Argument: daca se incubeaza actina si miozina pure impreuna cu ATP si Ca2+ se constata formarea legaturilor acto-miozinice. Introducerea in acest mediu a complexului troponina-tropomiozina blocheaza formarea acestor legaturi.
Figura 5. Relatia actina-tropomiozina si troponina in
stare de repaus..
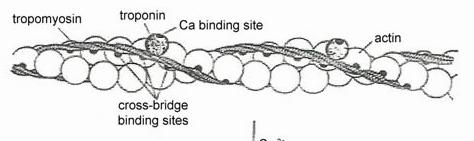
▪ TROPONINA - alcatuita din 3 componente:
- componenta I cu afinitate pentru actina: leaga complexul troponina-tropomiozina de actina si inhiba interactiunea cu miozina
- componenta T care fixeaza tropomiozina;
- componenta C cu afinitate mare pentru Ca2+ si Mg2+. Actioneaza ca un reglator al calciului intracelular.
Contractia incepe cand troponina leaga Ca2+ si tropomiozina este deplasata, descoperind siturile active de pe actina. Capetele de miozina se leaga de actina si contractia incepe.
In desfasuararea raspunsului contractil se descriu 3 etape:
I. Cuplarea excitatiei cu contractia reprezinta legatura functionala intre fenomenul electric de la suprafata sarcolemei si fenomenul mecanic intracelular - contractia. Aceasta legatura se realizeaza prin intermediul complexului diada-format din sistemul transvers in T (realizat prin invaginarea sarcolemei si o cisterna terminala apartinand sistemului sarcoplasmic longitudinal. Pentru a se facilita trasmiterea informatiei intre componentele diadei sunt prezente prelungiri jonctionale.
In stare de repaus, concentratia Ca2+ intracelular este redusa, de 10-7 M (0,1 M/L), fiind pastrata la acest nivel prin contributia a 3 proteine care pompeaza calciul in reticulul sarcoplasmic:
- calsequestrina - contribuie la captarea si pastrarea Ca2+ in reticulul sarcoplasmic.
- fosfolmban-ul - in stare fosforilata, capteaza Ca2+ in prezenta cAMP, fiind asociat cu popa Ca-Mg-ATPaza a reticulului sarcoplasmic; cand e in stare defosforilata blocheaza pompa.
- calmodulina - are afinitate pentru Ca2+ asemanatoare troponinei, contribuind in mare masura la homeostazia intracelulara a Ca2+.
II. Contractia. Sub actiunea unui stimul cu intensitate cel putin laminara se produc modificari conformationale ale canalelor rapide de Na+. Ca urmare a gradientului de concentratie si electric, Na+ patrunde masiv in celula. Intrarea Na+ in celula blocheaza activitatea pompelor de Ca2+ si ca urmare concentratia sarcoplasmica a lui creste la 10-5 M. Datorita cresterii concentratiei, Ca2+ se va fixa pe componenta C a troponinei; ca urmare, legatura troponinei I cu actina slabeste si tropomiozina se deplaseaza lateral descoperind locurile active de pe molecula de actina, permitand formarea puntilor acto-miozinice.
Tot datorita cresterii concentratiei de Ca2+, molecula de ATP de pe capul miozinei se desface prin hidroliza. Capul miozinei isi modifica pozitia la 450 permitand alunecarea filamentelor de actina si miozina, cele de actina aluneca spre centrul sarcomerului, care se scurteaza.
Acest mecanism este denumit ,,mecanism in roata dintata''sau ,,in cremaliera''si a fost descris de Huxley. El este unul dintre mecanismele larg acceptate, explicand majoritatea aspectelor privind contractia.
Relaxarea
Este o etapa care incepe odata cu stoparea intrarii Na+ in celula, moment in care pompele de Ca2+ isi reiau activitatea, pompand-ul din sarcoplasma in reticulul sacoplasmic (cisterne terminale) sau in afara celulei. Datorita scaderii concentratiei Ca2+ acesta se va desprinde de pe troponina, complexul troponina-tropomiozina revine la pozitia initiala, blocand locurile active de pe actina.
Relaxarea este o faza activa care are loc cu consum energetic, de aceea in absenta substantelor macroergice - ATP, relaxarea se face dificil, putand aparea starea de contractura (ex: la deces - rigor mortis).
Relatia dintre lungimea initiala si forta de contractie a fost studiata de catre Frank si Starling pe preparat cord-pulmon formuland ,,legea inimii". S-a observat ca exista o relatie de directa proportionalitate intre lungimea initiala a fibrei si forta de contractie pe care o dezvolta in sistola urmatoare. Alungirea initiala a fibrelor miocardice este estimata prin volumul telediastolic (volumul care se afla la sfarsitul diastolei) si chiar prin presiunea telediastolica, iar forta de contractie prin volumul sistolic ejectat.
S-a observat o relatie de directa proportionalitate intre cele doua marimi: pe masura ce fibra isi mareste lungimea initiala poate sa dezvolte o tensiune mai mare, proportionala cu alungirea. Acesta relatie se pastreaza pana la o alungire a fibrei de 120-130% din lungimea initiala. Peste aceasta valoare a alungirii, forta dezvoltata nu mai creste, ci scade.
Mecanismul Frank-Starling este un mecanism de adaptare heterometric, la care, in mod normal cardul nu apeleaza decat foarte putin pentru echilibrarea debitelor celor doi ventriculi. El nu intervine in mod normal deoarece eficienta adaptativa este redusa. La acest mecanism, cordul apeleaza doar in cursul unor eforturi epuizante cand posibilitatile de adaptare ale SNV sunt epuizate si in patologie (insuficiente cardiace -,,cord bovin")
METABOLISMUL MIOCARDULUI
Energia rezultata din metabolismul miocardului va fi folosita
pentru cresterea tensiunii din fibre;
pentru scurtarea fibrelor
pentru imprimarea unei viteze de curgere a coloanei de sange.
Din intreaga cantitate de energie, 90% este utilizata pentru asigurarea contractiilor, iar restul de 10% este folosita pentru diverse sinteze necesare mentinerii integritatii structurale, pentru activitatea pompelor ionice, a diferitelor sisteme enzimatice.
Stocarea energiei se realizeaza in substante macroergice: ATP, ADP si CP (creatinfostat).
Ca surse energetice, miocardul apeleaza atat la surse exogene, aduse prin sangele care circula prin vasele coronare, cat si la surse endogene reprezentate de depozitele energetice de la nivelul fibrelor.
Spre deosebire de muschii scheletici, care consuma numai substrat glucidic, miocardul mai poate metaboliza si: acid lactic, acizi grasi liberi, corpi cetonici, aminoacizi. In cursul efortului fizic creste utilizarea de catre cord a lactatului rezultat in urma travaliului muschilor scheletici.
Prin glicoliza aeroba se obtin 36 moli de ATP, iar prin glicoliza anaeroba 2 moli ATP.
La nivelul miocardului se desfasoara preferential glicoliza aeroba - ciclul Krebs. De aceea consumul de O2 al miocardului reprezinta 7-8% din consumul de O2 al organismului.
In conditii de hipoxie, metabolismul anaerob poate sa ajunga pana la 10%. In totala anaerobioza energia este insuficienta pentru a asigura travaliul ventricular!
La nivelul miocardului, oxiHb se disociaza in proportie de 65-70% asa cum se disociaza la celelalte tesuturi in efort. Cordul nu este niciodata in repaus ci mereu in efort!
Dintre subtantele metabolizate la nivelul miocardului:17% sunt glucide,17%acid lactic+piruvic, 57% acizi grasi, ~5% corpi cetonici, 4%aminoacizi.
La nivelul miocardului exista un mecanism important de modificare a circulatiei coronariene si respectiv aprovizionare cu O2 si substante nutritive. In efort se desface o cantitate marita de ATP, rezultand in final o cantitate crescuta de adenozina, o substanta putenic vasodilatatoare la nivelul circulatiei coronarieneàastfel mareste fluxul sanguin la miocard.
ANATOMIA FUNCTIONALA A CORDULUI
Inima fuctioneaza ca o pompa aspiro-respingatoare, care aspira sangele din sistemul venos si-l pompeaza in cel arterial.
La exterior, cordul este continut intr-un sac seros dublu, sacul pericardic; intre foitele acestuia se gaseste o pelicula de lichid care permite alunecarea foitelor intre ele, facilitand miscarile cordului. Un alt rol al pericardului este sa limiteze expansiunea distolica a cordului evitandu-se astfel supraincarcarea cu sange a micii circulatii, precum si aparitia unei insuficiente valvulare functionale determinata de marirea, dilatarea orificiilor valvulelor A-V.
In hemodinamica, rolul pericardului nu este foarte important. Ca argument: la persoanele cu absenta congenitala a pericardului nu se inregistreaza modificarea hemodinamicii.
Inima are forma de trunchi de con pozitionat cu baza superior si varful inferior in torace. Cele 4 cavitati au pozitii si forme diferite, iar miocardul este format din straturi diferite cu dispozitii caracteristice :
Atriile au forma de calota sferica si se insera pe partea superioara a inelelor atrioventriculare - mitral si tricuspidian; AD are o pozitie antero-laterala, iar AS are o pozitie postero-superioara.
Miocardul atrial e format din 2 straturi de muschi: unul superficial cu dispozitie transversala, comun celor 2 atrii si unul profund propriu fiecarui atriu format din fibre care se dispun perpendicular unele fata de altele. Fiecare atriu au un "apendice" atrial care se numeste "urechiuse"
Ventriculii:- VS are o forma cilindrica, cu un varf conoid;
- VD este aproape sferic, cu concavitatea spre sept (forma care sugereaza semiluna).
Aceste forme sunt adaptative, la regimuri presionale diferite si nu conditionate embriologic. Astfel forma VS, care are o raza mica este adaptata la presiuni mari, iar cea a VD la presiuni mici, acest ventricul avand o raza mai mare. Argument: Legea Laplace: P = T/r, in care P=presiunea, T= tensiunea peretilor, r=raza. Datorita razei mici, presiunea intraventriculara poate atinge valori de 250-300 mmHg la nivelul VS, astfel VS este adaptat sa evacueze volumul sistolic chiar contra unor rezistente periferice crescute.
Miocardul ventricular este format din 3 straturi: fibre spirale - pozitionate in 2 straturi: stratul spiral extern si intern si fibre constrictorii, circulare.
Straturile spirale formeaza cea mai mare parte a peretelui apexului. Cand se contracta cele doua straturi spirale, contractia startului spiral extern genereaza o tractiune oblica, ce va fi contracarata de tractiunea oblica exercitata de stratul spiral intern. Rezultatul global al contractiei: se produce scurtarea in sens longitudinal a camerei de ejectie si nu torsionarea acesteia.
Stratul constrictor se interpune intre cele doua straturi spirale si formeaza o parte din septul interventricular si peretele lateral al VS. Este putin reprezentat la nivelul VD. Prin contractia sa la VS produce o micsorare a diametrului transversal cu o scurtare minima de a apex la baza. La VD produce mai mult o scurtare in sens longitudinal.
Datorita acestor caracteristici, VD este o pompa eficienta mecanic doar la volume mari, debite mari si presiuni mici. Atunci cand aceste conditii de lucru se modifica se observa si o schimbare a caracteristicilor structurale ventriculare: peretele se va ingrosa si din forma de fragment de sfera va lua forma cilindrica.
Pe suprafata endocavitara, fibrele musculare ventriculare formeaza muschii papilari care se termina fiecare cu cate un cordaj tendinos care se insera pe aparatul valvular atrio-ventricular. Exista 2 muschi papilari in VS si 3 in VD. Muschii pilieri au rolul de a asigura etansietatea sistemului valvular atrio-ventricular in sistola.
APARATUL VALVULAR
Dispune de un aparat valvular care asigura circulatia in sens unic: dinspre atrii spre ventricule si din ventricule spre artere. Sunt descrise doua sisteme valvulare: sistemul atrio-ventricular si sistemul sigmoidian.
Sistemul atrio-ventricular este reprezentat de cele doua valvule A-V, dispuse intre A si V de aceeasi parte.
Intre AS si VS se gaseste valvula bicuspida/mitrala. Este reprezentata de o cuspida anterioara mai mare si o cuspida posterioara mai redusa. Valva mitrala suporta variatii presionale mari, diferentele(gradientele) de presiune sunt cam de 120 mmHg. Orificiul mitral are forma unui trunchi de con,Øsup=32 mm,Øinf=26 mm.
Intre AD si VD se gaseste valvula tricuspida, care are 3 cuspide fiecare avand o margine libera in contact cu cordajele tendinoase si muschii papilari. Orificiul tricuspidei este cilindric cu Ø=35 mm. Gradientul presional suportat de valvula tricuspida este ~30 mmHg
Pentru sistemul valvular A-V este caracteristica legatura cu cordajele tendinoase si muschii papilari. Datorita acestor legaturi, cuspidele valvulare sunt pozitionate corect in cursul sistolei, asigurand etansitatea orificiului, iar in diastola asigura deschiderea corecta, completa a lor.Tot datorita acestor legaturi, in sistola este impiedicata bombarea anormala a valvulelor spre atrii.
Daca se rup cordajele, sau muschii papilari, cuspidele colabeaza (bombeaza) spre atrii, inregistrandu-se prolapsul (cel mai frecvent este prolaps mitral)
Activitatea de inchidere si de deschidere a valvulelor A-V se face in functie de gradientul presional. Astfel, valvulele A-V se deschid la inceputul sistolei ventriculare, cand presiunea intraventriculara depaseste presiunea intratriala. Deschiderea valvulelor A-V va avea loc la sfarsitul relaxarii izovolumetric cand presiunea intraventriculara devine inferioara presiunii intraatriale.
Sistemul sigmoidian este reprezentat de valvulele sigmoide, prezente la baza arterelor mari. Sunt formate din 3 cuspide, avand forma de buzunar sau cuib de randunica. Orificiul aortic si pulmonar are Ø 20-23 mm.
Deschiderea valvulelor sigmoide se produce la sfarsitul sistolei izovolumetrice cand presiunea intraventriculara egaleaza si chiar depaseste presiunea diastolica din arterele mari. In timpul ejectiei sangelui, valvele nu se lipesc de peretii arteriali, la nivelul aortei permitandu-se astfel desfasurarea circulatiei coronariene.
VOLUME SI PRESIUNI
Inima lucreaza cu o gama destul de larga de debite: de la 3-4 l/min in conditii de repaus, pana la 30 l/min, chiar peste in conditii de efort.
Cordul are un regim presional cuprins intre 0 (chiar -2) mmHg si 250-300 mmHg.
Proprietatile sale vascoelastice ii permit ca in diastola sa primeasca volume relativ mari de sange fara modificari importante ale valorilor presionale.
Volumul de sange din atrii este mai mare decat volumul ventricular, constituind un rezervor pentru cazurile de crestere rapida, brusca a debitului cardiac.
Astfel, la nivelul:AD-volumul mediu~160 cm3
AS-volumul mediu ~140 cm3
VD-volumul mediu~140 cm3
VS-volumul mediu~120 cm3
Prin metode radiologice, echocardiografice Doppler, s-a putut observa ca din volumul sanguin ventricular, in fiecare sistola se evacueaza doar un volum de 40-45% reprezentand ,,volumul sistolic",,,volumul bataie" sau ,,stroke volume", cu o valoare de ~70 ml. Ceea ce ramane in ventricul la sfarsitul sistolei-VTS, cu o valoare de 60-70 ml
|
Cavitatea |
Diastola |
Sistola |
|
Atriul stang |
0- la -2 mmHg |
6-8 mmHg |
|
Atriul drept |
0, chiar valori negative |
-6 mmHg |
|
Ventricul stang |
0, chiar valori negative |
Aprox. 120 |
|
Ventricul drept |
0, chiar valori negative |
mmHg |
|
Aorta |
mmHg |
mmHg |
|
Artera pulmonara |
mmHg |
mmHg |
DEBITUL CARDIAC
=cantitatea de sange ejectata de VD si VS in mica si marea circulatie la interval de 1 min.
Debitul cardiac se va obtine din produsul dintre volumul sistolic si frecventa cardiaca.
Qc=VS x FC
Volumul sistolic - in mod normal, in repaus: 60-70 ml
Volumul care ramane in ventriculi la sfarsitul sistolei=volum telesistolic (VTS) = volumul de rezerva pe baza caruia creste volumul sistolic la cresterea fortei de contractie ~60 ml. Exista si un volum rezidual: ~10 ml care nu poate fi eliminat, ramane printre muschii pilieri
Volumul telediastolic (VTD)=cantitatea de sange existenta in ventriculi inaintea inceperii sistolei ventriculare ~120-140 ml.
Volumul sistolic este dependent de contractilitate, tonusul sistolic si postsarcina (rezistenta la evacuare).
VTD ete influentat de tonusul diastolic ventricular, de presiunea eficienta de umplere si de timpul de umplere-dependent de frecventa cardiaca.
Postsarcina e reprezentata de tensiunea arteriala de la nivelul aortei si arterei pulmonare.
Frecventa medie in conditiile ritmului sinusal este la barbati ~78 c/min, la femei ~84 c/min.
Relatia dintre FC (frecventa cardiaca) Qc este diferita la cordul izolat si la cordul,,in situ''.
La cordul izolat, pastrandu-se constanta forta de contactie si presiunea de umplere, se observa ca scaderea FC sub 60 b/min duce si la scaderea Qc. La o FC intre 60-100b/min, cresterea FC determina proportional cresterea Qc. Peste 100 c/min, Qc incepe sa scada, dar modificarile sunt nesemnificative. Scaderi marcante se realizeaza la FC >200 b/min.
La cordul ,,in situ'', cand creste FC se observa cresterea fortei de contractie si a presiunii de umplere, aceste modificari producand si cresterea vitezei de umplere ventriculara. Astfel, odata cu cresterea FC se inregistreaza si cresteri in paralel ale Qc pana la o limita de frecventa =170 b/min pentru persoanele neantrenate si 200 b/min pentru persoanele antrenate.
In efort, Qs poate creste pana la valori de 120 de ml, producand astfel, alaturi de cresterea frecventei, cresterea Qc la valori de 25 - 30 L/min.
La atletii de performanta, Qs poate creste pana la 170 - 180 ml/bataie, iar Qc poate ajunge la 38-40 de L/min.
FAZELE CICLULUI CARDIAC FIZIOLOGIC
Ciclului cardiac reprezinta succesiunea de modificari de volum si presiune din cavitatile cordului.
Un ciclu cardiac cuprinde 2 faze: sistola-contractie si diastola-relaxare
Activitatea electrica cardiaca o precede pe cea mecanica (contractia) cu 0,02-0,04 sec. Activitatea cordului drept precede activitatea cordului stang cu 0,02 sec (cordul drept este primul care se depolarizeaza). Valvele cordului stang se inchid mai repede si se deschid mai tarziu comparativ cu valvulele cordului drept.
La inceputul sistolei ventriculare, cand presiunea intraventriculara incepe sa creasca, depasind-o pe cea a atriilor, se produce inchiderea valvulelor A-V, adica Mi,Ti si incepe sistola ventriculara izovolumetrica, perioada in care ventriculii sunt cavitati inchise, cu volum constant. In acest moment prin punerea in tensiune a fibrelor, creste tensiunea intraventriculara, egaland si chiar depasind presiunea distolica din arterele mari. Acum se deschid valvele sigmoide, pulmonara si aortica: Pd, Ad si incepe urmatoarea faza a sistolei ventriculare - sistola izotona, cu o perioada de ejectie rapida, urmata de ejectia lenta. La sfarsitul ejectiei lente, presiunea intraventriculara este egala cu cea din arterele mari, ca urmare, coloana de sange se mai deplaseaza spre arterele mari prin inertie. Aceasta este perioada de protodiastola fiziologica.
La sfarsitul protodiastolei fiziologice, datorita tendintei de scadere a presiunii intraventriculare se inchid valvulele sigmoide: Ai,Pi. Din nou ventriculii sunt cavitati inchise iar prin relaxarea miocardului, presiunea intraventriculara scade foarte mult, fenomen ce caracterizeaza relaxarea izovolumetrica. Presiunea intraventriculara atinge valori minime presionale de 0 sau chiar valori negative, inferioare presiunii atriale si ca urmare se deschid valvulele A-V, in ordinea Td,Md si incepe diastola ventriculara, cu o faza de umplere rapida si una de umplere lenta, dupa care se produce din nou sistola atriala.
Sistola atriala, cu o durata de 0,11-0,15 sec. e caracterizata prin cresterea usoara a presiunilor intraatriale expulzandu-se o cantitate suplimentara de sange din A in V. Contributia sistolei atriale e apreciata la 10%din umplerea ventriculara (in efort 30%).
Duratele fazelor ciclului cardiac sunt dependente de frecventa cardiaca. La cresterea frecventei,
cel mai putin afectate sunt contractia si relaxarea izovolumetrica, iar cea mai afectata este umplerea lenta, consecinta fiind o umplere mai mica a ventriculilor si deci un debit sistolic mai mic
La o frecventa medie de 80 contractii/min, ciclul cardiac dureaza 0,8 sec dintre care:
- 0,3 sec sistola ventriculara;
- 0,5 sec diastola ventriculara;
Sistola atriala 0,1 sec
Diastola atriala 0,7 sec
Diastola generala 0,4 sec.
ZGOMOTELE CARDIACE
Apar in timpul activitatii cardiace si pot fi ascultate prin plasarea directa a urechii pe torace sau cu stetoscopul. Inregistrarea zgomotelor cardiace se numeste fonocardiografie.
Sunt caracterizate prin: intensitate (depinde de amplitudinea oscilatiei sonore si se masoara in decibeli (dB); frecventa sau tonalitate (Hz); durata si timbrul, care depinde de undele armonice asociate.
Frecventa medie a zgomotelor cardiace este de 20-250 Hz, acesta fiind banda oscilatorie a lui Luisada. Urechea umana percepe sunete intre 20 si 20000 HZ. Intensitatea lor ajunge pana la 40 dB, zgomotul de fond al unei incaperi in conditii normale fiind de 70 dB. Amplitudinea maxima a zgomotelor se inregistreaza la o frecventa de 24 Hz, foarte aproape de limitele audibilitatii. La frecvente >500 Hz se reduce amplitudinea oscilatiilor (deci intensitatea scade) facandu-le inaudibile.
Geneza zgomotelor cardiace:
- activitatea valvulara - inchiderea si deschiderea valvelor. Cel mai mult contribuie inchiderea valvelor deoarece se face in sens invers curgerii sangelui, determinand curenti turbionali.
- vibratiile peretilor vaselor mari, mai ales in sistola cand primesc coloana de sange;
- coloana de sange in miscare;
- contractia miocardica.
In activitatea cardiaca se descriu urmatoarele zgomote:
♦ sistolice: zgomotul I si II
♦ diastolice: zgomotul III si IV
Zgomotul I
- marcheaza inceputul sistolei;
- apare dupa complexul QRS la 0,02-0,04 sec;
- are o frecventa medie ~70 Hz, mai mica decat a zgomotului II datorita elasticitatii mai mici a peretilor ventriculari, comparativ cu cea a arterelor mari.
- durata 0,08-0,12 sec;
- intensitatea zgomotului I este direct proportionala cu marimea gradientului de presiune prin sistemul valvular. Cand creste rapid forta de contractie a inimii va creste si presiunea intraventriculara, deci si gradientul presional prin valva ceea ce va duce la o intensitatea crescuta a zgomotului I.
- este format din presegment, segment principal si postsegment. Segmentul principal este dat de componentele MiTi si PdAd
Cea mai mare parte din zgomotul I este dat de componenta valvulara, in special de mitrala inchidere (aproximativ 80%). De aceea zgomotul I este mai accentuat in focarele de auscultatie de la varful inimii.
Zgomotul II: marcheaza sfarsitul sistolei
- durata - 0,06-0,08 sec. mai mica deoarece vibratiile produse de valvulele sigmoide sunt mai rapid tamponate de peretii arteriali, elastici
- intensitatea zgomotului II depinde de viteza cu care scade presiunea in ventricul. Aceasta viteza este dependenta de presiunea atinsa in sistola. Atunci cand se inregistreaza HTA (se ating valori de ~200 mmHg), presiunea intraventriculara scade cu o viteaza de 2 ori mai mare decat in cazul unei presiuni normale. De aceea, in HTA zgomotul II va avea o intensitate mai mare (hipertensiune atat sistemica cat si pulmonara). In hipotensiune sau insuficienta cardiaca, intensitatea zgomotului II va fi diminuata.
- apare la 0,02-0,04 sec dupa unda T.
- are frecventa mai mare decat zg I, 120-130 Hz;
- compus din presegment, segment principal, postsegment.
Presegmentul - apare inconstant si se datoreaza vartejurilor formate de coloana de sange in miscare.
Segmentul principal - este dat de componentele valvulare Ai,Pi.
Postsegmentul - dat de deschiderea valvelor atrioventriculare : TdMd.
Cea mai mare parte a zgomotului II e data de componenta Ai - de aceea se aude mai bine in focarele de la baza inimii.
Zgomotele III si IV sunt zgomote diastolice
Zgomotul III: apare la 0,12-0,18 sec dupa zgomotul II (~13 sec);
- are amplitudine mica, frecventa joasa;
- se produce in timpul umplerii rapide;
- durata:0,04-0,06 sec;
- se aude si se inregistreaza la persoanele tinere cu peretele toracic subtire, fiind fiziologic pana la 20 de ani.
Zgomotul IV - apare la 0,02-0,04 sec dupa unda P;
- amplitudine mica, frecventa joasa, durata mica(0,04 sec);
- se produce in timpul sistolei atriale;
Inregistrarea zgomotelor cardiace se face prin Fonocardiografie, iar graficul obtinut se numeste fonocardiograma
Focare de ascultatie:
- Focarul mitral - spatiul V, intercostal stanga la intalnirea cu linia medioclaviculara (usor inauntrul acestei linii)
- Aria mezocardica - la nivelul spatiului IV parasternal stanga;
- Focarl Erb - in spatiul III intercostal parasternal stanga;
- Focarul tricuspidian - spatiul IV intercostal parasternal dreapta sau mediosternal la baza apendicelui xifoid;
- Focarul aortic - spatiul II parasternal drept;
- Focarul pulmonar - spatiul II parasternal stang.
Zgomotele cardiace pot fi accentuate, diminuate, dedublate si se pot inregistra si fenomene sonore supradaugate.
Accentuarea - fiziologica: la tineri cu perete toracic subtire, in tahicardia din efortul fizic sau stari emotionale.
- patologica in HTA, HT pulmonara.
Diminuarea - fiziologica: la obezi, la batrani unde intervine un grad de emfizem pulmonar.
- patologica: hipotensiune, insuficienta cardiaca, colectii pleurale si pericardice.
Dedublari:- fiziologice - dedublarea zgomotului II. Atunci cand decalajul intre inchiderea aortei si pulmonarei este pana la 0,02 sec, se aude un singur zgomot. Cand acest decalaj este mai mare de 0,02 sec se aud distinct inchiderea aortei si inchiderea pulmonarei. Acesta dedublare apare in inspir si dispare in expir-de aceea e fiziologica si se datoreaza cresterii afluxului venos in timpul inspirului. De aceea, timpul de ejectie al VD creste, marind intervalul dintre Ai si Pi.
- patologice - dedublarile zgomotelor I, II se percep atat in inspir cat si in expir chiar daca in inspir dedublarea e mai accentuata
Fenomene sonore supraadaugate- clicuri si sufluri.
Clicurile sunt fenomene sonore cu durata sub 0,04 sec. Se percep ca si componente distincte ale zgomotelor I sau II si se produc la deschiderea unor valve sclerozate, de exemplu deschiderea valvei mitrale in stenoza mitrala sau a valvei aortice in stenoza aortica.
Suflurile sunt fenomene sonore cu durata peste 0,12 sec. Ele apar datorita transformarii curgerii laminare in curgere turbulenta.
La tineri se pot intalni curgeri turbulente la nivelul aorei ascendente in perioada ejectiei rapide, cand se produc suflurile sistolice functionale.
In patologie apar cel mai frecvent in patologia valvelor atrioventriculare sau sigmoidiene, care poate fi de tip stenoza sau insuficienta valvulara.
Pentru localizarea precisa in sistola si diastola clinica, s-a facut impartirea in 3 parti egale.
Sistola clinica (pauza mica) intre zgomotul I si II: cuprinde proto, mezo si telesistola.
Diastola clinica (pauza mare) intre zgomotele II si III: cuprinde proto, mezo si telediastola.
REGLAREA ACTIVITATII CARDIACE SI A CIRCULATIEI
Activitatea cordului si vaselor trebuie reglata pentru a se asigura un aport adecvat de sange in conditii de lucru diferite induse de modificarile de mediu, de stress, de efortul fizic.
Aceasta implica:
o reglare optima a activitatii cardiace si a presiunii arteriale;
o perfuzie adecvata a tuturor organelor;
dirijarea sangelui spre organele implicate activ intr-o activitate (de ex. muschii in efortul fizic) si suntarea celor care sunt in repaus in acel moment.
Adaptarea cordului si vaselor la conditii de lucru diferite se face prin interventia:
▪ sistemului nervos vegetativ si in acest caz vorbim de reglarea nervoasa a activitatii cordului si vaselor;
▪ unor substante ce rol de hormoni sau mediatori si in acest caz vorbim de reglarea umorala;
▪ unor factori locali (de natura miogena) sau proveniti din metabolismul celular care actioneaza in acelasi loc in care au fost eliberati si in acest caz numim acest tip de reglare ca fiind locala; multi autori definesc mecanismele reglatorii de acest tip ca autoreglarea circulatiei.
Un prim parametru care trebuie ajustat la necesitatile diferite impuse de modificarile de mediu este debitul cardiac Qc sau, in engleza CO=cardiac output, adica volumul de sange pompat de inima intr-un minut. Debitul cardiac depinde de debitul sistolic (Qs) si frecventa cardiaca (FC), adica Qc = Qs x FC. Debitul sistolic este definit ca volumul de sange expulzat de cord intr-o sistola. Se mai numeste si volum/bataie sau, in engleza SV-stroke volume.
In situatia in care debitul cardiac trebuie crescut, ca in cazul efortului fizic, aceasta crestere se face prin marirea volumuli sistolic (prin cresterea fortei de contractie) si a frecventei cardiace.
Debitul sistolic depinde de volumul telediastolic (VTD) si de volumul telesistolic (VST) sau Qs = VTD - VTS. Debitul sistolic este reglat prin mecanisme intrinseci si extrinseci, ambele modificand forta de contractie si presiunea de ejectare a sangelui.
Mecanismele intrinseci de reglare a debitului sunt reprezentate de:
▪ mecanismul heterometric descris de Frank-Starling de adaptare la presarcina (mecanism care nu asigura componenta presionala, fiind un mecanism fiziopatologic de adaptare la solicitari) modificarile debitului cardiac fiind determinate de alungirea fibrelor cardiace: forta de contractie si implicit Qc creste proportional cu gradul de alungire initiala a fibrelor (legea inimii)
▪ mecanismul homeometric Anrep de adaptare la postsarcina - la aceeasi lungime initiala a fibrelor, forta de contractie este influentata de frecventa cardiaca si temperatura.
Ambele mecanisme au o contributie scazuta la adaptarea debitului la necesitati crescute, predominante fiind mecanismele nervoase si umorale.
Efecte inotrop pozitive si cronotrop pozitive exercita:
▪ sistemul nervos vegetativ simpatic prin intermediul adrenalinei (epinefrina), care prin actiunea asupra receptorilor β1 adrenergici determina: cresterea frecventei cardiace prin cresterea permeabilitatii membranare pentru Ca2+ si Na+ la nivelul celulelor P, cresterea fortei de contractie prin favorizarea intrarii Ca2+ in perioada de platou a potentialului de actiune si coronarodilatatie, iar prin noradrenalina (norepinefrina) determina vasodilatatie si scaderea rezistentei opusa la curgerea sangelui in periferie;
▪ distensia musculaturii atriale care stimuleaza celulele pace-maker;
▪ ionul de Ca2+;
▪ hipokalemia moderata are efect cronotrop pozitiv ;
▪ cresterea temperaturii sangelui;
▪ cofeina, hormonii tiroidieni.
Efecte inotrop negative si cronotrop negative exercita:
▪ hipoxia coronariana accentuata;
▪ hipercapnia (cresterea CO2);
▪ acetilcolina;
▪ excesul de K+, Mg2+, barbiturice, etc.
1. Mecanismele nervoase de reglare a activitatii cordului si vaselor
Aceste mecanisme sunt reflexe si presupun parcurgerea unui arc anatomic reflex care cuprinde:
▪ receptorii care sunt niste traductori sau senzori care preiau modificarile fizice, chimice sau biologice si le convertesc in diferente de potential;
▪ caile nervoase aferente - fibre nervoase senzitive care transporta stimulii (potentiale de actiune) la centrii de control din SNC;
▪ centrii cardiovasculari la nivelul carora se face interpretarea si analiza informatiilor primite si se elaboreaza un raspuns, o comanda adaptata la nevoile organismului;
▪ cai eferente - fibre nervoase vegetative motorii simpatice sau parasimpatice care vehiculeaza comanda la organele efectoare care sunt cordul si vasele.
Receptorii implicati in reglarea hemodinamicii sunt prezenti in interiorul aparatului cardiovascular (baro- si chemoreceptori), dar si in afara acestuia (receptori alveolari, receptori pentru durere, proprioreceptori, receptori mezenterici).
Receptorii din sistemul cardiovascular sunt prezenti la orice nivel, dar cei mai importanti functional sunt cei situati la nivelul unor zone reflexogene "strategice": sinocarotidiana, cardio-aortica (crosa aortei), a venelor mari, a cavitatilor cordului; au rol de traductori ai modificarilor presionale - baroreceptori si/sau ai modificarilor compozitiei biochimice a sangelui - chemoreceptori.
a. Baroreceptorii arteriali se gasesc in crosa aortica - la emergenta arterelor subclaviculare - si in sinusul carotidian, deasupra bifurcatiei carotidei primitive, pe carotida interna. Ei sunt stimulati de distensia peretilor arteriali, produsa cel mai frecvent de cresterea tensiunii arteriale sistemice.
Cresterea presiunii arteriale sistemice (sau compresiunea carotidelor deasupra bifurcatiei) mareste frecventa impulsurilor transmise de acesti receptori la un nivel proportional cu cresterea presiunii (in fapt, cu gradul de distensie al peretelui). Aceste impulsuri ajung pe cai aferente la centrul cardio-inhibitor din bulb si determina scaderea debitului cardiac (prin scaderea frecventei si a contractilitatii cardiace) si scaderea rezistentei vasculare si deci si a presiunii (reflex depresor).
In sens opus, o presiune scazuta are efect invers determinand readucerea presiunii la valori normale.
Arcul reflex al reflexelor depresoare cu punct de plecare in crosa aortica si sinusul carotidian.
▪ Baroreceptorii sinocarotidieni stimulati de distensia peretelui ca urmare a cresterii presiunii arteriale sau de compresia sinusului carotidian prin manevre vagale ►► cai aferente reprezentate de fibre ale glosofaringianului (nervul Hering) ►► centrul cardioinhibitor din bulb si vasodepresor din bulb si punte ►► cai eferente reprezentate de nervul vag care se distribuie efectorilor: nodulul sinoatrial (scade frecventa cardiaca), miocardul contractil care raspunde prin scaderea fortei de contractie si de ci a debitului sistolic si musculatura neteda din vasele periferice care raspund prin vasodilatatie. Efectul: scade debitul cardiac ca urmare a reducerii frecventei si debitului sistolic, vasodilatatie si revenirea la valori normale a presiunii arteriale
▪ Baroreceptorii de la nivelul crosei aortice stimulati de distensia peretelui ca urmare a cresterii presiunii arteriale ►► cai aferente reprezentate de vag (nervul Cyon-Ludwig) ►► centrul cardioinhibitor din bulb si vasodepresor din bulb si punte ►► cai eferente reprezentate de nervul vag; efecte asemanatoare cu cele declansate de stimularea baroreceptorilor sinocarotidieni.
b. Baroreceptorii atriali (situati subendocardic, endocardul atrial fiind zona cardiaca cu cea mai bogata inervatie) sunt de tip A care sunt stimulati de contractia atriala si de tip B stimulati de intinderea pasiva din timpul intorcerii venoase. Frecventa de descarcare a receptorilor atriali depinde direct proportional de presiunea venoasa.
Stimularea receptorilor din atriul drept produce tahicardie prin reflexul Bainbridge (1915) realizat pe cai aferente vagale si cai eferente simpatice, ce influenteaza frecventa de descarcare a nodulului sinoatrial. Reflexul Bainbridge are rolul de a preveni acumularea sangelui in vene, atrii si in circulatia pulmonara. Distensia atriului stang (AS) produce un raspuns asemanator, avand aceeasi explicatie.
c. Baroreceptorii ventriculari sunt reprezentati de mecanoreceptori, stimulati de distensia ventriculara, mai putin numerosi decat cei atriali (evidentiati subendocardic si subepicardic in ambii ventriculi). Ei produc efecte depresoare, dar numai in cazul destinderilor foarte mari ale ventriculului stang.
d. Baroreceptorii din circulatia pulmonara sunt localizati in adventicea trunchiului arterei pulmonare si a ramurilor dreapta si stanga ale acesteia si sunt stimulati de distensia patului vascular pulmonar, determinand vasodilatatie cu hipotensiune arteriala si bradicardie. In conditii experimentale acesti receptori pot fi stimulati de injectarea in circulatia pulmonara a veratridinei, fenilbiguanidei sau a serotoninei. Stimularea receptorilor pulmonari produce efecte de tip depresor.
b. Chemoreceptorii sunt localizati in special in zona sinocarotidiana si cardioaortica (crosa aortica) - sunt stimulati de modificarile presiunilor partiale ale gazelor si de modificarea pH-ului sanguin. Stimulul principal este reprezentat de scaderea pO2, stimularea lor facandu-se si de cresterea pCO2. Chemoreceptorii aortici, spre deosebire de cei sinocarotidieni, nu sunt activati de modificarea pH-ului, fiind chiar deprimati.
Cresterea temperaturii are efect stimulant asupra chemoreceptorilor, iar scaderea temperaturii efect inhibitor.
Raspunsul cardiac la stimularea chemoreceptorilor sino-carotidieni si cardio-aortici: stimularea chemoreceptorilor prin cresterea pCO2 sau scaderea pO2 determina cresterea ventilatiei si secundar, cresterea frecventei cardiace si a presiunii sanguine.
Alte zone cu chemoreceptori: terminatiile vaselor coronare, ventriculul stang, circulatia pulmonara.
Stimularea chemoreceptorilor situati la nivelul terminatiilor vaselor coronare:
▪ stimuli: toxice endogene, cataboliti acizi (ex. IM, obstruare experimentala a unei coronare)
▪ efect: declansare reflex depresor;
▪ aferenta: fibre senzitive ale vagului;
▪ centru: cardioinhibitor din bulb;
▪ eferenta: fibre ale vagului.
Reflexul Bezold - Jarisch injectarea de veratridina sau de de nicotina in ramurile arterelor coronare care iriga ventriculul stang, produce bradicardie, vasodilatatie si hipotensiune arteriala pana la colaps, prin stimularea chemoreceptorilor ventriculari. Declansarea acestui reflex in infarctul acut de miocard are efect protector pe inima lezata, dar poate fi nociv daca e prea puternic (stop cardiac, prabusirea TA).
Veratridina - alcaloid extras din Veratrum viride Ait. Liliaceae si Veratrum album L. Liliaceae. Are efecte parasimpaticomimetice (asemanatoare cu ale parasimpaticului) inducand hipotensiune. La doze mari deprima respiratia si produce aritmii.
Nicotina - alcaloid extras din frunzele de tutun. Activeaza receptorii colinergici pentru nicotina. Este un acetilcholinomimetic cu efecte stimulante asupra sistemului nervos. Creeaza dependenta, in acest caz se constituie intr-o adevarata "otrava neuronala".
Chemoreceptorii din ventriculul stang
- stimul: hipoxie si acidoza;
- efect: reflex presor (creste TA, creste FC, vasoconstrictie);
- aferente: simpatice; centrii: medulari vasoconstrictori si cardioacceleratori.
Chemoreceptorii din circulatia pulmonara (chemoreflexul pulmonar)
- stimul: hipoxie, experimental injectarea de veratridina sau serotonina ;
- efect: reflex depresor (scade FC, hTA, vasodilatatie);
- aferenta: vagala;
- centrii: cardioinhibitor si vasodepresor.
Si chemoreceptorii centrali sunt implicati in reglarea activitatii cordului si vaselor - cei mai cunoscuti sunt cei bulbari, situati pe fata ventrala, in apropierea radacinilor nervilor cranieni IX, X si XI. Sunt sensibili la modificarea pH-ului extracelular si a lichidului cefalorahidian (LCR).
Alti receptori implicati in reglarea activitatii cordului si vaselor: algoreceptori, termoreceptori, osmoreceptori, proprioreceptori.
Algoreceptorii: -stimul: durerea ???;
-efect: reflexe de tip presor sau depresor daca stimulul e de intensitate
foarte mare.
TA creste in durerea cutanata si scade in durerea viscerala, musculara sau arteriala!!!
Termoreceptorii:
stimul: temperaturi f. Scazute;
- efect: reflex presor sau depresor daca stimulul e foarte puternic.
Baroreceptorii mezenterici sunt mecanoreceptori cu structura asemanatoare corpusculilor Pacini, cu rol posibil in reglarea circulatiei viscerale si care, prin stimulare, initiaza reflexe cardiovasculare de tip depresor. Existenta lor este dovedita experimental prin experienta lui Goltz, cand stimularea unei anse intestinale determina reflex oprirea cordului. Pentru a preveni acest reflex in cursul interventiilor chirurgicale pe abdomen, in preanestezie se injecteaza atropina care blocheaza receptorii M pe care actioneaza acetilcolina. Fara receptori disponibili, stimularea parasimpaticului nu mai are efecte deprimante asupra cordului.
Caile aferente ale receptorilor cardiovasculari
Baroreceptorii din crosa aortica sunt legati prin fibre ale nervului vag (nervul Cyon-Ludwig) de centrul cardioinhibitor si vasodepresor. Baroreceptorii din sinusul carotidian sunt legati prin fibre ale glosofaringianului (nervul Hering) de centrul cardioinhibitor si vasodepresor din bulb.
Nervul vag si glosofaringian se mai numesc si nervi tampon. Stimularea lor electrica produce bradicardie, vasodilatatie si hipotensiune arteriala, iar sectionarea lor produce cresterea tensiunii arteriale pana la valori de 300/200 mmHg -"hipertensiune neurogena sau de defrenare".
Caile de legatura ale celorlalti receptori cardiovasculari cu centrii de reglare sunt realizate prin ramuri ale nervului vag.
De la receptorii din vase si viscere la centrii vasculari din maduva toracala T1-T4, prin n. aferenti simpatici.
Chemoreceptorii ventriculari sensibili la anoxie si acidoza au aferente apartinand sistemului simpatic, care ajung la centrii medulari vasoconstrictori sau cardioacceleratori.
Centrii nervosi de reglare cardiovasculara sunt:
Centrii de integrare: scoarta, hipotalamus, formatiunea reticulate (FR);
Centrii de comanda: bulb si maduva.
▪ Centrii de integrare
In neocortex exista centrii care-si exercita infuenta activatoare sau inhibitorie, cu sau fara participarea hipotalamusului ca statie intermediara. De exemplu: stimularea ariei premotorii (girus sigmoid anterior, regiunea fronto-orbitara) produce reactii de tip vasoconstrictor si presor; excitarea cortexului motor produce reactii vasodilatatoare la musculatura scheletica in efortul fizic.
Reactiile vasomotorii corticale se produc fie direct pe calea cortico-spinala, fie indirect via hipotalamus, bulb. Impulsurile sunt transmise prin n. simpatici colinergici vasodilatatori la musculatura activa. Zona este vecina cu zona motorie, controlul cortical al reactiilor motorii si circulatorii fiind sinergic.
Sistemul limbic - in nucleul amigdalian si hipocamp exista zone care care coordoneaza reactii vasomotorii importante.
Hipotalamusul prezinta zone cu structuri neuronale vasoconstrictoare (H. posterior si mijlociu) via centrul vasomotor bulbar si centrii cardiaci si zone cu structuri neuronale vasodilatatoare (H anterior).
La acest nivel exista mecanisme de reglare a secretiei de catecolamine din medulosuprarenala si sediul reglarii homeostaziei termice.
Formatiunea reticulara bulbopontina
stimularea electrica a zonei laterale si anterioare produce reactii de vasoconstrictie cu cresterea TA; stimularea zonei mediale produce efect vasodilatator.
▪ Centrii de comanda:
In bulb se gaseste: centrul cardioinhibitor (nucleul ambigu si nucleul dorsal al vagului) si centrul vasomotor cu o arie presoare si una depresoare. Primesc aferente din toate zonele corpului.
Stimularea electrica a centrul cardioinhibitor produce bradicardie si scaderea debitului cardiac prin eferentele vagale.
Centrul vasomotor, prin cele doua zone ale sale (presorie si depresorie), moduleaza activitatea cordului si vaselor, actionand numai asupra centrilor vasoconstrictori medulari, carora le stimuleaza sau le inhiba activitatea.
Centrii medulari:
- centrii vasomotori T1-T4;
- centrii cardioacceleratori (n. cardiaci superiori, mijlocii si inferiori) Centrii cardioacceleratori sunt localizati in principal in segmentele T1 si T2, dar la inervatia inimii mai participa si fibre simpatice cu originea in segmentele T3-T6;
- centrii vasoconstrictori T1-L4;
- centrii parasimpatici vasodilatatori S1-S4.
Sectionarea maduvii spinarii in regiunea cervicala produce o scadere rapida a presiunii arteriale si vasodilatatie periferica, fenomen numit "socul spinal". Daca se pastreaza nervul frenic si eferenta simpatica preganglionara, situatia revine la normal in cateva zile.
Caile eferente caracteristici generale:
pornesc de la centrii nervosi;
- sunt parasimpatice sau simpatice ;
- sunt amielinice ;
- au pe traseu un ggl unde se face sinapsa fibrelor preganglionare cu cele postganglionare ;
- deci calea eferenta are doi neuroni si o sinapsa ;
- fibrele postggl parasimpatice=scurte;
- fibrele postggl simpatice= lungi;
- fibrele vegetative formeaza plexuri periviscerale, periarteriale, intraviscerale.
- eferenta: pe cord sau pe vase
Eferenta parasimpatica. Eferenta parasimpatica are 2 neuroni: protoneuronul localizat in centrul cardioinhibitor si deutoneuronul localizat in musculatura atriala, in vecinatatea nodulului sinoatrial (NSA) si a celui atrioventricular (NAV). Inervatia vagala a musculaturii ventriculare si a fasciculului His este foarte slab reprezentata. Miocardul ventricular este sarac in fibre vagale, relativ independent de stimularea vagala.
Vagul drept inerveaza mai ales zona NSA, excitatia sa producand bradicardie sau chiar oprirea inimii pentru cateva secunde in diastola, in timp ce vagul stang influenteaza mai ales conducerea atrioventriculara (vagul stang se termina la nodulul atrioventricular).
Inervatia parasimpatica a ventriculilor se realizeaza pe calea axonilor postganglionari colinergici, ce au corpul celular in atrii.
Excitatia vagala, in afara de bradicardie si intarzierea conducerii atrioventriculare mai produce o usoara scadere a fortei de contractie prin efecte inotrop negative (scaderea presiunii maxime ventriculare si scaderea presiunii in diastola).
Excitarea vagala prelungita produce oprirea inimii 4-10 s. Chiar daca stimularea continua inima isi reia activitatea = fenomen de scapare de sub influenta vagala vagus escape. Ipoteze:
- stimularea vagului à inhibitia centrului primar de automatism + oprire cord;
- escape prin intrarea in activitate a centrilor secundari de automatism sau: oprirea inimii in diastola à mec. Frank Starling, sau: anoxia centrilor CV à reluare prin mecanism central;
- cea mai plauzibila: epuizarea mediatorului chimic din fibrele postggl (Ach);
- fenomenul de scapare are uneori aspect de rebound, excitatie postinhibitoare (forta de contractie si frecventa mai mare).
Denervarea parasimpatica a cordului, realizata prin metode chirurgicale sau farmacologice (cu atropina) produce tahicardie (frecventa cardiaca creste de la 70 la 150/min.).
Efectele parasimpatice se realizeaza prin eliberarea de acetilcolina la nivelul terminatiunilor nervoase. Acetilcolina actioneaza asupra receptorilor muscarinici - M - membranari deprimand toate proprietatile miocardului.
Acetilcolina determina eliberare de GMPc, urmata de scaderea activitatii adenilatciclazei si a AMPc. Activitatea inhibitorie la nivelul miocardului se datoreste hiperpolarizarii indusa de cresterea permeabilitatii pentru K+ a membranei celulare.
Tonusul vagal predomina celui simpatic, iar la sportivi este mai accentuat decat la nesportivi !!!
Eferenta simpatica. Caile simpatice au originea in cordoanele intermediolaterale ale primelor 5-6 segmente toracice medulare. Sinapsa cu neuronii postganglionari se face la nivelul ganglionilor cervico-dorsali, de unde pleaca fibrele postganglioanare sub forma nervilor cardiaci (superior, mijlociu si inferior), care inerveaza sistemul excitoconductor si fibrele contractile.
Fibrele simpatice drepte se repartizeaza mai ales in tesutul nodal, afectand in special frecventa, in timp ce cele stangi se distribuie cu precadere in micardul contractil, amplificand activitatea.
Denervarea simpatica, realizata pe un cord in prealabil denervat vagal, determina scaderea frecventei cardiace (la om de la 150 la 100/min.).
Stimularea simpatica elibereaza NA si A care actioneaza pe receptorii beta-adrenergici miocardici si coronarieni stimuland toate proprietatile miocardului.
Actiune la nivel transmembranar:
- stimuleaza adenilatciclaza, productia de AMPc si patrunderea acestuia in celula;
- scade permeabilitatea ptr K+;
- creste permeabilitatea pentru Na+ si Ca+, producand depolarizare;
- scade pragul de excitabilitate;
- scade durata potent de pacemaker;
- reduce inclinarea pantei de prepotential.
Caile eferente pe vase:
Nervii vasoconstrictori
- cap, gat à origine: maduva T1-T4 si C8, primul neuron: coarnele laterale, al doilea neuron: ggl cervical superior si mijlociu. Fibrele postggl à plexuri periarteriolare si perivenoase;
- extremitati à T4-T10 si T11-L3;
- viscerele abdominale: T6-T12 si L1-L3 à sinapsa: ggl celiac à fibrele postggl amielinice se distribuie vaselor viscerelor abdominale.
Simulul simpatic se distribuie predominant arteriolelor - bogate in fibre m. netede !!!
Nervii vasodilatatori
- reprezentati de parasimpaticul cranian (fibre ale n. cranieni VII, IX, X) si parasimpaticul sacrat (S1-S3) ;
- sunt fibre simpatice vasodilatatoare colinergice care la terminatiile postggl elibereaza mediatorul chimic acetilcolina àvasodilatatie.
Mediatorii SNV si influenta lor asupra cordului si vaselor
Functie de locul de eliberare a mediatorului:
▪ Sistem colinergic -Acetilcholina
▪ Sistem adrenergic - adrenalina sau epinefrina si noradrenalina sau norepinefrina
Acetilcolina - mediatorul sistemului parasimpatic, produce efecte deprimante asupra tuturor proprietatilor cordului (efecte cronotrop, dromotrop, inotrop, batmotrop si tonotrop negative), actionand asupra receptorilor muscarinici.
Efectele cronotrop negative ale acetilcolinei se explica prin cresterea permeabilitatii membranei celulelor "P" (pace-maker) pentru K+, ceea ce determina hiperpolarizare prin iesirea K+ din celula si aparitia unui eflux de K+ in cursul depolarizarii lente diastolice. Hiperpolarizarea determina indepartarea potentialului diastolic maximal de nivelul critic de declansare si scaderea consecutiva a frecventei de descarcare a impulsurilor. Efluxul de K+ in timpul depolarizarii lente diastolice determina contrabalansarea influxului lent, depolarizant de Na+ si Ca2+, care tinde sa aduca potentialul de membrana la nivelul critic de declansare, producand astfel scaderea vitezei de depolarizare lenta diastolica si deci a frecventei de descarcare a impulsurilor.
Efectele inotrop negative ale acetilcolinei se explica prin scurtarea duratei potentialului de actiune al fibrelor miocardice, realizata pe seama platoului, ceea ce reduce influxul de Ca2+ ce precede sistola. S-a determinat prin studii efectuate cu microelectrozi pe un singur canal de Ca2+ (tehnica de patch-clamp), ca acetilcolina blocheaza canalele de Ca2+, cu atat mai puternic cu cat acestea erau mai intens activate de stimularea adrenergica.
Catecolaminele - adrenalina si noradrenalina eliberate la nivelul terminatiilor postsinaptice simpatice ca si cele produse de medulosuprarenala stimuleaza proprietatile fundamentale ale miocardului, actionand asupra receptorilor miocardici alfa si beta adrenergici.
Adrenalina actioneaza pe: alfa si beta:
- prin actiunea asupra beta 1 de la nivelul cordului stimuleaza toate proprietatile miocardului;
- actiunea asupra beta 2: vasodilatatie musculara, cerebrala, coronariana, vasoconstrictie splanhnica, relaxare musculatura neteda bronsica, intestinala, uterina, midriaza, piloerectie, stimuleaza eliberarea de CRF, ACTH, activare corticala.
O cantitate mare de epinefrina provenita din meduosuprarenala are efecte vasoconstrictoare prin actiunea asupra receptorilor 1-adrenergici, in timp ce o concentratie mica exercita efecte vasodilatatoare prin intermediul receptorilor 2-adrenergici din miocard, muschiul scheletic, creier si ficat. In fapt, efectul diferit al epinefrinei se datoreaza tipului de receptor pe care actioneaza aceasta. Receptorii de tip 1 sunt dominanti in vasele din piele si rinichi in timp ce 2 predomina in organele amintite mai sus.
Noradrenalina: actiune pe: alfa1 si alfa 2; efect: vasoconstrictie generalizata
Efectele cronotrop pozitive ale adrenalinei se explica prin cresterea permeabilitatii membranei celulelor pace-maker pentru Na+ si Ca 2+, ceea ce accelereaza panta depolarizarii lente diastolice.
Efectele dromotrop pozive se produc prin cresterea vitezei de conducere prin NAV si fasciculul His, datorita cresterii permeabilitatii celulelor acestor structuri pentru Na+ si Ca2+ .
Efectele inotrop pozitive se realizeaza prin stimularea receptorilor b miocardici, ceea ce produce activarea in interiorul acestor celule a adenilatciclazei, proces care necesita energie furnizata de GPT. Adenilatciclaza determina transformarea ATP in AMP ciclic, care actioneaza asupra proteinkinazei ciclic AMP-dependente, disociind-o in 2 subunitati catalitice si 2 subunitati regulatoare. Una din subunitatile catalitice, proteinkinaza A, fosforileaza proteinele canalului de Ca2+, determinand cresterea influxului de Ca2+ in sistola de 3-4 ori, stimuland astfel contractilitatea.
Proteinkinaza A mai determina fosforilarea unei proteine, fosfolambanul, care activeaza pompa de Ca2+ din reticulul sarcoplasmic, determinand cresterea vitezei de recaptare a Ca2+ in reticul, in perioada care precede diastola ventriculara. Rezulta scurtarea fazei de relaxare, dar si cresterea cantitatii de Ca2+ din reticul disponibila pentru urmatoarele sistole, ceea ce favorizeaza stimularea inotropismului.
Proteinkinaza c-AMP dependenta fosforileaza si fractiunea I a troponinei, reducand afinitatea troponinei C pentru Ca2+, ceea ce scurteaza perioada de relaxare a miocardului, fenomen esential pentru prevenirea tetanizarii cordului la frecvente crescute de contractie. AMP ciclic impreuna cu complexul Ca2+-modulina activeaza fosforilkinaza, care produce degradarea glicogenului, furnizand astfel energia necesara pentru viitoarele cicluri de contractie-relaxare. AMP ciclic este degradat in celula sub actiunea fosfodiesterazei, enzima care poate fi blocata de substante din clasa metilxantinelor (cafeina, teofilina). Astfel, metilxantinele determina, ca si stimularea b-adrenergica, cresterea concentratiei AMPc intracelular.
Stimularea receptorilor b determina si vasodilatatie coronariana, creand conditii bune de de lucru ale miocardului.
2. Reglarea umorala a cordului si vaselor - caracteristici:
- asigura o reglare de durata
- completeaza si prelungeste in timp efectele reglarii nervoase
- se realizeaza prin: substante vasodilatatoare si vasoconstrictoare (mediatori ai SNV, hormoni locali, polipeptide vasoactive, cataboliti acizi, gaze sanguine, ioni).
Substante vasodilatatoare
▪ Acetilcolina - vezi efectele mai sus
▪ Histamina - Se formeaza prin decarboxilarea enzimatica a histidinei si este stocata sub forma de complexe cu heparina in granulele mastocitelor si a leucocitelor bazofile. Actioneaza pe trei tipuri de receptori: H1, H2, H3.
Rol: vasodilatatie si creste permeabilitatea vasculara, efect exprimat mai ales in timpul infectiilor sau a reactiilor de coagulare.
▪ EDRF (Factor Endotelial de Relaxare, NO) Monoxidul nitric actioneaza ca agent vasodilatator fiind eliberat de celulele endoteliale ca urmare a legarii acetilcolinei, ATP-ului, endotelinei sau histaminei de celulele endoteliale. Dupa eliberare, NO difuzeaza si produce relaxarea fibrelor musculare netede din vecinatate si implicit vasodilatatie.
▪ Bradikinina si kalidina sunt de asemenea agenti vasodilatatori ei provenind din kininogeni in urma actiunii enzimei kalicreina. Produce vasodilatatie arteriolara, creste permeabilitatea capilara, inhiba secretiile digestive etc..
▪ FAN (Factorul Atrial Natriuretic)- hormon secretat de miocitele atriale. Produce vasodilatatie si cresterea permeabilitatii capilare.
▪ Adenozina - sintetizata din AMP are rol de mediator chimic actionand pe receptorii A1 si A2. Stimularea receptorilor A2 stimuleaza adenilat-ciclaza care va produce mai mult cAMP care va determina relaxarea fibrelor musculare netede din peretii vaselor. Alte efecte: vasodilatatie coronariana, bradicardie, efect batmotrop si dromotrop negativ, creste permeabilitatea vasculara.
▪ Prostaglandine (Prostaciclina, E1, E2) - se formeaza din degradarea acizilor grasi nesaturati, in special acidul arahidonic sub actiunea ciclooxigenazelor. Se gasesc in majoritatea tesuturilor unde actioneaza ca hormoni locali. Prostacilina sau PGI2 este sintetizata de celulele endoteliale si are atat actiune vasodilatattoare, cat si de inhibare a agregarii plachetare.
▪ Catabolitii acizi si CO2 - au rol mai ales in reglarea locala a circulatiei. Acumularea de CO2, de ioni de hidrogen care scad pH, scaderea concentratiei de O2 (hipoxia) determina vasodilatatie prin actiunea directa asupra celulelor musculare netede mai ales de la nivelul arteriolelor precapilare. Deficitul de O2 determina in general dilatatia vaselor, cu exceptia celor de la nivel pulmonar unde scaderea presiunii partiale a O2 determina vasoconstrictie hipoxica. Efectele vasodilatatoare ale celor 3 factori se manifesta numai in periferie, local. Daca scaderea concentratiei de O2 sau acumularea de CO2 si H+ este sesizata de chemoreceptorii din zonele reflexogene sau de cei bulbari atunci se produce vasoconstrictie periferica generalizata. In cazul hipoxiei determinata de ischemia cerebrala acest raspuns vasoconstrictor periferic are rolul de a evita accidentele cerebrale deoarece efectul vasoconstrictiei este cresterea presiunii arteriale.
Subsatante vasoconstrictoare
Noradrenalina;
Adrenalina;
Serotonina -creste permeabilitatea vasculara, potenteaza raspunsul chemotactic al leucocitelor si stimuleaza formarea de colagen. Prin stimularea terminatiilor nervoase libere declanseaza durerea locala (factor algogen).
▪ Angiotensina II - secventa de sinteza: hipoxia renala determina eliberarea de renina. Acesta transforma angiotensinogenul (o proteina plasmatica de origine hepatica) in angiotensina I. Sub actiunea enzimei de conversie angiotensina I este transformata in angiotensina II, una dintre cele mai puternice substante vasoconstrictoare naturale. Din pacate, angitensinogenul se sintetizeaza si la nivelul cordului, inclusiv in circulatia coronara. Tot la acest nivel s-au descoperit receptori pentru angiotensina precum si prezenta enzimei de conversie, ceea ce creste riscul de boala coronariana.
ADH (vasopresina) - prin receptorii V1 scade cantitatea de cAMP (se blocheaza stimularea adenilat-ciclazei de catre adrenalina);
Endotelina;
Prostaglandinele F1, F2, Tromboxanul A2.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 13124
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved