| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
RASPUNSUL IMUN (IMUNOGENEZA)
Raspunsul imun sau imunogeneza este reactia organismului prin care acesta produce efectorii imuni consecutivi unui stimul antigenic. Prima conditie a dezvoltarii unui raspuns imun este recunoasterea non-selfului. Ca si in cazul imunitatii naturale, organsimul actioneaza in sensul distrugerii, al indepartarii sau anihilarii efectelor nocive ale non-selfului. In timp ce mecansimele imunitatii naturale, actioneaza insa in mod egal, nediferentiat impotriva non-selfului, indiferent de compozitia lui chimica si indiferent daca acesta are sau nu proprietati antigenice, raspunsul imun are loc numai fata de structuri non-self dotate cu atributele antigenitatii si se caracterizeaza printr-un inalt grad de specificitate.
In cazul raspunsului imun, organismul elaboreaza efectori imuni, reprezentati de structuri de neoformatie, inexistente anterior patrunderii antigenului. Acestea pot fi, ca nivel de organizare, molecule sau celule.
Efectorii imuni cu nivel de organizare molecular, se numesc anticorpi, denumirea subliniind complementaritatea lor fata de antigen sau imunoglobuline, termen mai modern care precizeaza compozitia chimica corelata cu interventia lor in procesele imune.
Efectorii imuni cu nivel de organizare celulara poarta denumiri diferite, in functie de natura celulelor efectoare, de modul lor de formare si de actiune.
Starea de imunitate instalata pe baza raspunsului imun poate fi considerata ca fiind de tip umoral sau celular, in functie de predominanta uneia din cele doua categorii de efectori imuni; impartirea aceasta, este arbitrara intrucat si efectorii imunitatii umorale sunt de fapt tot rezultatul unor activitati celulare.
Trasatura esentiala a raspunsului imun consta in faptul ca atat in recunosterea non-selfului cat si in procesul de elaborare a efectorilor imuni organismul reuseste sa diferentieze detalii fizico-chimice de extrema finete ale antigenului si sa determine negativul acestuia sub forma unor complementaritati de o extrem de riguroasa specificitate. Capacitatea de discernere si diferentiere a detaliului de catre organism este elocvent demonstrata de exemplul clasic in care, in elaborarea anticorpilor specifici unor antigene artificiale preparate prin cuplarea unei proteine cu acidul tartric, gradul de specificitate al anticorpilor merge pana la sesizarea tipului de rotatie al luminii polarizate. Astfel, anticorpii a caror formare este determinata de antigenul ce contine acid tartric dextrogir nu reactioneaza decat cu acest antigen si sunt incapabili sa reactioneze cu antigenul in care gruparea determinanta este reprezentata de acidul tartric levogir.
Producerea efectorilor imuni este determinata de acea insusire a unor sisteme morfofunctionale ale organismului numita imunocompetenta.
Intr-un sens mai larg, imunocompetenta este atribuita tuturor organelor, tesuturilor, celulelor care contribuie intr-un fel sau altul la realizarea raspunsului imun.
Intr-un sens mai ingust, imunocompetenta este recunoscuta numai elementelor morfofunctionale care produc prin biosinteza sau din care rezulta prin transformare blastica efectori imuni.
Intr-o conceptie mai moderna, inspirata din lucrarile imunologului VICTOR GHETIE, imunocompetenta ar putea fi definita 'ca insusirea unor celule din organism de a recunoaste cu ajutorul receptorilor imunologici de pe suprafata lor semnalele antigenice, de a le traduce si de a le amplifica intracelular, determinand celula sa produca efectori imuni, sau sa indeplineasca ea insasi acest rol'.
Treapta filogenetica la nivelul careia apare imunocompetenta o reprezinta vertebratele. Pestii sunt prin urmare vietuitoarele cele mai evoluate, capabile sa dezvolte un raspuns imun. Suind pe scara filogenetica se constata o stransa corelatie intre superioritatea treptei si complexitatea raspunsului imun.
1. EFECTORII UMORALI AI RASPUNSULUI IMUN (ANTICORPII)
Raspunsul imun umoral se realizeaza prioritar fata de urmatoarele antigene: eritrocite heterologe, bacterii, virusuri, proteine plasmatice heterologe, paraziti care determina infestatii cu evolutie acuta, etc. Veriga efectoare a raspunsului imun umoral o reprezinta niste proteine specifice numite anticorpi (Atc.) sau imunoglobuline (Ig) sintetizate in urma stimularii cu un anumit antigen, capabile sa se uneasca cu antigenul inductor formand asa numitele 'reactii antigen - anticorp'.
Atc. sunt molecule complexe cu 2 poli functionali.
Un pol serveste pentru 'recunoasterea antigenelor'. Este o zona cu structura moleculara foarte variabila, care este adaptata pentru recunoasterea si cuplarea cu enorm de multe structuri antigenice din mediu (se admite ca numarul maxim de structuri elementare recunoscute de catre anticorpi este de ordinul 1017).
Celalalt pol denumit 'efector' are o structura constanta. El serveste pentru amorsarea activarii complementului prin calea clasica si pentru atasarea (imunoaderenta) la macrofage. Din punct de vedere biologic, Atc. sunt molecule adaptative bifunctionale. Ei servesc ca piese de legatura care mijlocesc atasarea Atg. pe fagocite. Atc. nu distrug antigenele, dar convertesc diversitatea materiala a antigenelor intr-un semnal unic si accesibil mecanismului ancestral de aparare care este fagocitoza.
1.1. Structura Ig.
Ig sunt formate din combinatii de lanturi polipeptidice cu lungimi diferite: unele lungi, denumite lanturi grele sau H (de la 'Heavy'), celelalte scurte - lanturi usoare sau L (de la 'Light'). Exista cinci tipuri de lanturi H (gama, alfa, miu, delta si epsilon) si doua tipuri de lanturi L (kapa si lambda). Lanturile H definesc clasa imunoglobulinelor (ex. gama pentru IgG, alfa pentru IgA, miu pentru IgM, delta pentru IgD, epsilon pentru IgE.). Genele care comanda productia de lanturi H si L sunt plasate pe cromozomi diferiti (cromozomul 14 pentru lanturile H, cromozomul 2 pentru lanturile K si cromozomul 22 pentru lanturile lambda). Ig sunt elaborate de LfB. O celula produce un singur tip de lant usor (kapa sau lambda). Lanturile grele si cele usoare se formeaza pe poliribozomi diferiti, dupa care se asambleaza 2 cate 2 pentru a forma molecula de Ig.
1.2. Clasele de Ig.
IgG Structura acestei Ig este considerata modelul tip pentru toate clasele de Ig.: 2 perechi de lanturi formate fiecare din ele din cate un lant H plus un lant L. Cele patru lanturi sunt legate intre ele prin punti disulfidice. Configuratia generala a moleculei schematizata dupa imaginea observata la microscopul electronic, aminteste forma literei Y, in care cele doua brate reprezinta extremitatile aminoterminale, iar coada, extremitatile COOH- ale lanturilor.
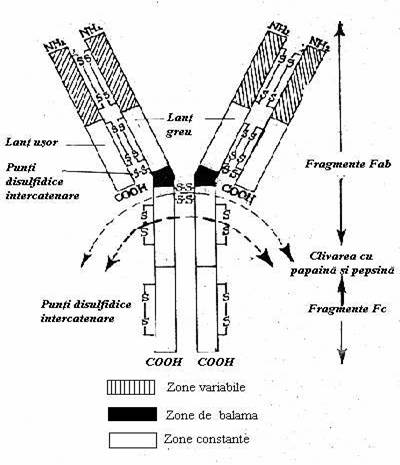
Fig. 17 Schema unei molecule de imunoglobulina G
Dupa compozitia in aminoacizi, lanturile H si L sunt formate din doua domenii: variabil (V), la extremitatea aminoterminala si constant (C) la extremitatea carboxilica. Secventa de aminoacizi din compunerea domeniilor V difera de la o molecula la alta. Diferentele sunt maxime in trei puncte ale secventei - zonele 'hipervariabile' care formeaza situsul combinativ cu antigenul. Domeniile C sunt cele care caracterizeaza lanturile gama, kapa sau lambda. Exista patru variante de lanturi gama (gama 1 gama 4) care definesc patru subclase de IgG (IgG1.IgG4). Papaina ataca moleculele de IgG in amonte de legatura -S-S- care uneste cele doua lanturi H. Rezulta doua fragmente Fab (ab = 'antigen binding') care contine secvente combinative cu Atg. si un fragment Fc (c = 'cristalizabile') care constituie elementul efector al moleculei (fixarea complementului si fixarea pe celule care poseda receptori specifici pentru Fc). (Fig. 17).
Locul activitatii proteolitice a pepsinei este plasat in aval de puntea disulfidica dintre lanturile H. Rezulta un rest F (ab)2 format din cele doua Fab - uri unite prin puntea -S-S- si care se comporta ca un anticorp, plus 2 resturi ale CH care sunt degradate in peptide mici. IgG pot strabate barierele endoteliale. Ele difuzeaza in spatiile extravasculare unde participa activ la reactia de aparare antibacteriana. Dupa atasarea pe corpii microbieni, IgG mijlocesc prin segmentul efector Fc, imunoaderenta si, uneori, activarea complementului. Prin abilitatea de a traversa placenta ele protejeaza nou-nascutul (al carui sistem imun este incapabil de reactii eficiente) impotriva infestarii cu microbi. (Tabelele nr. 3 si 4)
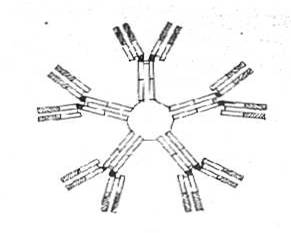
IgM au dimensiuni mult mai mari ('macroglobuline') rezultate din asocierea a 5 unitati
monomerice compuse si ele din imperecherile de lanturi H (aici lanturi miu) cu
lanturi L (kappa sau lambda). Exista doua feluri de lanturi miu (miu 1 si miu 2). Cele 5 unitati
monomerice se unesc intre ele prin extremitatile distale ale segmentelor Fc
formand imaginea unei stele cu cinci brate.
Fig. 18 Structura imunoglobulinei M
In consecinta molecula pentamerica contine 10 Fab-uri (10 situsuri combinative cu antigenul. (Fig.18).
Sinteza IgM precede pe aceea a IgG in raspunsul imun primar. Din cauza dimensiunilor lor foarte mari, IgM nu poate parasi usor spatiul intravascular, unde joaca un rol important in combaterea bacteriilor. Prin multiplele lor valente combinative cu antigenul, ele pot lega simultan, mai multi corpi bacterieni (fenomen care este denumit aglutinare), iar prin abilitatea lor marcata de activare a complementului (o molecula de IgM fixata pe un Atg. declanseaza calea clasica) produc liza acestora.
IgA se prezinta sub doua forme: IgA serica si IgA secretorie. Structura de baza este o combinatie de 2 lanturi alfa cu doua lanturi L (kappa si lambda).
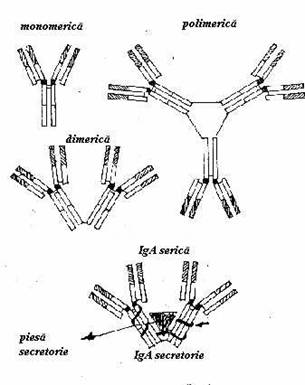
Fig. 19 Structura imunoglobulinei A
Exista doua subclase de IgA (alfa 1 si alfa 2). IgA serica poate fi monomerica, dimerica sau polimerica. Rolul sau nu este bine cunoscut. IgA secretorie este prezenta in concentratie ridicata in saliva, lapte, lacrimi, sputa sau secretii digestive. Este formata din doua unitati monomerice unite intre ele printr-o 'piesa secretorie' si un mic polipeptid (lantul J). (Fig.19)
IgA secretorie este produsa local de elemente limfoide specializate din submucoase, sub forma de dimeri stabilizati prin lanturi J.
Acestia se unesc cu un receptor expus pe membrana celulelor epiteliale cu care formeaza un complex care traverseaza aceste celule catre suprafata lor externa sau luminala. La acest nivel molecula receptorului se scindeaza, iar fragmentul care ramane legat cu dimerul de IgA formeaza 'piesa secretorie'. Aceasta din urma ar avea rolul de a apara de proteoliza IgA expusa pe suprafete. IgA secretorie asigura protectia antibacteriana la nivelul suprafetelor care vin in contact cu mediu extern. Ea inhiba aderenta microorganismelor la mucoase, prevenind in acest mod strabaterea barierelor externe de catre agentii microbieni.
IgD este la fel ca IgG o structura monomerica cu formula delta2 + L2 (kapa sau lambda). Concentratia serica a IgD este foarte redusa (0,025 g/l). La fel ca si IgM, IgD sunt frecvent exprimate pe suprafata limfocitelor B unde opereaza ca receptori specifici pentru Atg.
IgE sunt produse in titruri mari in cursul reactiilor alergice, cand se fixeaza pe receptorii Fc de pe memebrana granulocitelor bazofile si a mastocitelor declansand eliberarea de histamina si de alte amine vasoactive. Au fost denumite si 'reagine'. Sunt compuse din lanturi epsilon si L. Urme de IgE exista si in serul normal. Rolul fiziologic al IgE ar fi cel de protectia suprafetelor mucoaselor externe. Microbii care depasesc apararea locala mediata de IgA si se combina cu IgE, induc prin intermediul unor mecanisme mediate de acestea, o recrutare locala de efectori solubili (de ex. complement si IgG si de celule in special de fagocite).
Ig sunt produse de LfB sub doua forme (fig.20):
1. Ig exprimate pe suprafata membranei LfB: moleculele respective sunt incastrate in membrana Lf printr-o 'piesa de legatura' (secventa de aminoacizi de la nivelul carboxiterminal al moleculei). Extremitatea aminoterminala (care contine situsul combinativ) proemina in afara membranei. Ig de suprafata (sIg) joaca rolul de receptor pentru Atg. al LfB. Interactiunea specifica sIg - Atg. produce un semnal care activeaza LfB.
2. Ig secretate de catre LfB activate si transformate in plasmocite se gasesc ca molecule libere in plasma, in secretii si lichidele extravasculare unde joaca rol de anticorpi care se cupleaza cu antigenul recunoscut in prealabil de sIg. Aceasta dubla recunoastere si interactiune cu Atg. este posibila deoarece sIg. si Atc., ca produsi ai aceleiasi celule sau ai unei clone de celule, au situsul combinativ cu structura moleculara identica. Spre deosebire de sIg moleculele de Atc. nu au piesa de legatura. Extremitatea lor carboxiterminala poseda un situs de combinare cu componenta C1q a complementului. C1q se ataseaza pe Atc. dupa cuplarea acestora cu Atg. Atasarea C1q la Atc. declanseaza secventele activarii C' prin calea clasica.

a b
Fig. 21 Modul de eliminare a Atg prin fagocitoza imun
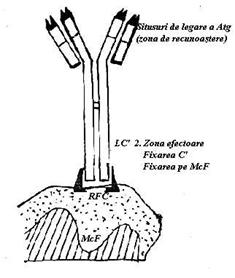 |
Fixarea si activarea complementului impreuna cu atasarea si fagocitarea complexelor Atg - Atc reprezinta cele doua mecanisme prin care se realizeaza eliminarea Atg. (Fig.21).
Ig reprezinta o situatie unica in biologie pentru ca sunt in acelasi timp si anticorpi si antigene (prin natura lor proteica se comporta ca imunogene care provoaca aparitia unor Atc anti - Ig). Acestia recunosc diversi epitopi ai moleculelor de lanturi H si L.
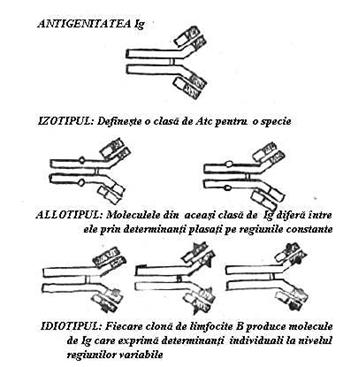
Exista
trei feluri de epitopi (determinanti antigenici) distribuiti in anumite zone
precise ale moleculei (Fig.22).
Fig. 22 Antigenitatea imunoglobulinelor (Ig)
1. Epitopi care determina specificitatea de specie (izotipica). Clasele de Ig sunt identice din punct de vedere antigenic pentru toti indivizii speciei sgr. isos (egal)+tupos (forma, amprenta)t. Izotipurile sunt determinate de lanturile H. Antigenicitatea izotipica poate fi exploatata in practica pentru prepararea de reactivi. Ig umane injectate unui iepure se comporta ca antigene. Animalul dezvolta o reactie imuna in urma careia se va produce anticorpii anti-Ig umane. Serul animalului imunizat a primit numele de 'antiser' deoarece reactioneaza in vitro (prin anticorpii sai anti-Ig) cu Ig din serul uman.
Cu ajutorul unui astfel de antiser se poate evidentia prezenta Ig umane in diverse umori sau fixate pe suprafata unor celule. Pe acest principiu functioneaza testul Coombs direct (TCD) care evidentiaza Atc. fixati pe Atg eritrocitare. In practica medicala TCD serveste pentru identificarea unor imunizari posttransfuzionale sau pentru diagnosticul anemiilor hemolitice autoimune si al bolii hemolitice a nou-nascutului.
2. Epitopii specifici pentru indivizii unei specii (allotipuri) (gr. allos = altul + tupos). Acesti epitopi sunt exprimati pe regiunile constante ale lanturilor gama (sistemul A2m) sau kapa (sistemul Km).
Allotipurile sunt comandate de gene somatice si se transmit dupa legile mendeliene. Datorita expresiei acestor allotipuri, Ig difera din punct de vedere antigenic de la un individ la altul: administrate la alti indivizi ai speciei, Ig unei clase (izotip) pot genera in anumite conditii sinteza de anticorpi anti-allotip. Foarte rar, incompatibilitatea dintre allotipuri pot genera reactii post transfuzionale.
3. Epitopi care sunt specifici numai pentru moleculele de Ig, elaborate de o clona celulara (idiotipuri): un individ va elabora familii de molecule de Ig care exprima fiecare alt idiotip s(gr.idios (particular) + tupos (forma)t
JACQUES OUDIN (1963) a demonstrat existenta idiotipurilor intr-un experiment pe iepuri cu identitate allotipica. El a administrat unui iepure un Atg. (X), apoi a separat Atc. anti-X si i-a administrat la alt iepure. In mod neasteptat, cel de-al doilea animal a dezvoltat Atc. anti-anti-X, sugerand ca moleculele anti-X poseda structuri antigenice particulare si individuale, altele decat specificitatea allotipica. Determinantii idiotipici sunt plasati la nivelul regiunilor hipervariabile ale lanturilor H si L. S-a demonstrat ulterior ca un individ normal ar dispune de mai multe sute de mii de idiotipuri diferite. distribuite pe Ig serice si pe receptorii pentru antigen ai limfocitelor B si T. Raspunsul la antigenul X nu s-ar limita numai la productia de Atc. specifici, deoarece acestia prin determinantii lor idiotipici (care se comporta ca antigene) ar declansa si productia unor anticorpi anti-idiotip (anti-Id1). Acestia la randul lor vor genera molecule de anti-anti-idiotip (anti -Id2) care in continuare vor duce la productia unor anti -Id3, s.a.m.d. Daca admitem foarte schematic ca situsurile combinative ale unui Atc. ('paratopii') reproduc 'in negativ' configuratia spatiala a Atg. si ca aceleasi situsuri combinative contin si idiotipurile generatoare de anti-Id, putem concepe doua aspecte caracteristice ale acestei reactii in lant:
a) anti-Id2 pot prezenta un grad mare de identitate cu Atc. initiali anti-X si deci posibilitatea de recunoastere a Atg;
b) configuratia paratopurilor ce intra in compunerea anti-Id1 ar putea fi extrem de asemanatoare cu Atg. insusi. Atc. anti-Id1 s-ar comporta deci ca un fel de 'imagine interna' a Atg., ceea ce inseamna ca in interiorul sistemului imunitar sunt reproduse elemenete ale lumii exteriroare organismului. Aceasta retea de interactiuni, in care mai sunt implicate si celulele limfoide B si T (prin idiotipurile exprimate de receptorii lor pentru Atg.) creaza un sistem de reglare foarte flexibil (teoria retelei idiotipurilor conceputa in 1974 de NIELS JERNE).
1.3. Biosinteza si catabolismul imunoglobulinelor
Biosinteza imunoglobulinelor se realizeaza in interiorul plasmocitului, prin formarea independenta a lanturilor H si L la nivelul ribozomilor, fiecare lant fiind codificat de o gena separata in interiorul celulei secretoare. Poliribozomii cu 7-8 unitati ribozomale sintetizeaza lanturile. Asamblarea lanturilor polipeptidice se face la nivelul reticulului endoplasmatic plasmocitar.
Procesul de sinteza a imunoglobulinelor se supune legii cibernetice a conexiunii inverse (Feedback) in sensul ca la un prag inalt de concetratie biosinteza este inhibata, iar in cazul scaderii nivelului de imunoglobuline in umori, procesele de sinteza sunt accelerate.
Degradarea moleculelor de imunoglobuline sau catabolizarea lor, exprimata ca 'timp de injumatatire' (T 1/2) este un proces fiziologic care permite inlocuirea permanenta a moleculelor imbatranite cu altele tinere cu mentinerea unui nivel constant al lor in organism. Exista o relatie direct proportionala intre rata de catabolizare (K), concentratia de imunoglobulina (A) si timpul (T) exprimat in zile, relatia fiind K=A/T, in care K este rata de catabolizare intr-o singura zi. Deci T 1/2 este cu atat mai lung cu cat rata sa de catabolizare, este mai mica. Dar in afara de rata de catabolizare T 1/2 mai este dependent si de clasa de imunoglobuline, de specia animala, de viteza de sinteza a moleculelor, respectiv de nivelul concentratiei lor in ser, etc. Pentru determinarea valorii acestui timp, moleculele sunt marcate radioactiv, de regula cu 125I, dupa care se urmareste prezenta iodului in organism. Asa s-a determinat ca la om T 1/2 a IgM este de 5,1 zile, iar la IgG de 22 zile. La soarece, animal cu greutate corporala mica si cu o activitate metabolica intensa, T 1/2 a IgG este de numai 3,9 zile, in timp ce la animalele cu greutate corporala mare, valoarea lui este mult mai mare.
Procesul de degradare al Ig se realizeaza la nivelul ficatului precum si la nivelul altor organe bogate in sistem reticulo-histiocitar.
Mecanismele care stau la baza catabolizarii moleculelor de imunoglobuline nu sunt bine cunoscute fapt pentru care au fost emise diferite ipoteze prin care se incearca explicarea lor.
O ipoteza a 'catabolizarii selective', sustine ca molecula de imunoglobulina, atunci cand imbatraneste, isi modifica conformatia devenind straina de organism. Acesta are preformati anticorpii care recunosc moleculele modificate, se combina cu ele, formand complexe care sunt fagocitate si eliminate din organism de catre celulele din seria monocitar-macrofagica. O alta ipoteza a receptorilor de protectie, sustine ca celulele din seria monocitara care catabolizeaza imunoglobulinele, au receptori de membrana pentru Fc. Celulele pot lega moleculele din ser fie direct, fie prin intermediul acestor receptori. Moleculele fixate la receptori sunt trecute prin filtrul celular si eliminate extracelular fara a fi distruse de catre celula. Moleculele fixate, odata ajunse in celula in lipsa protectiei pe care o confera receptorii, sunt atacate de catre 'protein-disulfid-reductaza', o enzima care desface legaturile dintre lanturile H sau dintre H si L. Lanturile odata eliminate devin susceptibile la actiunea catepsinelor care le vor descompune in continuare pana la nivel de aminoacizi.
La hipergamaglobulinici, deci in exces de molecule de imunoglobuline, T 1/2 scade la jumatate fata de valoarea normala (cca. 12 zile), iar la hipogamaglobulinici creste de aproximativ 3 ori fata de normal (cca 70 zile). Catabolizarea claselor IgM si IgA nu este influentata de concentratia lor in ser.
1.4. Dinamica imunoglobulinelor in raspunsul imun umoral
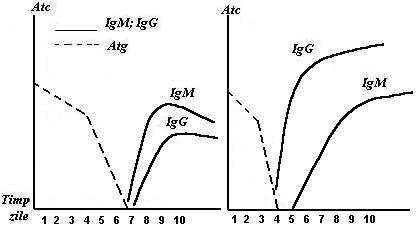
In cazul unei stimulari antigenice, organismul va reactiona prin sinteza de Atc., acest raspuns fiind cunoscut sub denumirea de raspuns primar. Prima categorie de anticorpi specifici care apar sunt anticorpii IgM, a caror crestere este de 80 molecule/sec. si au tendinta de epuizare rapida. Ulterior apare IgG a caror crestere este lenta si progresiva, cu tendinta deseori de descrestere. Daca organismul revine in contact cu acelasi Atg. apare raspunsul secundar (reactia anamnestica) in care cresterea IgM este neglijabila si se epuizeaza rapid, in timp ce anticorpii IgG cresc intens si rapid, atingand un nivel ridicat si persistent, urmata de scaderea lenta pana la un anumit prag de protectie imuna reziduala.
Diferentele intre raspunsul imun primar si cel secundar constau in scurtarea timpului de aparitie a efectorilor imuni si titrul mult mai ridicat al acestora, indiferent de doza de antigen reinoculata. (Fig.23).
Raspunsul
secundar se bazeaza pe persistenta in organism a limfocitelor de memorie care,
la al doilea stimul antigenic, sunt responsabile de promtitudinea si
intensitatea raspunsului imun.
Fig. 23 Dinamica imunoglobulinelor in raspunsul imun primar
si secundar
1.5. Unele paticularitati privind imunoglobulinele la animale
La diferite specii de animale, imunoglobulinele inregistreaza variatii in ce priveste antigenicitatea, greutatea moleculara, viteza de sedimentare, proportia lor in diferite umori, variatii conditionate de o mare diversitate de factori ca: specia, rasa, varsta, starea de intretinere, anotimpul, etc.
Taurinele si ovinele au urmatoarele caracteristici in ce priveste imunoglobulinele.
NORHEIM si col. (1985) au cercetat relatia intre continutul in imunoglobuline si varsta, zona geografica, anotimp, tehnologia de crestere si starea epizootologica a efectivelor si au observat valorile maxime ale IgG serice in perioada august-octombrie si minime primavara (martie-mai). Efectul anotimpului asupra cantitatii de imunoglobuline a fost mai pregnant la vitei in varsta de 31-90 zile. Continutul in imunoglobuline a fost influentat in sens pozitiv la cresterea viteilor in grup si pe asternut si in sens negativ la tinerea lor in boxe individuale, alaturi de vacile mame si de absenta asternutului.
Tabelul 5
Imunoglobulinele la bovine
|
Caractere si proprietati |
IgG |
IgM |
IgA |
||
|
IgG1 |
IgG2 |
Ser |
Secretii |
||
|
Numar subclase | |||||
|
Greutatea moleculara | |||||
|
Constanta de sedimentare (S20 W) | |||||
|
Mobilitatea electroforetica |
si | ||||
|
Procent hidrati de C | |||||
|
Concentratia de ser * | |||||
|
Concentratia in saliva* |
|
||||
|
Concentratia in colostru* | |||||
|
Timp de injumatatire (zile) | |||||
|
Fixarea complementului | |||||
|
Fixarea la piele | |||||
* g/100 ml; + = reactie pozitiva;
0 = reactie cu caracter absent;
? = necunoscut.
Zona geografica a influentat in mod constant concentratia serului in imunoglobuline. Autorii opineaza ca titrul de IgG seric ar putea constitui un indicator uzual al influentei factorilor ecologici asupra sanatatii viteilor.
La varsta de 100 zile, concetratia serului in IgG se stabilizeaza itrucatva la 18 mg/ml, fiind mai putin supus variatiilor in functie de factorii de mediu.
Desi s-a reusit izolarea si identificarea clasei IgE de origine bovina, totusi, la bovine si ovine clasele IgE si IgD au fost putin studiate.
Tabelul 6
Imunoglobulinele la ovine
|
Caractere si proprietati |
IgG |
IgM |
IgA |
||
|
IgG1 |
IgG2 |
Ser |
Secretii |
||
|
Numar subclase | |||||
|
Greutatea moleculara | |||||
|
Constanta de sedimentare (S20 W) | |||||
|
Mobilitatea electroforetica |
si | ||||
|
Procent hidrati de C (%) | |||||
|
Concentratia de ser* | |||||
|
Concentratia in saliva* | |||||
|
Concentratia in colostru* | |||||
|
Concentratia in lactoser* | |||||
|
Fixarea complementului | |||||
* g/100 ml; + = reactie pozitiva;
0 = reactie cu caracter absent;
? = necunsocut.
Bubalinele sunt apropiate in ce priveste clasele si subclasele de imunoglobuline de taurine.
Cabalinele au acelasi echipament in imunoglobuline existent la om si celelalte specii animale, dar cu unele deosebiri caracteristice pentru aceasta specie. Cabalinele par sa aiba mai multe izotipuri, fiind astazi admise la aceasta specie cel putin sase clase.
IgT reprezinta o clasa specifica ecvinelor, care a fost initial considerata ca fiind analoaga cu IgA a altor specii, de aceea poate fi intalnita in lucrari mai vechi si sub notatia IgAT. Ulterior, s-a dovedit insa a fi mai apropiata de IgG decat de IgA; lanturile ei usoare sunt antigenic identice cu cele ale IgG; In privinta compozitiei in aminoacizi a fractiunilor Fc se constata insa o diferenta fata de IgG (14 aminoacizi la IgT, fata de 19 la IgG). Masa moleculara a IgT este de 150000 da.
IgB, descrisa si sub simbolul AI (agregating imunoglobulinis), reprezinta, ca si IgT, o alta imunoglobulina specifica ecvinelor, careia fortele inonice ii asigura un slab grad de agregare. Masa ei moleculara este de 160 000 da., iar coeficientul de sedimentare variaza intre 7 S si 19 S.
Datele privind IgD si IgE la cabaline sumt sumare si neconcludente.
Suinele poseda in umorile organismului izotipurile clasice de imunoglobuline; studii mai detaliate au fost realizate privind IgG,IgM, IgA si intr-o masura mai redusa IgE.
Pasarile contin in umori aceleasi clase de imunoglobuline ca si mamiferele. IgG si IgM au fost identificate in sange ca principalii anticorpi serici, iar IgA in secretii, majoritatea lucrarilor referindu-se la prezenta ei in bila.
Valorile imunoglobulinelor, releva faptul ca IgA si IgM sunt prezente in tubul digestiv al embrionului si provin mai degraba din albus decat din galbenus. IgG din intestin pare sa-si aiba originea in galbenus si sa ajunga in embrion prin lichidul amniotic, in care in a 12-a zi este inca absenta, dar poate fi detectata in a 17-a zi de viata.
Datele de mai sus sugereaza o analogie intre modalitatile de transfer vertical pasiv a imunoglobulinelor la pasari cu cea de la mamifere.
1.6. Anticorpii monoclonali. Tehnica hibridomului.
Serurile, contin mai multi anticorpi in functie de determinantii (epitopii) antigenului. Deci, serurile conventionale 'clasice' sunt policlonale, avand si anticorpi inutili de balast. Inconvenientul serurilor policlonale care contin mai multi Atc., consta in nespecificitatea unor reactii si imposibilitatea de a recunoaste antigenii comuni. Deasemenea mai intervin si alte dificultati:
Ø dificultatea de a discrimina diversele tipuri de virusuri sau bacterii (virusul herpetic I de virusul herpetic II);
Ø dificultatea de a discrimina hormoni care au fragmente comune (hormonul foliculostimulant hipofizar -FSH, de hormonul gonadotrop corionic uman-HCG);
Ø dificultati in dozajul imunologic al medicamentelor (digoxina si teofilina) care dau reactii incrucisate datorita produsilor lor de metabolism;
Ø lipsa de omogenitate din cauza reactivitatii individuale a producatorului de ser.
S-a nascut in felul acesta ideea unui ser monospecific.
1.6.1. Tehnici de preparare a anticorpilor monoclonali
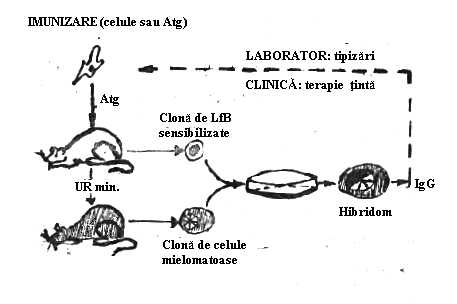
G. KOHLER si C.MILSTEIN (1975 Cambridge) cautand structura sau modalitatea fuzionarii celulare 'in vitro' ajung printr-o fericita intamplare sa descopere principiul prepararii anticorpilor monoclonali. In formarea anticorpilor monoclonali intervin doua celule: limfocitul B luat din splina de soarece, imunizat contra hematiei de oaie care se cloneaza si se ajunge la o clona specifica si celula mielomatoasa de soarece (linia P3).
Principiul producerii anticorpilor monoclonali consta in fuzionarea acestor doua celule. In ceea ce priveste necesitatea de fuzionare si de participarea fiecaruia dintre celule la edificarea anticorpilor monoclonali, limfocitul B vine cu informarea antigenica iar celula mielomatoasa vine cu doua proprietati:
Ø celula mielomatoasa produce Ig, dar totdeauna de un singur tip (de obicei IgG);
Ø aceasta celula are capacitatea de diviziune practic infinita (nu i se deceleaza modificari de cariotip nici dupa 20 de generatii).
Se obtine in acest mod o celula noua de tip 'himera' denumita hibridom (Fig.24). Pentru obtinerea limfocitului de tip B utilizat in reactia de fuzionare s-a utilizat ca Atg. hematia de oaie, care a fost purificata de o serie de epitopi ce nu erau specifici lasandu-i-se numai epitopii ce confera specificitatea stricta pentru acest Atg. S-a inoculat acest Atg. direct in splina la soarece.
Dupa un anumit timp, necesar formarii de Atc. (7 - 14 zile) se preleveaza limfocite din splina. Celulele mielomatoase se obtin prin inocularea soarecilor cu uleiuri minerale intraperitoneal obtinandu-se mieloame (cancere ale sistemului imunitar) si implicit linii mielomatoase murine (linii X63; NS1; MSO1; etc.).
 |
Urmeaza partea cea mai dificila, hibridarea, adica punerea in contact a limfocitului normal cu celula mielomatoasa. Fuziunea celulara se face prin doua modalitati:
- a) in prezenta polietilenglicolului;
- b) prin electrofuziune.
Prin metoda cu polietilenglicolul numai un limfocit dintr-o populatie de 200 000 este capabil de fuzionare.
In metoda electrofuziunii rata de fuzionare creste la 80% si aceasta presupune existenta unui ecran microscopic in care se evalueaza apropierea celulelor limfocitare de cele mielomatoase.
In momentul in care in campul microscopic se observa mai multe apropieri se face o descarcare de curent electric in lichid, producand fuzionarea celulelor.
Momentul urmator consta in clonarea celulelor fuzionate pentru a gasi si a alege numai ser monoclonal cu Atc. specifici.
Celula hibridom are mostenire de la celula mielomatoasa capacitatea de a sintetiza un singur Atc. Clonarea celulelor fuzionate are loc pe mediul HAT (hipoxantina- aminoterina- timidina).
Celulele hibridom se mentin in conditii de laborator pe culturi de tesuturi si se inoculeaza intraperitoneal la soarecii 'nuzi' care sunt lipsiti de timus si nu au par. Acestia nu au nici un fel de aparare imunologica. In urma inocularii celulelor mentionate, acestia fac tumori limfomatoase iar lichidul ascitic recoltat de la subiectii in cauza este foarte bogat in anticorpi monoclonali. In esenta modelul Kohler-Milstein de obtinere a celulei hibridom propune urmatoarele etape:
Ø purificarea antigenelor cu obtinerea epitopilor specifici;
Ø inocularea antigenului purificat la soareci;
Ø scoaterea splinei si recoltarea limfocitelor B la 7 - 14 zile de la inoculare;
Ø hibridarea ( fuziunea limfocitului cu celula mielomatoasa);
Ø clonarea celulelor fuzionate pe mediul HAT;
1.6.2. Aplicatiile in practica ale Atc. monoclonali.
Anticorpii monoclonali prezinta o serie de avantaje deosebite in diagnosticul foarte divers imunologic. De ex.:
Ø diferentierea tipurilor de bacterii sau virusuri, strans inrudite;
Ø identificarea chlamidiilor;
Ø identificarea rapida si specifica a germenilor din diareile nou-nascutilor;
Ø identificarea salmonelelor in timp scurt (2 ore) din alimente (procedeul clasic durand 6 zile);
Ø identificarea virusurilor respiratorii sincitiale;
Ø dozarea progesteronului;
Ø depistarea si supravegherea unor celule maligne in special din colon si rect;
Ø pentru identificarea populatiilor si subpopulatiilor de limfocite, etc.
In vederea diagnosticului se prepara 'kitt-uri' de Atc. monoclonali de catre diverse societati farmaceutice occidentale, cifra de afaceri in acest sector atingand nivelul de 500 mil.$. Dificultatile obtinerii Atc. monoclonali, pe langa cele tehnice, sunt legate in primul rand de gasirea epitopilor specifici.
1.7. Citokinele (monokine si limfokine)
Efectul de cooperare T-B poate fi demonstrat in culturi in care cele doua populatii celulare sunt separate de o membrana cu pori foarte fini. Suplimentarea mediului de cultura cu diverse populatii celulare induce activarea sau inhibitia acestora. Reiese ca celulele implicate in reactia imuna pot comunica intre ele prin factorii solubili care poseda anumite activitati biologice.
Mcf. si LfT sunt surse principale de citokine. De aceea produsele acestor celule mai sunt denumite si monokine sau, respectiv, limfokine.
Monokinele si limfokinele sunt secretate numai de celule activate fie specific (in urma contactului cu Atg.), fie nespecific (de catre diverse citokine). Activarea prin citokine a celulelor implicate in raspunsul imun atrage dupa sine recrutarea unui numar mare de celule la locul conflictului cu Atg. si implicit amplificarea reactiei de aparare. Aceasta interactiune intre diferite celule a dus la formarea termenului de 'interleukine' pentru regruparea acestor limfokine si monokine care induc relatii reciproce de stimulare intre celulele partenere.
Citokinele sunt glicoproteine si cu toate ca exista anumite familii de substante care au un precursor comun, ele se deosebesc de Ig, deoarece nu poseda nici aceeasi structura moleculara de baza si nici nu demonstreaza o homologie importanta a secventelor lor de aminoacizi cu acele ale Ig.
Citokinele nu sunt specifice: mai multe celule diferite pot secreta dupa stimularea antigenica sau nespecifica o aceeasi substanta; substanta respectiva poate actiona pe mai multe tipuri celulare carora le induce activitati biologice diferite.
Majoritatea citokinelor actioneaza asupra celulelor-tinta de o maniera analoaga celei a hormonilor peptidici: fixarea pe un receptor specific membranei, activarea unui mesager secundar intracelular care transmite semnalul la nucleu si induce astfel aparitia efectului biologic specific.
Numarul si densitatea receptorilor pe suprafata celulelor tinta sunt bine determinate. Efectul biologic al citokinelor nu este in mod necesar corelat cu numarul receptorilor exprimat pe celula. Afinitatea receptorului pentru ligandul sau fiziologic poate avea un rol important in acest sens.
1.7.1. Interleukinele
Principalele substante din aceasta clasa sunt:
IL 1 secretata de celule prezentatoare de Atg. (Mcf., celule Langerhans), granulocite.
Productia de IL 1 creste substantial dupa activarea celulelor monocito-macrofage. Aceasta activare poate fi indusa dupa:
endocitarea Atg. (in cadrul procesului de 'prezentare' a acestuia)sau
prin actiunea altor citokine asupra Mcf. (IL2, IFN gama).
IL 1 actioneaza pe mai multe substraturi celulare. Contribuie la activarea limfocitelor B si T. Pentru ca aceasta activare sa se produca este nevoie ca asupra limfocitelor sa opereze simultan mai multe semnale. IL 1 ar furniza unul dintre acestea. Ca raspuns la stimulare, limfocitele vor secreta si alte limfokine (IL 2, IL 3, IFN gama.). Stimuleaza productia de prostaglandina din celulele endoteliale prin intermediul careia induce febra. Stimuleaza hepatocitele sa produca proteine de faza acuta cu rol in apararea nespecifica cum ar fi proteina C-reactiva. Mareste capacitatea functionala a granulocitelor si mobilizarea lor chemotactica.
IL 2 T-cell growth factor): este produsa in special de LfTh. Secretia IL 2 necesita doua semnale: Atg. (prezentat pe suprafata Mcf.) si IL 1. In stare de repaos LfTh exprima pe membrana un numar mic de receptori pentru IL 2. Dupa receptarea semnalelor de activare se produce initial o crestere rapida a numarului R IL 2, dupa care are loc secretia de IL 2. Aceasta va actiona asupra propriei celule producatoare (efect autocrin) si asupra altor celule T pe care le activeaza nespecific. Se pare ca IL 2 activeaza toate tipurile de subpopulatii T (Th, Ts, Tc) care raspund identic prin exprimare R IL 2 si proliferare. In plus, IL 2 activeaza si alte celule (LfB, Mcf., celulele NK) si ar determina cresterea activitatii 'killer' a limfocitelor.
Glucocorticoizii inhiba productia de IL 2, prin intermediul inhibarii secretiei de IL 1 de catre Mcf.
IL 3 este produsa de LfT activate de Atg. si are efecte functionale multiple. Din cauza ca promoveaza proliferarea 'in virto' a celulelor stem multipotente si eritroide (din care rezulta formarea de colonii celulare) a primit si numele de 'multi-CSF' (CSF - colony stimulating factor). Rolul sau in imunitate este neclar: se pare ca ar contribui indirect la diferentierea LfT si la cresterea activitatii citotoxice a macrofagelor. La acestea se mai adauga efectele asupra sistemului hematopoietic care furnizeaza majoritatea celulelor auxiliare cu rol in aparare.
IL 4 ar actiona ca factor de proliferare a LfB (de aici si vechea denumire a substantei: BCGF1-(B-cell growth factor 1). Actioneaza intr-o etapa a activarii LfB, cand aceste celule exprima receptorii sai specifici si poate induce secretia de IgG1 (o subclasa de IgG care apare rareori in raspunsul imun). In plus ar putea conferi activitate citotoxica macrofagelor normale si ar induce proliferarea mastocitara.
IL 5 (BCGF II-B cell growth factor II) ar induce la soareci proliferarea LfB si secretia de IgM, IgG si IgA. La om rolul IL 5 este incomplet elucidat. Este un produs al limfocitelor T (un astfel de factor a fost izolat din culturile de celule T umane stimulate cu HTLV-1 (Human T-cell lymphoma/leukemia virus I).
IL 6 produsa de Mcf. actioneaza sinergic cu IL 3 asupra celulelor stem multipotente ale hematopoiezei. Este implicata in activarea LfT si in inducerea productiei proteinelor de faza acuta. Sinteza IL 6 este stimulata de IL 1.
IL 7 este produsa de celulele stromale ale maduvei osoase. Induce diferentierea limfocitelor foarte tinere (pre-B) dar nu are efecte asupra limfocitelor B mature.
IL 8 a fost cunoscuta la `nceput ca un factor proteic de activare a neutrofilelor, face parte din familia chemokinelor (citokine pro-inflamatorii). Astazi se stie ca IL-8 este o citokina produsa de monocite, macrofage, neutrofile, LT, celulele endoteliale, epiteliale si fibroblaste ca raspuns la o varietate de stimuli care include LPS, TNF-alfa, IL-1, IL-7 precum si `n cazurile de hipoxie. Are proprietate de chemotactism asupra neutrofilelor, bazofilelor, LT, efecte proinflamatoare ( degranularea neutrofilelor si bazofilelor, stimularea metabolismului oxidativ si aderentei PMN) si angiogenetice (neoformare de vase sanguine). De asemenea determina sporirea actrivitatii enzimelor lizozomale si exprimarea receptorilor pentru complement.
IL 9 este un factor de proliferare pentru LT si mastocite, fiind secretata de LT activate (CD4+) si T cu memorie. Rolul biologic este multiplu, prin actiunea asupra LB, LT, precursorilor hematopietici si mastocitelor. La nivelul LB ea stimuleaza productia de IgG, IgE (sinergism cu IL-4) iar al LT amplifica proliferarea LT din sangele periferic, fiind totodata un stimulator al precursorilor eritropoietici (alaturi de EPO), hematopoietici si al diferentierii mastocitelor ( mai ales `n parazitozele intestinale).
Actiunea IL-9 se manifesta `mpreuna cu alte limfokine produse de celulele Th 2 ca IL-3, IL-4 si IL-10. Receptorii pentru IL 9 sunt prezenti pe diferite celule cum ar fi Th, clone celulare de limfoame, macrofage si mastocite.
IL 10 a fost descrisa `n 1989, produse de clone de celule Th2 ce inhiba sinteza de citokine de catre un alt set de clone Th1.
Cand a fost identificata, i s-a atribuit rolul de factor inhibitor al sintezei unor citokine proinfalamtorii produs de limfocitele Th2. ~n prezent se stie ca interleukina 10 (IL-10) este produsa de mai multe tipuri de celule: LT CD4+ si CD8+ ( `n repaus si cu memorie), LB, monocite, eozinofile, mastocite, celule epiteliale bronsice, unele tipuri de celule canceroase etc. Efectele biologice sunt pleiotrope imunomodulatoare: pe de o parte anti-inflamatoare si imunosupresoare, iar pe de alta imunostimulatoare.
Se poate spune ca principalele actiuni biologice ale IL-10 sunt cele inhibitorii asupra activitatii monocitelor/ macrofagelor, ale LTh1 si antiinflamatorii.
IL 11 a fost identificata `n supernatantul unei linii celulare de maduva osoasa.
Interleukina 11 (IL-11) este secretata de LT si are efecte stimulatoare asupra LB. Exista date care sugereaza ca IL-11 `n conjunctie cu IL-3 induce formarea precursorilor megacariocitari prin cresterea lor numerica si `n dimensiuni. De asemenea este activata proliferarea si productia de Ig de catre LB.
IL 12 a fost izolata `n 1989 si reuneste structural elemente `ntalnite la alte citokine. IL-12 este o citokina heterodimerica de 70 kDa care este produsa de APC profesioniste ( macrofage, celule dendritice, LB activate) si are o dubla actiune- proinflamatorie si imunoreglatoare.
IL 13 descoperita aproape concomitent cu IL-12 are functii "de tip Il-4", dar mai putin pregnante. Ea este o proteina de 10 kDa produsa de LT CD4+ (Th0, Th1, Th2) si CD8+ activate, care actioneaza la nivelul LB si monocitelor/macrofagelor.
IL 14 a fost identificata `n 1991; este produsa de LT si stimuleaza LB activate ( dar nu si pe cele `n repaus). ~n schimb inhiba sinteza de Ig `n cazul `n care LB este activat de mitogeni (ConA, PHA)
IL 15 a fost descrisa `n 1994, independent de doua grupe de cercetatori ca analog functional al IL-2), capabil sa stimuleze LT si NK. Sursa principala de Il-15 `n organism o constituie placenta, dar este exprimata consecutiv si `n alte tesuturi ( plaman, ficat, rinichi, inima, muschi striati) sau celule (monocite/macrofage, celule stromale medulare, epiteliale). Asemanator situatiei `nmtalnite la IL-13, Il-15 ar putea fi o forma alternativa a IL-2 necesara `n limfopoieza si/sau modularea raspunsului imun, avand `n plus activitate de proliferare a mastocitelor.
IL 16 este produsa de LT (Tc CD8+ si CD4+), mastocite, eozinofile, celule epiteliale din caile respiratorii, are actiune proinflamatorie si imunomodulatoare. Ea este chemoatractanta pentru celulele CD4+ (LT monocite) si eozinofile, recrutandu-le la locul inflamatiei. Stimuleaza exprimarea receptorului pentru IL-2, IFN gamma si a celulelor de adeziune la matricea extracelulara a monocitelor si eozinofilelor.
IL 17 este o citokina identificata recent, care are activitate de stimulare a secretiei de citokine proinflamatoare, chemokine si prostaglandine de catre fibroblasti, celule epiteliale si endoteliale si a exprimarii moleculelor de adeziune de catre aceste celule. Efectele ei inflamatoare sunt similare cu cele ale TNF si LT. ~n plus are efecte hematopoietice, argumeteaza producerea de Il-6, G-CSF si de neutrofile.
IL 18, sinonime: factor de inductor al sintezei de IFN gamma (IGIF-IFN gamma- inducing factor, IL-1 gamma). Il-18 a fost descoperita de echipa lui Okamura `n 1995 ca o consecinta a capacitatii acesteia de a induce producerea de IFN gamma la soarecii cu soc endotoxic. IL-18 are o greutate moleculara de 18,3 kDa.
~n prezent se considera ca aceasta citokina este un nou membru al familiei de citokine si anume Il-18
IL-18 este exprimata larg, dar nu universal, pe diferite tipuri tisulare, fiind detectata `n pancreas, rinichi, muschi scheletic, ficat, plaman, os (osteoblaste) si piele (keratinocite). A fost identificat si receptorul pentrul IL-18 (IL-18R) care este exprimat mai ales `n splina, timus, leucocite, ficat, plaman, inima, intestin, prostata si placenta (dar nu si `n creier), muschi scheletici, rinichi, pancreas.
Il-18 actioneaza, asemanator IL-12, ca un factor stimulator asupra LTh1 si are un efect de activare a celulelor NK.
1.7.2. Interferonii
IFN gama este produs de LfT.IFN gama face parte dintr-o familie de substante cu activitate imunomodulatorie, antivirala si antitumorala.
IFN gama este implicat in urmatoarele activitati: actioneaza sinergic cu alte citokine in inducerea productiei de Atc. si sporeste activitatile de tip citotoxic ale Mcf, LfT si celulelor NK asupra celulelor transformate.(Fig.25)
1.7.3. Factorii de necroza tumorala
Limfotoxina (TNF beta) inrudita cu factorul precedent este produsa de limfocitele T activate si are efecte citolitice asupra celulelor tinta (tumorale, straine sau alterate de agresiunea antigenica).
Factorul de necroza tumorala (TNF alfa = tumor necrosis factor alfa) sau 'casectina' este produsa de macrofage. Pe lamga efectul de distrugere a celulelor tumorale 'in vitro', TNF alfa coopereaza sinergic cu IL 2 in urmatoarele activitati: activarea chimiotaxiei si a functiei de bactericidie a granulocitelor, activarea limfocitelor T (care la randul lor vor produce IL 2 si IFN gama si alti factori stimulatori), reglarea productiei de anticorpi, eliberarea activa de proteine de faza acuta din hepatocite.
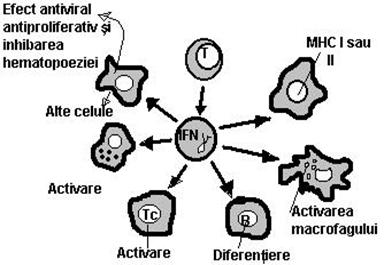
Fig.25. Schematizarea efectelor IFN asupra mai multor categorii de celule.
1.7.4. Neuroleukinele sunt substante care permit examinarea noilor interactiuni intre sistemul nervos si reteaua de citokine.
Proteina murina matura, comportand 588 acizi aminati este un factor de crestere pentru anumiti neuroni. Ea este produsul glandelor salivare, muschiului striat, creierului, dar mai ales al limfocitelor T stimulate prin lectine. Din punct de vedere imunologic, in mod cert, neuroleukina joaca un rol impreuna cu IL 4 si IL 5 in activarea expansiunii clonale a celulelor B. In mod curios, ea prezinta o foarte mare homologie cu o enzima (fosfoexoza izomeraza) importanta in glicoliza.
1.7.5. Factorii de stimulare ai coloniilor celulare
CSF (Factorul stimulator de colonii)
Numeroase citokine sunt capabile, printre alte activitati sa stimuleze cresterea uneia sau mai multor linii hematopoietice. Acest lucru este evident in cazul IL 3, dar si in cazul IL 1, IL 6, IL 7.
Ultimele 3 sunt considerate in acest caz drept CSF, avand acelasi titru ca si IL 3, dar fara a fi disociat decat dintr-o ratiune de nomenclatura.
GM -CSF (factorul stimulator al coloniilor de granulocite si macrofage)
El se defineste ca o citokina care stimuleaza proliferarea liniilor precursoare pentru macrofage si granulocite. El activeaza in mod egal si polimorfonucleare mature.
La om, el reprezinta un lant polipeptidic de 127 acizi aminati, cu care proteina murina prezinta 50% homologie.
Gena corespondenta comporta 3 introni si a fost localizata pe regiunea q23-q31 a bratului lung al cromozomului 5 de la om.
Este produs in principal, de macrofage, el putand fi in mod egal produs de catre anumite linii de celule tumorale, de celule stromale ale maduvei osoase prin limfocitele T activate, fibroblaste si celulele endoteliale.
G-CSF (factorul stimulator al coloniilor de granulocite)
Acest factor stimuleaza in mod particular linia precursoare de granulocite.
El reprezinta un lant de 177 aminoacizi, fara sa prezinte homologii cu GM -CSF sau IL-3. Gena corespondenta este localizata pe bratul lung (q11,2-q21)al cromozomului 17 la om.
M-CSF sau CSF-1 (factorul stimulator al coloniilor de macrofage)
Acest factor stimuleaza proliferarea liniei precursoare a monocitelor.
Proteina matura contine 224 aminoacizi. Homologia cu proteina murina este probabil crescuta (de ordinul 78% pe o parte a secventei). Gena corespondenta comporta 10 exoni la om si este localizata la nivelul regiunii q33 ml al bratului lung al cromozomului 5.
M-CSF poate fi produs de fibroblaste, celule endoteliale sau macrofage.
1.7.6. Prostaglandinele
Sunt compusi biologici activi, produsi practic de catre toate tesuturile si celulele care au suferit un proces de stimulare la nivelul membranei plasmatice. Sunt sintetizate local `n cantitati foarte mici si actioneaza scurt timp datorita metabolizarii lor rapide.Sunt printre cele mai raspandite si active substante din natura, prezenta lor fiind semnalata de la vietuitoare inferioare pana la specii din lumea vegetala.
Pe baza structurii si activitatii lor biologice s-au descris mai multe clase de PG : PGA, C,D,E,F,H, I iar `n cadrul acestor clase exista molecule cu una sau doua duble legaturi notate ca molecule de tip 1 sau 2. De exmplu, se vorbeste de PGE1 si PGE2
O sursa importanta de PG sunt acizii grasi nesaturati, hidrolizabili si polioxigenati, care contin un inel de ciclopentan si au un schelet de baza alcatuit din acid prostanoic. Acesta sub influenta fosfolipazelor, se transforma `n acid arahidonic care este un precursor al tuturor moleculelor de prostaglandine. Sub influenta lipooxigenazelor, are loc formarea de leucotriene (LT), substantele cu implicatii majore `n desfasurarea proceselor inflamtorii.
Initial s-a considerat ca PG au efect fiziologic predominant asupra muschilor netezi si `n inducerea proceselor inflamatorii. Ulterior, s-a demonstrat clar ca au multiple functii biologice: intervin `n fiziologia muschilor netezi, `n transmiterea influxului nervos, `n interactiuni hormonale si implicit `n procesele imune.
Prostaglandinele influenteaza comportarea celulei, inducand schimbari `n functiile membranei, careia `i altereaza compozitia `n lipide, proteine si glicoproteine, inducand schimbari `n fluiditatea si permeabilitatea membranei.
1.7.7. Factorii de transformare a cresterii (TGF
TGF formeaza o familie de citokine produse de LT activate de catre antigene si de catre celulele mononucleare activate bacterian. Cuprind doua tipuri de proteine notate cu si formand citokinele TGF- si TGF-
TGF- este implicata `n cresterea si diferentierea celulelor mezenchimale si epiteliale.
TGF- este principalul factor de transformare a cresterii avand o actiune stimulatoare sau inhibitoare asupra celulelor tinta, `n functie de tipul celular si de interactiunea cu alti factori celulari. TGF- moduleaza cresterea, diferentierea si activitatea LB, TLTh, LTc, celulelor NK si LAK
TGF-β inhiba activarea celulelor NK precum si productia de citokine a LT, fiind considerata o citokina importanta cu rol imunosupresor al expansiunii celulare excesive `n cadrul raspunsului imun.
Descoperirea citokinelor are o mare importanta fundamentala. Ea a demonstrat ca in cadrul raspunsului imun se produce de fapt o comutare de la un mecanism dirijat in mod specific de Atg. la un sistem universal de reglare de tip hormonal.
Deoarece citokinele actioneaza ca imunomodulatori, dirijarea activitatii lor ar putea conastitui noi linii de terapie mai dezvoltate la boala neoplazica.
Ø adminstrarea de IL 2 ar duce in anumite conditii la selectarea unor celule cu potential citotoxic antitumoral ('lymphokyne activated killer cells'= LAK). In ciuda unor rezultate promitatoare la bolnavi cu cancer avansat, metoda ramane limitata de efectele toxice ale IL 2 (produce grave tulburari ale permeabilitatii capilare cu anazarca);
Ø diversi IFN preparati prin inginerie genetica sunt utilizati cu succes in mai multe afectiuni neoplazice: leucemia cu celule paroase, limfoame cutanate, leucemia granulocitara cronica, sarcomul Kaposi secundar sindromului SIDA; in hepatita cronica activa tratamentul cu IFN, grabeste eliminarea virusurilor HBs si HBc; in asociere cu acyclovir IFN constituie tratamentul de selectie al keratitei herpetice;
Ø unele boli autoimune ar fi corelate cu o activare dezorganizata a LfT. Recent s-a descoperit o substanta extrasa dintr-o ciuperca Cyclosporina A) care este capabila sa inhibe selectiv LfT fara efecte secundare asupra hematopoiezei. Pe aceste baze s-a conceput administrarea Cy-A in aplazia medulara idiopatica. Cy-A mai are un domeniu de aplicare major: la receptorii de grefe de organ pentru stapanirea fenomenelor de respingere.
2. EFECTORI CELULARI AI IMUNITATII
Celulele limfocitare sunt responsabile de specificitatea reactiilor imunologice.
In sanul populatiei limfocitare se disting doua subpopulatii cu functii distincte: limfocitele T care asigura imunitatea celulara si limfocitele B care asigura imunitatea umorala.
In favoarea limfocitului ca celula 'omniprezenta' in procesul de imunitate specifica, pledeaza o serie de argumente de ordin filogenetic, ontogenetic, clinic si experimental.
Argumente de ordin filogenetic. Procesele de aparare specifica, nu apar decat la speciile de animale la care apar celule, care morfologic pot fi identificate ca limfocite (incepand cu ciclostomii).
Argumente de ordin ontogenetic. In cursul dezvoltarii embrionare, fetusul la vertebrate nu castiga imunitate specifica, decat in momentul cand apar primele celule limfocitare. La caine de exemplu, raspunsul imun apare in a 40 zi de viata fetala, cand limfocitele apar in timus si in alte organe limfoide (maduva osoasa si ganglionii limfatici).
Argumente de ordin clinic. S-a constatat ca in sindromul de deficit imunitar secundar la om ( Sindroamele Glazmann-Riniker si Good), nu pot fi decelate, la individul bolnav, celulele cu aspect limfocitar. Sindromul de deficit imunitar, se traduce prin lipsa oricarui raspuns imunitar, dupa o stimulare antigenica.
Argumente de ordin experimental. Soarecii expusi in mod experimental unei iradieri (Rx) cu doze subletale, fac o aplazie limfocitara, consecutiva iradierii. Aceasta aplazie este insotita de lipsa oricarui raspuns imunitar fata de o incitatie antigenica.
2.1 Ontogeneza limfocitelor
Diferentierea LfB si T are loc in OLC si parcurge mai multe etape care se caracterizeaza prin aparitia succesiva a unor molecule functionale in citoplasma sau sprafata celulara. Celulele maturate parasesc OLC si se distribuie in zonele B si respectiv T-dependente din OLP (homing), gratie unor receptori specifici de suprafata. Fenomenul de homing caracterizeaza Lf. pe toata durata vietii lor: un LfB, de exemplu, care paraseste un ganglion si circula in organism se va aseza in alt teritoriu limfoid dar tot intr-o zona B-dependenta. Celulele mature imunocompetente, care au parasit OLC si nu au intalnit inca Atg. specific sunt denumite 'virgine'. Dupa intalnirea cu Atg. aceste celule sufera o serie de transformari morfologice si structurale care se finalizeaza cu o mitoza. Celulele 'transformate' au o talie foarte mare si citoplasma intens bazofila. Ele au fost denumite celule 'blastice' (deoarece au o infatisare asemanatoare cu elementele medulare tinere -'blastice'- din care se formeaza celulele sanguine) sau 'imunoblasti'. Prin mitoza imunoblastii genereaza 2 celule-fiice diferite: una va parcurge pana la completare programul de maturatie al liniei celulare respective si va deveni celula efectoare a raspunsului imun (RI), cealalata va ramane identica cu celula-mama persistand ca 'celula de memorie'. Compartimentul celulelor de memorie au o importanta functionala deoarece asigura mentinerea clonelor specifice pentru Atg. Fenomenul transformarii blastice poate fi reprodus 'in vitro' daca Lf. cultivate sunt incubate cu Atg. sau cu lectine vegetale (denumite si substante mitogene). Fitohemaglutinina si Concanvalina A stimuleaza transformarea blastica a LfT. Pokeweed mitogen produce transformarea si a LfT si a LfB. Testul de transformare blastica este curent utilizat in clinica pentru testarea functiei limfocitelor.
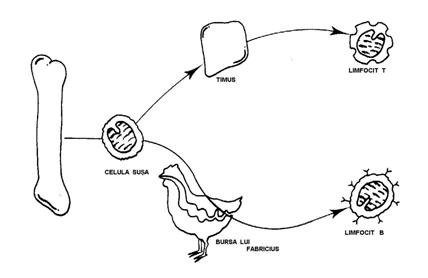
Maduva osoasa este sediul CSL care provin din diferentierea CS multipotente. CSL sunt bipotente: genereaza LfB si T. La mamifere si la om joaca un rol de organ burso-echivalent de diferentiere si de maturare a LfB. (Fig. 26)
Schema generala a distributiei celulelor limfoide
|
Compartimentul de diferentiere si de maturare |
Compartimentul functional |
|
|
Timus LfT Celule stem limfoide (CSL) M.O. LfB |
Ganglioni limfatici Splina Structurile limfoide asociate tractului digestiv si respirator, inelul limfatic Waldeyer, placile Peyer, apendicele, patura submucoasa. Altele: stratul dermic, etc. |
|
|
Organe limfoide centrale (OLC) (primare) |
Organe limfoide periferice (OLP) (secundare) |
|
|
aparitia precoce in cursul dezvoltarii embrionare; centrele limfopoiezei: initial diferentiere CSL* in celulele B si T imature: in continuare maturarea LfB si T si migrarea celulelor devenite imunologic competente in OLP; procesele de diferentiere-maturare sunt independente de Atg.; extirparea sau iradierea lor precoce, atrag dupa sine lipsa formarii Lf mature si imunodeficiente grave. Repopularea lor poate fi realizata numai cu celule stem |
se formeaza in urma OLC; sediile unde se distribuie LfB si T imunologic competente formate in OLC; LfB si T se repartizeaza preferential in zone microanatomice, distincte B-dependente (foliculii limfoizi) si T-dependente (zonele paracorticale din ganglioni si tecile periarteriale din splina); trafic permanent de Lf intre diversele OLP; dupa iradiere corporala totala pot fi repopulate cu Lf mature; sedii ale raspunsului imun; |
|
* CSL - celule stem limfoide
2.2. Ontogeneza limfocitelor B
a) etapa medulara (independenta de Atg). Evenimentul initial care anunta angajarea CSL in diferentierea catre linia B este rearanjarea genelor pentru lanturile H de Ig, care se materializeaza prin aparitia de lanturi miu intracitoplasmatice (c
Rearanjarile incep pe ambii cromozomi 14, dar se finalizeaza numai pe unul singur impiedicand astfel producerea de lanturi cu specificitati diferite in aceeasi celula. Ulterior se rearanjeaza si genele pentru lanturile L (K) si astfel se asambleaza in citoplasma molecule complete de IgM (cIgM). Celulele c si cIgM+ au fost denumite 'pre-B'. Intr-o etapa ulterioara IgM devine exprimata pe suprafata (sIgM) (celulele B imature). sIgM poate functiona ca receptor pentru Atg. si se presupune ca celulele B imature pot reactiona cu Atg. proprii daca moleculele de Ig exprimate pe suprafata sunt corespunzatoare. Aceasta interactiune nu ar reproduce modelul celulelor mature (transformarea blastica, mitoza, etc.) ci ar fi urmata de desprinderea IgM de pe suparafata. Deoarece LfB imature nu pot reface aceste sIg, ele vor ramane in continuare inactive. Acest fenomen de selectie negativa ar preveni dezvoltarea unor clone autoreactive si ar putea sta la baza tolerarii selfului. Celulele care nu au fost facute nereactive continua sa se maturizeze. In cursul maturarii sunt achizitionate pe suprafata si alti markeri: Atg. CMH de clasa I si II (DR), CR1, CR2,, RFcγ. LfB 'virgine' care parasesc MO exprima pe suprafata 2 molecule de sIg: IgM + IgD. Unele celule B sufera in procesul de maturare o comutare a sintezei IgM in alta clasa de Ig. In acest caz, grupul de gene VDJ care fusesera asamblate anterior cu o gena CH miu, ajunge sa se cupleze cu alte gene CH (Cgama sau Calfa, de exemplu). Noua Ig produsa de celula are deci acelasi idiotip cu IgM originala. Procesul de comutare explica variatiile sintezei Atc. in raspunsul imun: in raspunsul primar domina secretiile de IgM; in raspusurile secundare, Atc. dominanti apartin altor clase cu aceeasi specificitate (IgG sau IgA in exemplele citate).
b) etapa dependenta de Atg.
Dupa intalnirea cu Atg. specific, LfB mature sufera procesul de transformare blastica in urma caruia vor genera celule de memorie si plasmocite efectoare.
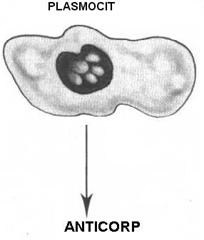
Plasmocitul reprezint stadiul terminal al ontogenezei LfB(Fig.27).
~n acest proces LfB pierd sIg [i vor exprima in schimb al]i markeri de suprafa] ('plasma cell antigens'): PCA1 [i PCA2.
Celulele de memorie sunt celule de repaos, ins se activeaz rapid dup orice reantalnire cu Atg. specific reluand ciclul de mai sus (Fig.28 [i 29).
Fig.27. Veriga finala a imunitatii umorale: sinteza anticorpilor de catre plasmocite
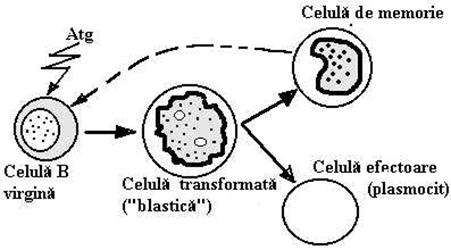
Fig. 28 Etapa dependentei de Atg.
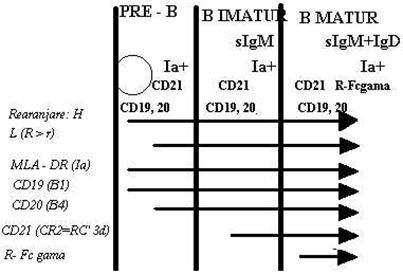
Fig. 29 Exprimarea markerilor in
cursul diferentierii limfocitelor B
2.3. Ontogeneza limfocitelor T.
a) Etapa timica. Celulele stem migrate din maduva osoasa ('protimocitele') se dispun in zona corticala unde vin in contact cu celulele epiteliale ale stromei si cu celulele fagocitice (dendritice) localizate in timus. Toate aceste celule secreta factori stimulatori ('hormoni timici'- timo-poietina, timulina, timozina, factorul timic seric- care sunt secretati de celulele epiteliale si IL1 care este produsa de celulele dendritice) si exprima antigenele condificate de CMH. Sub influenta acestor molecule protimocitele se vor diferentia ireversibil in celulele T. Procesul diferentierii implica doua serii de evenimente care se intrepatrund:
- maturatia si diversificarea in subseturi (subpopulatii) functionale;
- dobandirea tolerantei fata de self (educatia LfT).
Ca si in cazul LfB, maturatia LfT conduce in final la exprimarea pe suprafata a receprorului specific pentru Atg. (Ti) si a altor molecule functionale care asigura 'imunocompetenta'. Pe masura ce se matureaza, Lf intratimice ('timocitele') migreaza din corticala timica in medulara. De-a lungul acestui parcurs ele vor trece prin trei stadii de diferentiere (Fig.30)
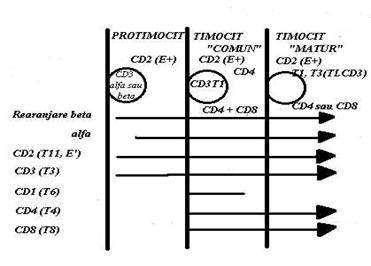
Fig. 30 Exprimarea markerilor in cursul diferentierii limfocitelor T
Stadiul I ('protimocitele' corticale) este initiat de exprimarea pe suprafata, a receptorului pentru eritrocitele de oaie (CD2 sau T 11)si de sinteza de molecule CD 3 care raman localizate intracitoplasmatic. Incepe rearanjarea genelor pentru lanturile beta si ceva mai tarziu aceea a lanturilor alfa ale receprorului pentru Atg. (Ti). Aceste lanturi vor ramane in acest stadiu cantonate in citoplasma.
Stadiul II ( timocitul 'comun'): pe suprafata celulara apar inca trei Atg.: CD 1 ( sau T6), un Atg. efemer care se exprima numai in acest stadiu, CD 4 si CD 8 (care apar ca coexprimate pe aceeasi celula).
Stadiul III ( timocitul 'matur'). Timocitele migreaza in medulara si segrega in doua grupe: celulele CD4+ si celulele CD8+.
Paralel cu aceasta selectie apar pe suprafata trei molecule noi: Ti, CD3 si CD Din acest moment timocitele au ajuns sa completeze exprimarea pe suprafata, a tuturor moleculelor implicate in recunoastere si activare: receptorul pentru Atg. (Ti) care se asociaza fizic cu CD3 (si formeaza complexul de recunoastere Ti-T3), moleculele CD4 si CD8 (exprimate fiecare pe cate un subset specific de LfT) si moleculele CD2 (care se mentin pe suprafata LfT). Devenite deci, imunocompetente cele doua subseturi de LfT (CD4+ si CD8+) parasesc timusul si se distribuie in zonele T-dependente al OLP. Etapa timica de la celulele stem pana la eliberarea LfT mature dureaza circa 3 zile.
Activarea LfT, mediata de moleculele de suprafata semnalate mai sus, implica doua mecanisme:
- activarea specifica de Atg si cuplata respectiv cu recunoasterea moleculelor codificate de CMH. Limfocitele T pot recunoaste Atg. numai daca acesta este expus pe o suprafata celulara si asociat in mod obligatoriu cu moleculele CMH. Pentru acest model de recunoastere, LfT opereaza cu doua grupe de molecule de suprafata: complexul Ti-T3 si moleculele de legare CD4 si CD Interactiunea LfT cu celulele care expun Atg. ar incepe prin formarea unor legaturi intre moleculele CD4 si CD8 cu moleculele CMH expuse pe celula partenera (perechile de molecule partenere sunt CD4 (CMH) clasa a II-a si CD8 (CMH) clasa I; memotehnic 4x2=8 si 8x1= Ulterior receptorul Ti, recunoaste Atg., iar semnalul de recunoastere este amplificat si transmis in interiroul LfT prin intermediul lui CD3.
In acest sistem, CD4 si CD8 joaca un rol asociativ in recunoasterea Atg. care este indeplinita specific de catre Ti;
- activarea independenta de Atg. si de moleculele CMH este mediata de CD2. Aceasta molecula a fost semnalata ca o proteina care mediaza formarea rozetelor de eritrocite de oaie dar efectul de activare a LfT si formarea rozetelor sunt independente, deoarece sunt mijlocite de epitopi diferiti. Activarea prin CD2 este independenta de complexul macromolecular Ti-T3 si ar functiona ca o cale alternativa de recrutare nelegata de Atg. si care ar avea rol de augmentare (amplificare) nespecifica a reactiei imune moleculare. Activarea prin CD2 este controlata de receptorul Ti: dupa ce o celula a fost stimulata prin intermediul acestuia, activarea nespecifica prin CD2 este blocata. Activarea prin CD2 poate fi indusa in vitro cu AcMo speciali, dar 'triggerii', care actioneaza in vivo, nu au fost inca identificati. Deoarece moleculele CD2 apar inca in stadiul I al diferentierii intratimice, calea alternativa de activare poate actiona in etapele dinaintea exprimarii complexului Ti-T3 si ar servi la colectarea si eliminarea clonelor LfT autoreactive.
Comportamentul LfT vis-a-vis de Atg. si de proteinele codificate in CMH prezinta urmatoarele particularitati:
recunosc si raspund daca li se prezinta:
un Atg. strain asociat cu proteinele codificate de CMH-ul propriu (proteinele CMH);
proteinele CMH non-self in absenta antigenului, ca de exemplu in situatia allogrefelor (respingerea grefelor s-ar datora faptului ca proteinele CMH caracteristice tesutului transplantat se comporta si apar la nivelul molecular ca un Atg. strain);
nu reactioneaza impotriva
proteinelor selfului codificate in CMH, in absenta Atg. ('tolereaza') selful;
Atg. straine prezentate impreuna cu proteinele CMH non-self.
Rolul timusului in ontogeneza LfT ar avea deci o dubla valenta:
Ø organ de diferentiere si maturare;
Ø organ de educare a LfT pentru recunoasterea selfului.
Deoarece in cursul maturizarii intratimice, majoritatea celulelor limfoide prolifereaza intens si sunt lizate in numar foarte mare, s-a propus ca procesul denumit 'educare' ar consta in eliminarea selectiva a celulelor care reactioneaza impotriva selfului (antigenele codificate in CMH, asociate eventual cu alte Atg. proprii). Aceasta reactie care ar determina proliferarea intensa a clonelor autoreactive ar duce in final la epuizarea ('erodarea') lor. In acest proces, celulele destinate sa recunoasca Atg. straine nu sunt eliminate in aceasta faza deoarece nu au aproape deloc sanse sa intalneasca aceste antigene. Ele isi perfectioneaza maturarea, trec in sistemul limfoid periferic, unde vor stabili contactele cu Atg. straine. Rolul major al educarii l-ar juca celulele stromei timice care exprima Atg. CMH proprii si cu care timocitele in curs de maturare stabilesc contacte intercelulare stranse. In esenta mecanismele discriminarii selfului nu sunt descifrate. Se presupune ca LfT poseda nativ un repertoriu de posibilitati de a produce molecule de receptori pentru Atg. straine si pentru o gama larga de Atg.CMH (si proprii si straine). Epiteliul timic ar selectiona numai acele celule limfoide care poseda receptori pentru moleculele CMH autohtone exprimate pe suprafata celulelor epiteliale. Desi, este logic, modelul presupus nu explica paradoxul comportarii LfT mature: cu toate ca sunt tolerante fata de moleculele CMH-self poseda un aparat de receptie prin care se recunoasc Atg. straine numai daca sunt asociate cu Atg. CMH proprii.
b) Etapa posttimica (dependenta de Atg.)
Subpopulatiile T mature au fost grupate in doua mari categorii cu functii distincte:
celule cu functii efectoare
Tc = T. citotoxice, implicate in distrugerea celulelor infectate cu virus a celulelor tumorale si in respingerea grefelor.
celule cu functii de reglare
Th = T helper sau auxiliare, au rol esential in stimularea aparitiei plasmocitelor si activarea celulelor Tc si Ts.
Ts =T supresoare, moduleaza raspunsul imun umoral si celular in sens limitativ.
LfTh sunt T4+ (exprima markerul CD4). Populatia CD4+ cuprinde, insa. pe langa celulele Th si limfocitele T 'inductoare'. Acestea favorizeaza selectia celulelor corespunzatoare Atg. si induc maturarea LfTc si Ts.
La fel populatia CD8+ include atat LfTc cat si LfTs. O subpopulatie aparte ar fi aceea a LfTHD implicate in reactiile de hipersensibilitate intarziata.
In plus celulele Ts sunt expuse si ele unui sistem de reglare negativa exercitat de un subset auxiliar de celule contrasupresoare (Tcs) care au capacitatea de a elibera LfB de sub influenta negativa a celulelor Ts.
LfT in repaos exprima pe suprafata, moleculele implicate in cele doua cai de activare: CD2 si complexul Ti-T3. Numarul moleculelor Ti-T3 este maxim. In schimb receptorul pentru IL2 (RIL2) este absent sau slab exprimat.
Dupa receptia semnalului eliberat de Atg. are loc urmatoarea secventa de evenimente celulare:
internalizarea ('modularea') complexului Ti-T3.
exprimarea rapida a RIL2 pe suprafata celulara.
secretia IL2
diviziunea celulara
In inducerea fenomenelor initiale este necesara interventia Il1 eliberata de Mcf.
Reamintim calea alternativa de activare prin CD2 care este independenta de IL1. Rolul acestuia ramane de precizat. Dupa incetarea stimularii prin Atg., receptorii Ti-T3 revin pe suprafata paralel cu disparitia RIL2.
2.4. Metode de identificare a celulelor limfoide
Principalele seturi de elemente limfoide
Ø B: Secretia de Atc. (efectorii de RI umoral)
Ø T (cu mai multe subpopulatii:
- Tc - citotoxice (elemente efectoare ale RI celular)
- Th - auxiliare ('helper')
- Ts - supresoare elemente ale reglarii RI
Ø LGL (large granular lymphocytes) - asigura functia NK ('natural killer'): citotoxicitate directa
Ø K ('Killer') - celule mononucleate asemanatoare cu Lf care indeplinesc functia de citotoxicitate mediata cu Atc. (ADCC = antibody dependent cellular cytotoxicity).
a) Identificarea prin anticorpii monoclonali (AcMo)
In prezent s-a elaborat un numar extrem de mare de AcMo care servesc la identificarea multor tipuri de Atg.celulare. Pentru evitarea confuziilor generate de diferentele de terminologie dintre acei AcMo produsi de firme diferite dar care recunosc epitopi comuni, s-a introdus nomenclatura 'CD' (Cluster Designation) pentru definirea markerilor.
Markerii au semnificatii diferite. Unii sunt exprimati pe toate celulele unei serii celulare si in toate treptele de maturatie ale acestor celule ('panantigene'). Ele servesc pentru diagnosticul acelei serii (ex. CD2 exprimat de LfT; CD24 apare pe LfB). Alte Atg. sunt exprimate numai intr-o anumita treapta a maturatiei celulelor unei serii, permitand identificarea etapei de maturatie sau a 'varstei' celulare (ex. CD1 este exprimat pe LfT imature din timus, iar CD3 pe celulele T adulte). Exista si markeri care definesc subseturi functionale ale unei populatii. CD4 este markerul LfT helper: CD8 este markerul LfT supresoare. Multi markeri au putut fi asociati cu o stare functionala a celulei (ex. CD25 reprezinta receptorul pentru interleukina 2 care apare dupa activarea LfTh). Exista si markeri comuni pentru celulele ce apartin unei serii celulare diferite: CD35 este exprimat pe LfB si T dar si pe monocite, neutrofile, eozinofile sau eritrocite si joaca rol de receptor (R) pentru componenta C3 a complementului.
Exemplele citate arata ca pentru unii dintre markeri s-a descifrat rolul functional. Ei pot fi receptori pentru semnalele diferite din mediu. CD3 este strans asociat cu R pentru Atg al LfT (RACT) impreuna cu care participa la recunoasterea Atg. CD4 si CD8 participa la procesul de recunoastere a Atg. ca molecule de legatura. Celulele prezentatoare de Atg.exprima pe suprafata Atg cuplat cu molecule ale complexului major de histocompatibilitate. LfT exprima pe suprafata lor receptorul pentru Atg. (denumit conventional Ti)
impreuna cu moleculele CD3 cu care formeaza un complex functional de recunoastere (TiT3). (Fig.31)
Pe suprafata LfTh complexele TiT3 sunt exprimate alaturi de moleculele CD4, in timp ce LfTs exprima complexul TiT3 alaturi de CD
Recunoasterea Atg. este conditionata de legaturile care se creaza intre aceste 4 molecule partenere. CD4 se asociaza cu molecule CMH de clasa a II-a, iar CD8 cu moleculele CMH de clasa I-a.
Aceste asocieri apropie celulele, creand conditiile 'analizarii' Atg. de catre Ti. Daca semnalul antigenic este acceptat, el va fi transmis cu ajutorul CD3 in celula limfoida, provocand activarea acesteia. Daca semnalul Atg. nu este recunoscut de Ti, cele doua celule se desprind urmand sa-si caute alte partenere (Fig.32).
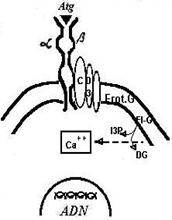
Fig.31 Ti-T3 ( Ti-CD3): unitatea
func]ional de recunoa[tere a Atg. de ctre limfocitele T. Ti este
receptorul pentru Atg. Semnalul receptat este preluat de CD3 [i prin intermediu unor relee
proteo-enzimatice membranare ( proteina G, fosfolipaza C) provoac
generarea mesagerilor secundari (inositol trifosfatul [i diacil
glicerolul). I3P [i DC induc cre[terea
Ca+ intracelular. Anumite proteine
specifice transportatoare de Ca+ de exemplu Calmodulina) mediaz n continuare activarea unor gene
nucleare care comand activarea
celulei ( exprimarea R.IL 2, secre]ia IL2, proliferare)
Receptorii sunt determinati genetic. Impreuna cu ceilalti
markeri (a caror semnificatie functionala nu o cunostem) alcatuiesc fenotipul
specific pe baza caruia pot fi identificate celulele limfoide.
b)
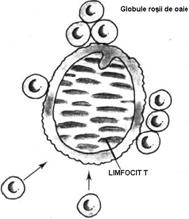
Identificarea LfT [i B prin metoda de rozetare Eritrocitele
de oaie incubate impreun cu LfT se ata[eaz formand un complex celular in
care LfT apare inconjurat de Er. (rozet). Ata[area este mediat de dou
molecule: CD2 (Th) de pe suprafa]a LfT (care prezint unele similitudini
structurale cu sIg) [i LFA 3 de pe memebrana eritrocitelor de oaie. LFA 3
(Lymphocyte Function Associated Antigen) este o glicoprotein din familia
mare a proteinelor de adeziune [i deci difer de Atg. eritrocitare 'de grup sanguin'. Aceste rozete
au fost denumite 'directe' sau 'E' (Fig.33.)
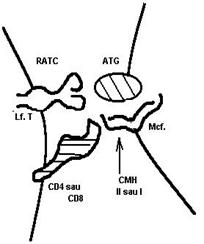
Fig.32 Interactiuni moleculare
care se stabilesc intre Mcf si LfT
cu ocazia prezentarii Atg.
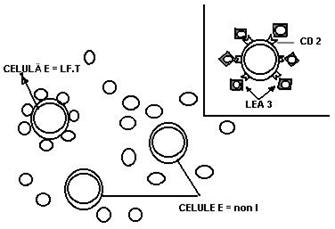
Fig. 33 Formarea rozetelor cu Er. de oaie
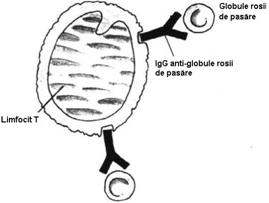
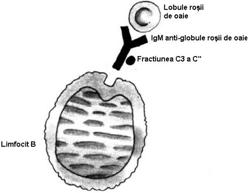
LfB exprima pe suprafata R pentru fragmentul Fc al IgG si R pentru C3b. Ele ataseaza pe acesti receptori eritrocite invelite cu IgG (rozete 'EA'- erytrocyt-antibody) si respectiv, eritrocite invelite in molecule de complement care au fost fixate prin intermediul unor Atc. IgM, care servesc ca piese de legatura (rozete 'EAC' - erytrocyte-antibody-complement). (Fig.34).
Prin
urmare dintr-o cohorta de celule limfoide neidentificate cele care vor fi EA+
si EAC+ apartin populatiei B.
Fig. 34 Rozete 'EA' si 'EAC
Fig. 35 Principiul formrii rozetelor EAC, utilizate pentru studiul receptorilor pentru C'
In
sangele periferic circula 1500 -3000 limfocite/mmc. Dintre acestea aproximativ
15-20% sunt limfocite B (EACH) iar aproximativ 80% sunt limfocite T
(E+) cca 60% sunt LFTh si aproximativ 30% LfTs. Raportul normal
Th/Ts este aproximativ 2. Circa 1-2% dintre limfocitele circulante sunt E-
si EAC- ('non-B non-T').
Printre aceste celule 'nule'
unele demonstreaza activitate NK si K.
|
Fenotipul LfB mature |
|
- sIg (Ig de suprafata) = R pentru Atg: serveste la recunoasterea Atg - RFc = R pentru segmentul Fc al IgG - CMH.DR = Atg de histocompatibilitate de clasa a II-a - CD35 (CR1) = R.C3b - CD21 (CR2) = R.C3b - M (mouse) = R pentru eritrocite de soarece: este exprimat pe LfB incomplet mature - CD19 = pan B. - CD20 = pan B. |
|
Fenotipul LfT mature |
|
- YT35 (Ti) = R pentru Atg al LfT - CD3 (T3) = indica maturarea, este asociat functional cu Ti - CD2 (T11) = R pentru eritrocite de oaie - CD25 (Tac) = R pentru interleukina 2 (IL2) - CD4 (T4) = este exprimat numai pe LfT helper si inductoare (Th/i) - CD8 (T8) = este exprimat numai pe LfT citotoxice si supresoare (Tc/s) - CMH.DR = este exprimat numai de LfT activate |
c) Identificarea LfB si T dupa receptorii pentru Atg.
Receptorul pentru Atg. al LfB este sIg. sIg. este exprimata pe LfB mature si poate fi identificata prin tehnici de imunofluorescenta. Se prepara un ser anti-Ig umana prin imunizarea specifica a unui animal. Atc. anti-Ig umana din acest antiser se concentreaza, se purifica si apoi se cupleaza cu fluoresceina. Dupa incubarea cu LfB, anticorpii marcati se ataseaza pe sIg. Preparatele se examineaza la un microscop cu sursa de UV. Pe fondul intunecat al campului microscopic celulele B apar inconjurate de un nimb stralucitor de culoare verzuie. Cu metode similare de marcare a unor AcMo s-a putut observa ca LfB imature ('celulele pre-B') nu exprima receptorii imunoglobulinici pe suprafata, dar prezinta in schimb Ig intracitoplasmatica (cIg). Prin urmare, decelarea cIg si sIg permite aprecierea maturarii celulei (a 'varstei') si aparteneta la populatia B.
Pe suprafata a peste 95% dintre LfT exista un receptor specific pentru Atg. (RACT sau Ti) format din 2 lanturi alfa si beta de marime inegala. Structura primara a acestor lanturi amintesc de aceea a lanturilor H si L din care sunt formate moleculele de Ig: toate contin o portiune variabila (V) unde este plasat situsul de combinare cu Atg. si o portiune constanta (C). Celelalte 5% dintre LfT exprima un receptor diferit, format prin combinarea unor lanturi gama si delta. Ca si sIg, RACT reactioneaza specific cu Atg. Specificitatea este asigurata de diferentele extrem de mari ale secventei de aminoacizi din portiunea variabila a moleculelor receptorilor. O celula matura exprima pe suprafata receptori cu o singura specificitate. O astfel de celula este membra a unei 'clone' (o familie de celule identice care provin dintr-un precursor comun).
Prin urmare, organismul contine o multitudine de clone dotate fiecare cu specificitate reactiva pentru un Atg. dat. Se admite ideea ca ar trebui sa existe tot atatea clone cate antigene. Aceasta ar insemna construirea unui numar imens de modele moleculare de receptori pentru Atg. Variabilitatea structurala a receptorilor este comandata genetic.
2.5. Macrofagele (Mcf)
Mai multe metode experimentale au demonstrat existenta unei cooperari intre Mcf si Lf in raspunsul imun. Intr-unul dintre acestea s-a lucrat cu o suspensie pura de Lf (din care s-au inlaturat Mcf) preparata de la un soarece imunizat in prealabil cu un Atg. (in acest experiment s-a folosit complexul gamaglobulina + dinitrofenol) ( gamaglobulina DNP).
Fig 36. Macrofagul - microscopie electronic `n baleiaj
a) incubarea Lf sensibilizate cu gamaglobulina-DNP nu este urmata de secretie de Atc.
b) amestecul Lf + gamaglobulina-DNP + Mcf. provenite de la soarecii singenici neimunizati in prealabil cu gamaglobulina-DNP, produce un raspuns anticorpic moderat.
c) Lf + gamaglobulina-DNP + Mcf. preincubate cu gamaglobulina produce un raspuns energic.
d) Mcf. care au fagocitat Atg. il pot transfera la un animal nesensibilizat.
Mcf. sunt necesare pentru declansarea raspunsului imun. Mcf fagociteaza Atg., dar il degradeaza doar partial mentinand intacti determinantii (epitopii) pe care-i va expune pe suprafata 'oferindu-i' pentru recunoastere, limfocitelor (fenomenul 'prezentarii' Atg).( Fig.37)
Captarea Atg. de catre Mcf. ar fi deci prima treapta necesara pentru declansarea raspunsului imun. Dupa administrarea unui Atg. in circulatie, se observa disparitia lui treptata (fenomen denumit 'clearance' prin care Atg. este captat de Mcf., care-l transporta in organele limfoide). Fenomenul de clearance a fost demonstrat prin administrarea de flagelina de Salmonella Adelaide marcata cu I125. Autoradiografierea organelor limfoide a evidentiat Atg. legat selectiv de macrofagele migrate in aceste organe. Mcf. provin din precursori care se diferentiaza si se matureaza in M.O. Celulele maturate in MO sunt monocite. Acestea sunt eliberate in circulatie de unde migreaza in diverse tesuturi unde-si vor indeplini functiile. Mcf. sunt monocite stabilite in diverse tesuturi, unde indeplinesc doua activitati: fagocitoza si secretie de substante functionale solubile. In raspunsul imun sunt implicate mai multe asemenea celule ale sistemului macrofagic: Mcf. tisulare, celulele Kupffer ( in ficat), celulele Langerhans (in piele), celulele dendritice (in foliculii limfoizi), celulele interdigitate (in zonele paracorticale, T-dependente din ganglionii limfatici). Unele sunt specializate numai in functie de prezentare a Atg.: celulele Langerhans, celulele dendritice si interdigitate.
Altele intervin numai ca elemente fagocitante: celulele Kupffer care sechestreaza si fagociteaza eritrocite invelite cu anticorpi si molecule de complement. In fine, Mcf. tisulare indeplinesc ambele activitati: de prezentare a Atg. si de eliminare prin fagocitoza a resturilor conflictului imun.
Mcf. activate secreta substante solubile denumite generic monokine, dintre care cea mai importanta este 'interleukina'1 (IL1). IL1 este capabila sa produca activarea nespecifica a altor celule aflate in proximitatea zonei in care a patruns Atg.: Mcf.,LfT si B. Se produce o activare in lant care mareste eficienta raspunsului local. In plus, IL1 are si efecte generale: febra, anorexie, somnolenta.
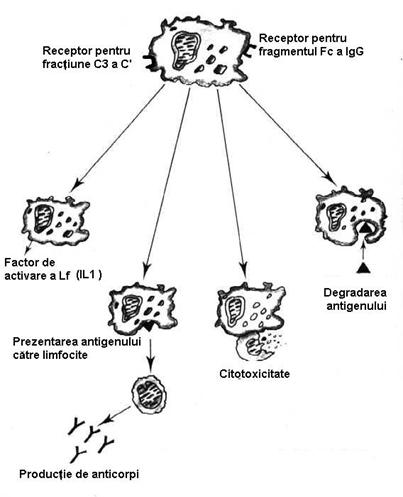
Fig.37. Prezentarea schematica a functiilor macrofagelor
In afara de IL1 Mcf activate secreta si alte substante capabile sa moduleze secretia imuna: o varianta de interferon, constituenti ai sistemului complement (C1, C3, C4, factorul B) , prostaglandinele, TNF alfa ('tumor necrosis factor alfa' sau casectina). TNF alfa induce febra, stimuleaza secretia unor limfokine,activeaza Mcf. si poseda capacitatea de liza a unor celule tumorale.
Mcf. poseda pe suprafata receptori pentru C' si pentru segmentul Fc al moleculelor de Atc. Cu ajutorul acestor receptori capteaza Atg. pe care s-au atasat C' si/sau Atc (imunoaderenta). Gratie imunoaderentei Mcf. elimina Atg. din organism.
Mcf. intervin in doua momente ale reactiei imune prin valorificarea functiilor lor principale: fagocitoza si secretia de substante active (IL1,etc.).
Cu ajutorul fagocitozei, Mcf. intervin atat la polul aferent (inductor) al raspunsului imun (ca celule prezentatoare de Atg.), cat si ulterior, la polul eferent (efector) al raspunsului cand elimina Atg. din organism.
Exista o relatie dubla intre Mcf. si Lf : prin oferta de Atg. si prin secretia de interleukine (care creaza in plus efecte reciproce de activare). Astfel, ca raspuns la IL1, LfTh secreta o substanta stimulatoare (IL2) care produce activarea Mcf, a LfB si a diverselor subpopulatii de LfT (Th, Ts, Tc).
Modul general de reactie al limfocitelor, ca raspuns la o activare, este secretia unor substante care mediaza rolul acestor celule in aparare, LfB secreta Atc. care se ataseaza pe Atg., permitand macrofagelor sa elimine aceste Atg. prin intermediul fenomenelor de imunoaderenta. LfT secreta alte tipuri de molecule (limfokinele) LfTc care vin in contact cu celulele straine sau infectate, secreta moleculele toxice cu ajutorul carora ucid celulele-tinta (de ex. limfotoxina sau 'tumor necrosis factor' beta, o substanta inrudita din punct de vedere structural cu TNF alfa produs de Mcf.) - LfTh isi indeplinesc multiplele lor functii de reglare prin intermediul interleukinelor. Acestea joaca rolul unor adevarati hormoni ai mecanismelor imunitare.
2.6. Celulele K
Tintele celulare cele mai variate, invelite prin concentratii scazute de anticorpi IgG, devin sensibile la efectul citotoxic al anumitor celule mononucleate, celulele K (de la engl. killer). Aceasta este citotoxicitatea celulara dependenta de anticorpi (ADCC- 'antibody dependent cell-mediated cytotoxicity').(Fig.38)
Nu exista o categorie unica, ci numeroase tipuri de celule sunt capabile de a exercita o activitate ADCC. Ca o caracteristica comuna aceste celule poseda pe suprafata lor receptori de inalta afinitate pentru fragmentul Fc (FcR) al IgG, dupa legarea de acestia devenind capabile sa exercite o activitate citotoxica.
Nu exista o categorie unica, ci numeroase tipuri de celule sunt capabile de a exercita o activitate ADCC. Ca o caracteristica comuna aceste celule poseda pe suprafata lor receptori de inalta afinitate pentru fragmentul Fc (FcR) al IgG, dupa legarea de acestia devenind capabile sa exercite o activitate citotoxica. Celulele cele mai bine cunoscute pentru activitatea lor ADCC sunt polinuclearele, macrofagele, plachetele sangvine, celulele ficatului fetal si mai ales celulele mononucleare, prezente in organele limfoide lipsite de markeri si proprietati ale celulelor B si T carora de obicei li se rezerva termenul de celule K. Natura si mecanismele de actiune al acestor celule K nu sunt fundamental diferite de cele ale altor celule capabile de ADCC.
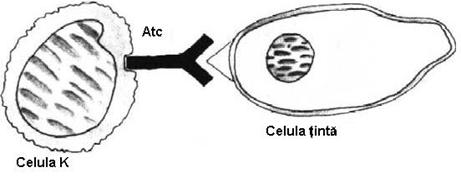
Fig. 38 Schematizarea fenomenului de citotoxicitate celulara
dependenta de Atc
Definitia celulelor K este in mod esential negativa. Celulele K nu prezinta Ig de membrana si sunt lipsite de markeri ai celulelor T (in particular receptorii de rozetare E si antigenele de diferentiere recunoscute prin anticorpii monoclonali). Acest fenotip de celule nule le apropie de efectorii NK. In ultima instanta nu exista markeri specifici care sa permita distingerea de o maniera viabila a celulelor K si NK. Experientele functionale de competitie au demonstrat ca anumite celule nule prezinta simultan o activitate K si NK, in timp ce altele sunt fie K, fie NK.
2.7. Celulele NK
Anumite celule mononucleate prezinta o activitate citolitica aparent nespecifica unui antigen, dirijata contra anumitor celule-tinta, in general reprezentate de linii de celule tumorale, independent de o sensibilizare prealabila si de restrictia CMH. Celulele in chestiune sunt denumite ucigase naturale prin analogie cu anticorpii naturali produsi fara imunizare intentionata (NK pentru 'natural killer').
Linia celulara la care apartin celulele NK ramane inca obiect de dezbatere. Celulele NK pot fi reprezentate de o noua familie de celule limfoide, derivate din maduva osoasa, fie de un stadiu particular de diferentiere celulara proprie uneia sau mai multor linii hematopoietice. Ele se regasesc in organele limfoide (cu exceptia timusului) si in sange.
Atat la soarece cat si la om, celulele exprima antigenele de diferentiere proprii celulelor limfoide.
Mai multe argumente sugereaza legatura cu linia celulara T: prezenta anumitor markeri ai celulelor T (antigene Thy -1, de densitate mica la soarece, fixarea anumitor anticorpi monoclonali anti-T si formarea de rozete E la om: sensibilitatea la interleukina 2). Prezenta unui nivel crescut de activitate NK la soarece ca si modularea prin hormonii timici, sugereaza ca celulele NK par sa reprezinte celulele T intr-o faza imatura a diferentierii lor in timus.
Celulele NK se disting de monocite prin absenta fixarii anticorpilor Mo 2 si LeuM1, de celulele B prin absenta imunoglobulinelor de suprafata si a receptorilor pentru complement si de celulele T mature prin absenta anumitor antigene (Lyt 2 la soarece, CD3, CD4 si CD6 la om). Ele se apropie totusi de celulele T prin prezenta pe o proportie importanta dintre ele, a antigenelor Lyt1 si Thy-1 la soarece, CD8, CD11, Leu5 si CD10 la om si de celulele fagocitare prin reactivitatea lor cu anticorpii OKM1 ca si cu anticorpii dirijati contra receptorilor Fc.
Anumite limfocite mari granulate de 16-20 m si care asigura in mod esential activitatea NK au fost observate in sangele circulant la om. Aceste celule purtatoare de granule citoplasmatice azurofile, care reprezinta lizozomi sau filamente de ectina poseda un nucleu reniform.
Ele pot fi purificate prin centrifugare in fractiuni mai usoare. Aproximativ jumatate dintre ele formeaza rozete E si sunt recunoscute prin diversi anticorpi monoclonali specifici celulelor T, prin anticorpi OKM1 (specific monocitelor) si Leu7 dar nu prin OKT3. Ele sunt in egala masura purtatoare de receptori Fc si sunt inzestrate cu activitate citotoxica ADCC. Aceste limfocite la care par sa existe numeroase subpopulatii, sunt regasite intr-un nivel ridicat in sange (7%), intr-un procentaj mai scazut in splina (4%), in ganglioni (1%) si in epiteliile mucoase. Ele nu se gasesc in timus.
Interferonii de toate originile, cresc foarte evident activitatea NK, la toate speciile testate. IL-2 stimuleaza deasemenea, activitatea NK si poate determina aparitia unei activitati citotoxice de tip NK in celule CTL (celulele T citotoxice) autentice.
Asemenea celule care si-au pierdut stricta lor specificitate de CTL sunt denumite 'Lymphokine Activated killer' (LAK). pe de alta parte celulele NK sunt sensibilizate de actiunea mecanismelor supresoare in care intervin celulele T. Nivelul activitatii este controlat prin factorii genetici, din care unii sunt asociati CMH.
3. DINAMICA RASPUNSULUI IMUN
Imunitatea este definita ca ansamblul fenomenelor prin care organismul recunoaste si neutralizeaza structuri celulare sau necelulare agresoare, care sunt straine de organismul respectiv.
Raspunsul imun are 3 caractere fundamentale:
este xenofob ( anti-non-self), implicand o discriminare a selfului (totalitatea constituientilor proprii) de non-self (molecule, macromolecule sau celule straine de organismul respectiv).
Reamintim ca selful este definit de moleculele CMH pe care LfT sunt 'educate' sa le recunoasca. Moleculele CMH de clasa I (exprimate pe toate celulele nucleate) permit limfocitelor sa recunoasca celulele proprii si sa le deosebeasca de structurile impotriva carora trebuie sa reactioneze:
agresori exogeni care nu poseda Atg. CMH ( de ex, microbi);
molecule CMH non-self, exprimate pe celule straine (ca in cazul grefelor);
molecule CMH proprii modificate (self alterat), exprimate pe celule proprii infestate cu virus sau transformate malign.
este specific: raspunsul imun este declansat numai daca o substanta straina se uneste cu receptorii pentru Atg. exprimati de Lf si numai daca exista o potrivire structurala intre Atg. si situsul combinativ al receptorului (specificitate inseamna receptor pentru antigen sau un tip de receptor/o celula, sau o clona de celule pentru un anumit Atg). Imunitatea stabilita impotriva unui agent strain se exercita numai impotriva acelui agent.
este memorizat: reantalnirea cu un Atg. fata de care organismul a fost imunizat in prealabil va declansa un raspuns mai precoce si mai intens ( raspuns secundar sau 'anamnestic').
Exista doua modele de raspuns imun (RI):
A. RI cu mediere umorala
declansat in conditiile in care Atg. au reusit sa se raspandeasca in organism;
se desfasoara departe de locul unde au patruns Atg;
este mediat de Atc. circulanti care in vivo, demonstreaza capacitatea de neutralizare a toxinelor, opsonizarea bacteriilor, inactivitatea virusurilor;
este activat in infectii bacteriene, in reinfectiile virale;
raspunsul aberant duce la reactia de hipersensibilitate imediata (anafilactica).
B. RI cu mediere celulara
declansat in general de Atg. care raman cantonate la poarta de intrare (ex. grefelor) sau care sunt fixate in celule ale gazdei (agenti patogeni cu dezvoltare intracelulara ca: virusuri, fungi B.K., Brucella, Atg. tumorale);
se desfasoara la locul unde este cantonat Atg. (de ex. la poarta de intrare).
se constutie lent ('raspunsul intarziat');
este indreptat in special impotriva unor celule (straine, tumorale, infestate cu agenti patogeni) care exprima Atg.;
este mediat de Lf. cit. specifice, sensibilizate care actioneaza asupra celulelor-tinta direct sau asistate de producerea unor mediatori solubili.
Derularea unui raspuns imun poate fi sistematizata in urmatoarele etape:
intalnirea cu Atg.
activarea Lf specifice;
eliminarea Atg.
supresia raspunsului;
Intalnirea cu Atg.
Antigenele patrund in organism strabatand barierele naturale care separa organismul de mediul inconjurator dintre care cele mai importante sunt pielea si mucoasele care tapiseaza arborele respirator si tractul digestiv. La acest nivel Atg. sunt interceptate de mecanismele de aparare nespecifice, sunt captate de celulele sistemului monocitar-macrofagic si dirijate catre ganglionii regionali sau catre alte structuri ale sistemului organelor limfoide periferice (OLP) care sunt intercalate in acest circuit (amigdale si alte struturi Waldeyeriene, placi Peyer, etc.) Atg care depasesc bariera limfatica si patrund in sange (sau cele care accidental, intra direct in sange), interactioneaza in plasma cu anticorpii specifici preexistenti (rezultati in urma unui raspuns anterior fata de Atg. dat.) si sunt vehiculate spre splina. In OLP, Atg. sunt retinute de macrofage si de celulele dendritice si interdigitate. OLP sunt sediile in care se declanseaza raspunsul imun. Aici Atg. persista un timp suficient de lung ( zile sau luni) pentru a putea fi inglobate, prelucrate si reexpuse pe suprafata celulelor dendritice si interdigitate, conditie necesara pentru a putea fi prezentate limfocitelor. Pe suprafata acestor celule, determinantii antigenici sunt reexpusi in asociere cu proteinele CMH de clasa I-a si clasa II-a.
Se pare ca, intocmai ca macrofagele, si limfocitele B sunt capabile sa inglobeze sa prelucreze si sa reexpuna pe suprafata Atg., impreuna cu moleculele CMH de clasa II-a, putand servi in anumite conditii, ca celule prezentatoare de Atg.
Recunoasterea Atg. de catre Lf este conditionata de particularitatile functionale ale receptorilor specifici pentru Atg:
receptorul pentru Atg. al LfB (sIg) poate recunoaste Atg. ca atare;
receptorul pentru Atg. a LfT (Ti) poate recunoaste Atg. numai daca este prelucrat si asociat cu proteine CMH. Restrictia CMH pentru recunoasterea Atg. este diferentiata: celulele CD4+ recunosc Atg. asociat cu proteinele CMH de clasa II-a (CMHDR) (aceste proteine sunt prezente numai pe celule care coopereaza in raspunsul imun: macrofagele, LfB si unele celule T activate);
Celulele CD8+ recunosc Atg. asociat cu proteinele CMH de clasa I-a (virtual prezente pe toate celulele nucleate).
Activarea celulelor specifice.
Activarea LfTh (CD4+). Ocupa locul central al raspunsului imun. Initial LfTh primeste doua semnale de activare de la macrofagul care-i prezinta Atg., semnale pe care le asimileaza cu ajutorul a trei molecule receptoare.
|
Cooperare Mcf - LfT.CD4 |
|
|
Semnale Il1 Atg + CMH II |
Molecule receptoare R.IL1 Ti + CD4 |
Receptia acestor semnale provoaca activarea limfocitelor, care este urmata de secretia de IL2, care se adauga ca al treilea semnal de autoactivare a celulei respective (stimularea autocrina). IL2 se deverseaza in mediul inconjurator unde se asociaza altor actiuni al celulelor CD4+ activate. Activarea LfTh are mai multe consecinte functionale:
1. Proliferarea clonala a LfT auxiliare (cu producere de celule fiice activate care secreta diverse limfokine cu efecte stimulatoare) si diferentierea unor subseturi de celule CD4+ capabile sa induca rapid capacitatea T cit. de a raspunde la Atg. (limfocitele T inductoare) (Fig.39).
Proliferarea si activarea T cit.;
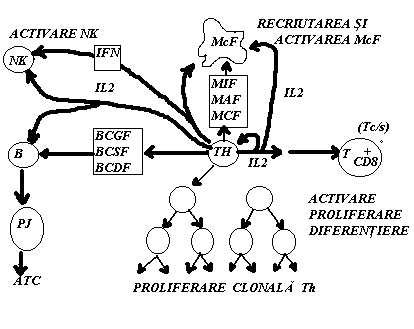
Fig. 39 Activarea limfocitelor T-helper
3. Activarea reciproca a Mcf. prin IL2: Mcf. activate secreta pe langa IL1, IFN si prostaglandine;
4. Recrutarea de noi Mcf. in focarul de recunoastere a Atg. si activarea lor prin efectul unor limfokine: MIF (factorul de inhibitie a migratiei macrofagelor), MCF (factorul chemotactic pentru excitarea Mcf.). MAF (factorul de activare a Mcf);
5. Stimularea LfB specifice la Atg., declansata prin doua semnale: interrelatia celulara directa T-B si IL2.;
6. Amplificarea activarii, a proliferarii si a transformarii LfB, in plasmocite secretoare prin intermediul mai multor limfokine: IL2, factorul de stimulare, factorul de proliferare, factorul de diferentiere;
7. Inducerea maturarii si activarii celulelor Ts si activarea prin IFN a celulelor NK.
Activarea LfB
Selectarea LfB este initiata de atasarea Atg. pe sIg. Aceasta legatura nu este capabila sa provoace singura activarea LfB specific decat in cazul Atg. timo-independente (molecule mari-polimerice, cu multipli determinanti identici si dispusi in serie sub forma de unitati repetitive care se leaga concomitent pe mai multi receptori sIg declansand o sumatie de semnale care depasesc pragul de activare a Lf.).
In majoritatea cazurilor insa, activarea LfB este timo-dependenta. Atg. sufera degradarea limitata urmata de reexpunerea epitopilor in conjunctie cu molecule CMH II atat in Mcf. cat si in LfB preselectate. Captarea Atg. de catre Mcf. este nespecifica. In schimb, Atg. va fi endocitat si prelucrat numai de acele LfB care exprima molecule de sIg specifice pentru Atg. in cauza. Complexele Atg.+ CMH II exprimate pe suprafata Mcf si a LfB se leaga pe cuplurile de molecule partenere pe care LfTh le exprima pe membrana (Ti+CD4).
Se concepe realizarea unui model de cooperare intre 3 celule: Mcf - Th si Th - B. In cooperarea Mcf - Th recunoasterea CMH II de catre CD4 permite transmiterea semnalului antigenic de la Mcf. la LfTh prin intermediul R.Ti. In cooperarea Th - B interrelatii simultane CD4 + CMH II si Ti + Atg. fixeaza cele doua celule si elibereaza un semnal helper care induce excitarea LfB. Acest semnal se insumeaza cu primul semnal produs prin cuplarea Atg. pe sIg. si se asociaza cu efecte stimulatoare ale IL2 si a altor interleukine emise de LfT. In acest model Atg. serveste ca o punte de legatura intre celulele partenere iar raspunsul este specific deoarece este conditionat de recunoasterea Atg. de catre receptorii specifici ai celor doua celule.
Activarea LfB se soldeaza cu transformarea acestora in plasmocite care vor secreta Atc. cu aceeasi specificitate ca aceea a sIg. Aceasta similitudine asigura reactivitatea Atc. cu Atg.
Functia principala a Atc. este legarea sa pe Atg. Atc. fixati pe Atg. contribuie la eliminarea acestora prin intermediul asa-numitei functii efectoare a Atc.: atasarea componentei C1q a complementului urmata de activarea cascadei prin calea clasica si aderarea lor la celule care poseda receptori pentru segmentul Fc (Mcf. si celulele K). Imunoaderenta la Mcf. este urmata de fagocitarea si digestia complexelor Atg-Atc., indiferent ca Atg. sunt corpusculare (celulare) sau solubile (macromolecule).(Fig.40)
Atasarea complexelor formate din celule-tinta + Atc. la R.Fc exprimati de celulele K produce un efect citotoxic exercitat de acestea din urma asupra celulei-tinta (ADCC= antibody- dependent cell mediated cytotoxicity). ADCC ar fi o reactie de aparare indreptata impotriva celulelor tumorale sau asupra unor celule proprii alterate.
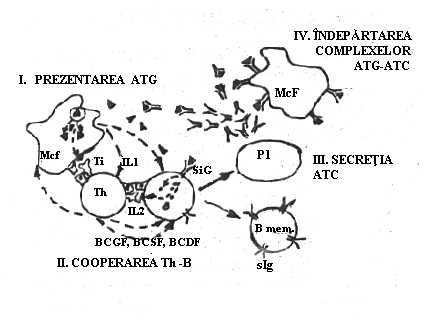
Fig. 40 Raspunsul B Timodependent
Activarea LfT cit.
Necesita doua semnale. Semnalul specific este eliberat de Atg. expus impreuna cu molecula CMH de clasa I pe o suprafata celulara prezentatoare si este receptionat de celulele T cit. prin intermediul complexului Ti+CD Activarea mediata pe aceasta cale este specifica. Al 2-lea semnal este furnizat de IL2 secretata de LFTh prin mecanismul descris mai sus. Tintele LfT cit. sunt celule infectate cu virus sau celule straine care sunt distruse printr-un efect citotoxic direct.
Spre deosebire de citoliza mediata de celulele K si NK citoliza produsa de LfT cit., este foarte specifica, gratie dublei recunoasteri a tintelor. Din acest motiv LfT cit. nu recunosc celulele care exprima numai antigenele specifice in absenta moleculelor CMH ca de ex. hematiile sau bacteriile. Recent s-au izolat cateva cazuri care se caracterizeaza prin deficienta formarii (sau exprimarii) Atg.CMH ('sindromul limfocitelor dezgolite'). Aceasta se traduce clinic printr-o imunodeficienta grava cu infectii severe si repetate. (Fig.41)
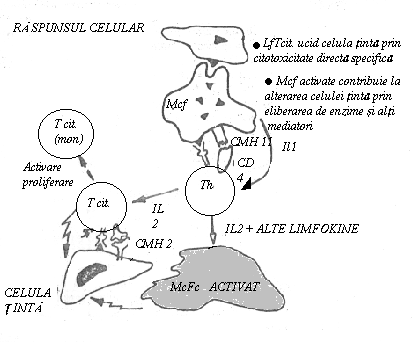
Activarea LfTs se produce, cronologic ca un eveniment tardiv al lantului activarilor B si T. Aceasta permite desfasurarea raspunsului imun. Au fost descrise 2 tipuri de celule Ts: Ts specifice pentru Atg. care reactioneaza numai cu Atg. corespunzator, suprimand raspunsul imun exclusiv fata de acesta si Ts nespecific pentru Atg. care suprimeaza raspunsul imun in general si care ar avea specificitate pentru receptorii LfT si B ( si deci si prin idiotipurile acestora). Maturarea lor este stimulata de LfT4 inductoare iar activarea lor ar fi dependenta de un helper al celulelor T4 auxiliare. In dinamica raspunsului normal celulele Th stimulate de Atg. ar emite mai intai semnalele de diferentiere pentru LfB si mult mai tarziu pe cele de producere a celulelor Ts. Cand acestea ajung intr-un numar suficient de mare, induc o inhibitie retroactiva a celulelor auxiliare care le-a stimulat, realizandu-se un circuit imunologic de reglare. Efectul supresor ar putea fi mediat de un factor solubil supresor.
Memoria imunologica
Este procesul biologic datorita careia, la orice reintalnire cu un Atg., raspunsul imun se instaleaza mai rapid si este mai energic (raspunsul secundar sau 'anamnestic').
Ambele populatii de Lf (B si T) produc celule de memorie. Tratarea soarecilor cu ser anti-B respectiv anti-T suprima capacitatea acestora de a determina un raspuns secundar in vitro si in vivo. LfT de memorie sunt activate mai rapid decat celulele virgine in prezenta Atg. specifice. Activarea este urmata de conversia lor mai rapida in celule efectoare.
Dupa ce s-au format, celulele de memorie parasesc locurile lor de nastere (splina, ganglioni, etc.) si patrund in circulatie ca celule de viata lunga (luni, ani) care 'patruleaza' (recircula) pe traiectul vaselor de sange - tesuturi - vase limfatice - ganglioni - conexiuni limfatice interganglionare - duct toracic - sange.
Unele din ele patrund in ganglionii limfatici prin arteriolele aferente si ajunse in venule post capilare strabat peretii acestora trecand astfel in tesutul limfatic ganglionar.
Mecanismul formarii celulelor de memorie nu este bine descifrat. Dupa una din ipoteze, Lf de memorie s-ar forma printr-o diferentiere asimetrica a celulelor activate de Atg.: dupa mitoza, una din celulele fiice se transforma in celula efectoare iar cealalta ramane ca limfocit mic de memorie. Aceasta s-ar produce paralel cu expansiunea clonei respective.
3.1. Reglarea genetica a raspunsului imun
Exista animale care raspund viguros la unele Atg. si altele incapabile sa produca Atc dupa stimularea cu Atg. respective.
Fenomenul a fost studiat de BENACERRAF si MC DEVIT pe suse de soareci cu ajutorul unor substante antigenice simple care contin cate un singur determinant. S-au utilizat soareci din doua rase (A si B) cu puii de prima generatie rezultati din incrucisarea lor, (hibrizii F1). Soarecii A si B raspund in mod diferit (+ sau -) la diversele Atg. Hibrizii F1 raspund la Atg. recunoascute de oricare dintre cei doi parinti. In concluzie, aptitudinea soarecilor de a raspunde la Atg. este controlata de gene dominante. Raspunsul este mediat de limfocite: Lf recoltate de la animale reactante transfera capacitatea de raspuns la animale nonreactante preparate prin iradiere letala. Genele respective au fost denumite 'Ir'('Immune response').
Genele Ir ar fi plasate in subregiunea CMH care codifica proteinele de clasa II-a. Produsele genelor Ir ('immune response associated antigens') sunt exprimate pe aceleasi celule care exprima Atg.HLA de clasa II-a. Drept urmare ele ar putea fi implicate in procesele de recunoastere a Atg.
Date fiind, analogiile observate intre CMH murin si uman se admite ca prealabila existenta a genelor Ir si la om. Acestea ar fi plasate in regiunea D si ar putea fi chiar gene DR.
Genele raspunsului umoral. Raspunsul anticorpic al soarecilor dupa administrarea unor antigene (eritrocite de oaie) este diferit: exista soareci care raspund printr-o mare secretie de Atc. si altii la care se inregistreaza raspunsuri slabe. Selectia riguroasa a acestor animale a demonstrat ca aceasta capacitate de raspunsuri ample este ereditara si nespecifica: raspunsurile sunt ample atat vis-a-vis de Atg. inductoare cat si dupa administrarea altor Atg. T-dependente si T-independente. Genele care controleaza intensitatea raspunsului umoral sunt diferente de genele Ir. Localizarea lor nu este inca stabilita.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 6653
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved