| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Ciclul lizogen
Ciclul infectios lizogen este cunoscut, in primul rand, pentru cuplul fag l si celulele de E. coli (fig. 100).
Fagul l se deosebeste de fagii din seria T (par si impar), atat din punct de vedere morfologic, cat si al interactiunii sale cu celulele sensibile. Capul este un icozaedru aproape perfect, cu diametrul de 55 nm, iar coada este mai lunga (l55 nm), mai subtire si mai flexibila si se ingusteaza progresiv spre extremitatea libera. Este goala in interior, necontractila, formata din 35 de discuri suprapuse.
Genomul fagic este o molecula de ADN dublu catenara, lineara si se termina cu secvente monocatenare, complementare, denumite cozi adezive sau coezive. In interiorul celulei bacteriene, molecula se circularizeaza devenind dublu catenara, circulara inchisa covalent, sub actiunea catalitica a unei polinucleotid-ligaze.
Genomul fagului l cuprinde circa 50 de gene. Cele din jumatatea stanga (a moleculei lineare) codifica proteine cu rol morfogenetic, iar cele din jumatatea dreapta codifica factorii de interactiune ai genomului fagic cu cromosomul bacterian, fiind importante in evolutia lizogena a cuplului fag-bacterie.
Corespunzator celor doua programe genetice, cuplul fag - E. coli are doua cai posibile de evolutie :
calea litica, ce implica sinteza proteinelor virale si morfogeneza fagului, urmata de liza celulei bacteriene;
calea lizogena, in cursul careia, replicarea genomului fagic este stopata. Genomul fagic se circularizeaza prin legarea capetelor sale adezive si se integreaza ca profag in cromosomul bacterian.
Fagii care dupa infectie nu lizeaza celula se numesc temperati, iar o celula bacteriana care contine un set de gene fagice se numeste lizogena. ADN fagic se gaseste fie integrat in cromosomul bacterian, fie in stare autonoma (fizic independenta).
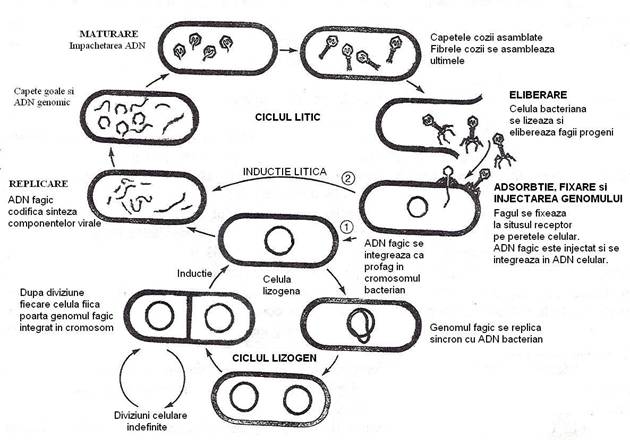
Fig 100. Dupa infectie, cuplul E. coli - fag λ poate evolua litic sau lizogen.
Dintre celulele infectate de fagii temperati, numai o proportie mica evolueaza lizogen, marea majoritate fiind lizate.
Fagul l virulent, inoculat pe o panza de celule bacteriene sensibile formeaza o plaja de liza clara, deoarece toate celulele sunt lizate. Fagul l temperat formeaza o plaja de liza cu o zona centrala turbida (opalescenta). Turbiditatea este determinata de multiplicarea bacteriilor lizogene, imune fata de fag.
Decizia intre liza si lizogenie este data de un mecanism reglator, complex si subtil, influentat de structura genetica a fagului, a celulei si de factorii de mediu: concentratia nutrientilor in mediu si multiplicitatea de infectie. La o multiplicitate scazuta de infectie se desfasoara preponderent ciclul litic. Dar dupa cateva cicluri litice, multiplicitatea de infectie devine inalta si un numar mic de celule sunt lizogenizate.
Odata cu epuizarea nutrientilor din mediul agarizat, celulele bacteriene inceteaza sa se divida, virusul nu se mai multiplica si plaja inceteaza sa se extinda. Deoarece in aria plajei, nutrientii sunt inca disponibili, datorita lizei timpurii a majoritatii celulelor, cele lizogene continua sa creasca si sa se divida, rezultand plaje cu un centru turbid.
Oricare ar fi ponderea acestor factori, lizogenia este consecinta blocarii transcrierii si traducerii informatiei genetice a fagului, a blocarii ciclului sau de multiplicare, urmata de integrarea genomului ca profag, in cromosomul bacterian.
Integrarea genomului fagului l in cromosomul bacterian este rezultatul recombinarii intre situsurile specifice de legare att (attchment, englez = atasare), situate pe cromosomul bacterian si pe cel fagic. Ele conditioneaza afinitatea de legare a celor doua genomuri.
Situsurile att bacterian si fagic au secvente omologe de baze si sunt alcatuite din doua jumatati situate simetric fata de o regiune mediana O, de circa l5 baze, in care omologia celor doua genomuri este totala. Situsul fagic de legare are o secventa de 3l7 baze si este notat POP' (Phage), iar cel al cromosomului bacterian are o secventa de 250 de baze si este notat cu BOB' (Bacterium). Localizarea sa a fost identificata intre gena gal (codifica enzimele metabolizarii galactozei) si gena bio (codifica sinteza biotinei). Recunoasterea celor doua genomuri se face prin intermediul regiunii O, la care participa secventele PP' si BB'.
Esenta mecanismului de integrare este circularizarea cromosomului fagic. Pentru ca integrarea genomului fagic sa aiba loc, cele doua genomuri sunt sectionate la nivelul situsului de legare, sub actiunea unei enzime fagice din setul timpuriu o terminaza cu functie de endonucleaza, denumita integraza, o enzima ce recunoaste situsurile de legare a ADN fagic si ADN bacterian si catalizeaza legarea recombinatorie a celor doua molecule.
In procesul de integrare, zona centrala de omologie este cea care determina si conditioneaza integrarea, iar zonele laterale au un rol secundar in procesul de legare recombinatorie a celor doua genomuri.
Integraza este o topoizomeraza I. Ea catalizeaza clivarea unei catene a dublului helix, roteste un capat al catenei clivate in jurul catenei intregi, sectioneaza si cea de a II-a catena si apoi reuneste extremitatile libere. Catenele sectionate ale celor doi cromosomi, sunt asezate intr-o continuitate perfecta.
Ca rezultat al integrarii, ordinea lineara a genelor in molecula de ADN fagic este permutata. Apar doua noi situsuri de legare: BOP' si POB'.
La sfarsitul procesului de integrare, cromosomul bacterian este mai lung cu echivalentul unui genom fagic, fapt demonstrat prin conjugare, prin masurarea intervalului de timp necesar transferului celor doua gene (gal si bio) in procesul conjugarii.
Integrarea fagului l in cromosomul bacterian corespunde unui proces unic, denumit recombinare integrativa sau recombinare la situsuri specifice (fig. 101). Omologia dintre situsurile de legare a celor doua genomuri, nu este perfecta ca in recombinarea generala, dar procesul este rezultatul actiunii unor enzime care recunosc anumite situsuri specifice si sectioneaza decalat secvente relativ omologe ale celor doua genomuri, generand extremitati adezive, care permit legarea incrucisata si integrarea genomului viral ca profag.
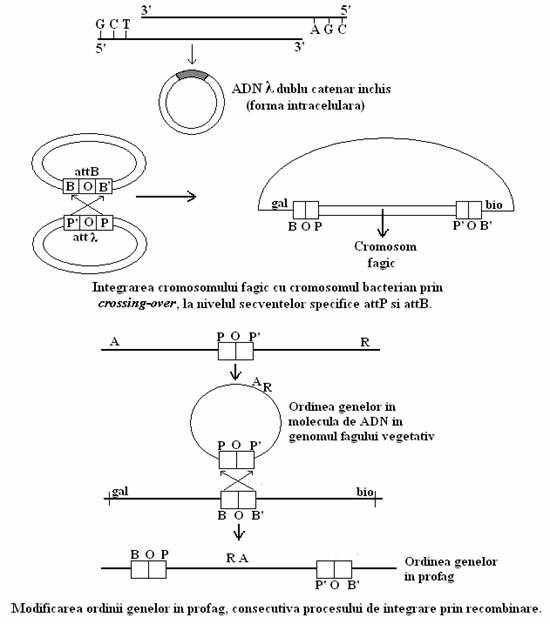
Fig. 101. Reprezentarea schematica a etapelor procesului de integrare a cromosomului fagic λ in cromosomul bacterian.
Dupa integrare, genele fagice se comporta ca si genele cromosomale: se replica sincron cu cromosomul bacterian si se transmit prin diviziune celulara, generatiilor succesive, ca si genele cromosomale.
Starea lizogena este controlata de represorul l, o proteina de 200 aminoacizi, codificata de profag, care se leaga de regiunea operator a genomului fagic integrat. Astfel, represorul fagic, intra in competitie cu ARN-polimeraza, care, pentru initierea transcrierii se leaga de acelasi situs al profagului. Legarea prompta a represorului l de situsul de legare a operonilor virali are ca efect blocarea transcrierii tuturor genelor fagice, cu exceptia celor care codifica sinteza represorului.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4729
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved