| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Mutatiile
Mutatia este o modificare stabila a materialului genetic, transmisibila la descendenti. Molecula de ADN este o tinta cu situsuri multiple pentru actiunea factorilor lezionali endogeni si exogeni. Nucleotidele din incizura majora a dublului helix, sunt mai sensibile la modificare decat cele din incizura minora. Unii dintre atomii bazelor purinice si pirimidinice sunt mai sensibili decat altii. Din aceste considerente, molecula de ADN este o tinta heterogena pentru actiunea agentilor lezionali.
Mutatiile sunt produse prin modificari stabile ale secventei de baze ale unei gene, transmisibile la descendenti. Ele pot fi punctiforme, adica implica modificarea unei singure baze si pot altera situsul de actiune al unui agent antimicrobian, interferand cu activitatea lui sau modifica o secventa de baze.
Mutatiile modifica totdeauna genotipul, dar in functie de natura lor pot sa nu fie exprimate in fenotip.
In functie de agentul cauzal, mutatiile sunt induse de factori endogeni (denumite si mutatii spontane) si de factori exogeni (denumite si mutatii induse).
Expresia fenotipica a unei mutatii este precedata totdeauna, de aparitia unor modificari lezionale ale moleculei de ADN, care scapa actiunii corective a mecanismelor reparatorii.
Mutatiile spontane se produc in absenta actiunii oricarui agent, cunoscut ca avand efecte asupra moleculei de ADN. Diferite activitati metabolice, in special anaerobe, pot sa genereze intermediari cu efecte lezionale fata de ADN. Cei mai cunoscuti sunt internediarii reducerii O2 (H2O2, radicalul O2-, radicalul OH.) Generarea intermediarilor care lezeaza ADN, in metabolismul anaerob, este argumentata de inducerea enzimelor de reparare a ADN in conditii de anaerobioza.
Mutatiile spontane sunt punctiforme. Ele apar in timpul replicarii ADN si par a fi datorate erorilor de imperechere a bazelor intre catena nou sintetizata si cea veche.
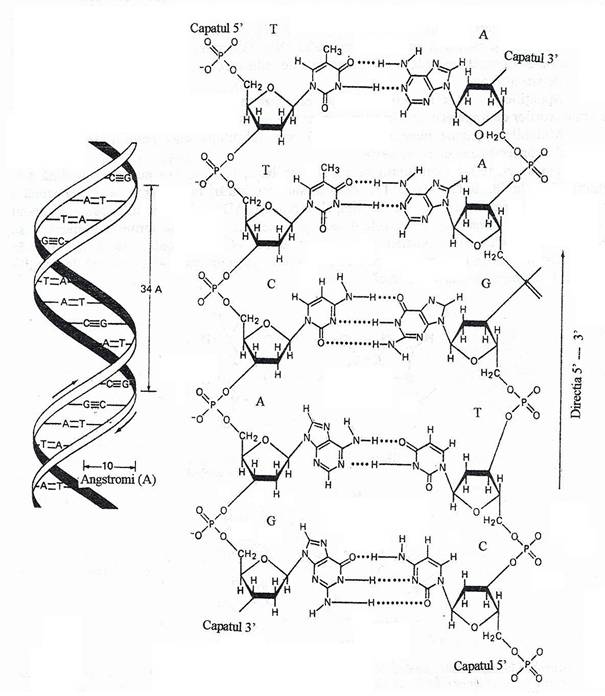
Fig. 113. Modelul structural al moleculei de ADN care evidentiaza configuratia dublu helicala si raporturile celor doua catene nucleotidice. In stanga, reprezentarea diagramatica a moleculei. Catenele asemanatoare unor panglici reprezinta axul central al moleculei de ADN. Cele doua catene sunt antiparalele. Este ilustrata imperecherea bazelor, legaturile de H si dimensiunea moleculei de ADN. Un tur de helix cuprinde o pereche de baze. In dreapta, o scurta secventa a moleculei de ADN care arata raportul dezoxiribozei si acidului fosforic cu bazele.
Mutatiile spontane punctiforme rezulta prin substitutia unei perechi de baze si se datoreaza unor fenomene de transversie.
Tranzitiile sunt mutatii punctiforme care apar prin substitutii de perechi de baze, in care o baza purinica (A sau G) este substituita cu o alta purina, sau o pirimidina (C sau T) este substituita cu o alta pirimidina. Perechea purina - pirimidina este schimbata de la GC la CC sau la TA. Acestea sunt modificari rare si apara ca o consecinta a imperecherilor gresite ce au loc in timpul replicarii ADN.
Transversiile sunt mutatii punctiforme in care o baza purinica este substituita de o baza pirimidinica sau invers si se datoreaza fenomenului de tautomerie. Cele 4 baze pot coexista in diferite forme tautomere ce se deosebesc prin schimbarea pozitiei unui proton: grupul -C=O (cetonic) este transformat in grupul -C=OH (enol), iar gruparea -NH2 este transformata in gruparea -NH (imina). In conditii normale, predomina gruparile cetonica si aminica (fig. 114).
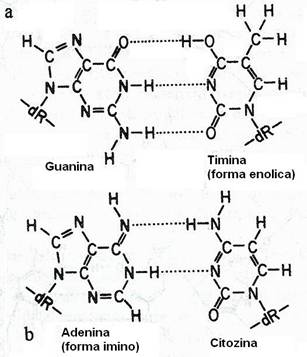
Fig. 114. Schimbari in imperecherea bazelor datorate starilor tautomerice. In forma enolica (a), timina formeaza legaturi de H cu guanina, in locul adeninei. In forma imino (b), adenina formeaza legaturi de H cu citozina, in locul timinei. Modificari similare ale guaninei si citozinei determina modificari ale imperecherii bazelor.
Aparitia gruparilor tautomere (enol si imino) produce o relaxare a imperecherii bazelor si permit guaninei sa formeze pereche cu timina (in loc de citozina), pastrand perechea purina-pirimidina. Daca perechea GT nu este corectata la replicare, unul dintre cromosomi va avea perechea normala AT (in loc de GC) si astfel s-a produs o mutatie punctiforma.
Depurinarea este consecinta sensibilitatii inalte la hidroliza, a legaturilor glicozidice care leaga guanina in catena nucleotidica, dar celelalte, care leaga adenina sau bazele pirimidinice (citozina, timina) sunt mai stabile. Situsurile apurinice astfel rezultate (si mai rar apirimidinice) sunt recunoscute de sistemele de supraveghere si reparare.
In cursul replicarii ADN, uneori, nucleotidele sunt incorporate gresit. Perechile incorecte pot scapa functiei de control (editing) a ADN-polimerazei.
Disponibilitatea limitata a dUTP sau inhibitia sistemului enzimatic al timidilat-sintetazei, are ca rezultat inhibitia sintezei dTTP si incorporarea uracilului in locul timinei. Prezenta uracilului este recunoscuta de sistemele de reparare.
O21, O2-, radicalul OH (OH.) liber produc leziuni la nivelul restului glucidic, a caror consecinta poate fi ruperea catenei ADN.
Mutatiile induse sunt rezultatul leziunilor produse de actiunea factorilor exogeni asupra moleculei de ADN si apar cu o rata net superioara fata de nivelul ratei mutatiilor spontane.
Factorii mutageni exogeni sunt agentii chimici si radiatiile.
Agentii chimici cu efect mutagen actioneaza la nivel molecular prin alterarea secventei de baze a ADN: analogi ai bazelor, agenti alchilanti, agenti de dezaminare, derivati ai acridinei.
Un analog al unei baze este o molecula similara ca structura, cu una dintre bazele azotate ale ADN. O celula poate incorpora in ADN, un analog al bazei, in locul bazei normale. De exemplu, 5-bromo-uracilul (5-BU) poate fi incorporat in locul timinei.
Analogii bazelor induc substitutia unei baze prin fenomenul de tranzitie. Tranzitia rezulta prin inlocuirea unei purine cu o alta purina, sau a unei pirimidine cu alta pirimidina, adica o schimbare de la perechea GC la AT sau invers.
5-BU si analogii sai tautomerizeaza mai usor decat bazele naturale. In forma -C=O, 5-BU se imperecheaza cu A si este incorporat in ADN, formand o pereche pseudo-AT. In forma -C=OH, la urmatorul rund de replicare, 5-BU se imperecheaza cu G, in locul lui A. La un nou rund de replicare, tranzitia de la AT la GC se va stabiliza.
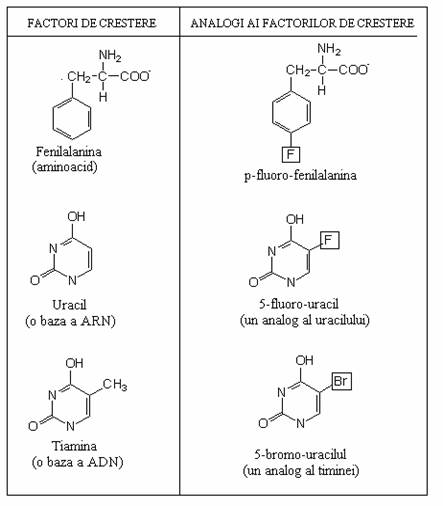
Fig. 115. Exemple de analogi ai factorilor de crestere.
Agentii alchilanti* sunt substante care inlocuie H cu o alta grupare (de exemplu, -CH3 sau C2H5), intr-o baza, ca de exemplu guanina. Adaugarea un grup -CH3 la o baza azotata, ii modifica dimensiunile si determina imperecherea eronata. Agentii alchilanti actioneaza in special asupra ciclurilor purinice azotate, la nivelul O6 al guaninei sau O4 din bazele pirimidinice, producand leziuni mutagene, dar si la nivelul legaturilor fosfodiesterice ale catenei de ADN. Guanina metilata se imperecheaza gresit cu Timina, in locul Citozinei. Astfel, la urmatoarea runda de replicare a ADN, G initiala este inlocuita cu A, care va face pereche cu cu T.
Sinteza si transferul gruparii -CH3 este un proces metabolic important. Atomii C,O,N,S ai compusilor organici functioneaza frecvent ca acceptori ai gruparii -CH3 in procese metabolice primare si secundare. Procesul poarta denumirea de "biometilare". Termenul de "bioalchilare" semnifica procesul prin care o grupare alchil se leaga direct de un element cu numar atomic mai mare de 10. Termenul de bioalchilare presupune acceptarea altor grupari alchil: etil, propil, desi procesul are o frecventa mica.
Agentii alchilanti se formeaza prin prepararea multor produse alimentare, in gazele de esapament prin combustia interna a N2 atmosferic, formadu-se nitrati si nitriti. Prin arderea tutunului, a produselor petroliere si prin prepararea alimentelor, din resturile de guanina se formeaza hidrocarburi policiclice aromatice, cu efect alchilant asupra ADN.
Agentii de dezaminare (de exemplu, acidul nitros) scot un grup -NH2 dintr-o baza azotata. Conversia adeninei la hipoxantina, a guaninei la xantina si a citozinei la uracil, prin dezaminare, are efecte mutagene pentru ca bazele dezaminate se imperecheaza cu alte baze decat cu cele normale. De exemplu, dezaminarea citozinei la uracil modifica perechea C=G la perechea U=A, iar dezaminarea adeninei la hipoxantina (analoga guaninei), modifica perechea A=T la G=C (fig. 116).
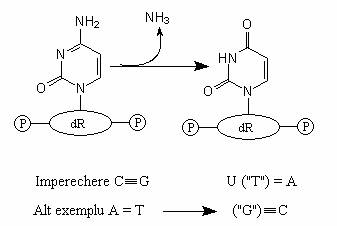
Fig. 116. Reactiile de dezaminare au consecinte mutagene, deoarece bazele dezaminate se imperecheaza gresit.
NO3- si NO2- se adauga la unele alimente (preparate din carne) pentru culoare, miros sau pentru actiune antibacteriana. In tubul digestiv se formeaza nitrozamine. Acestea sunt agenti de dezaminare care produc cancere ale tractului digestiv si efecte defavorabile asupra dezvoltarii fatului.
In contrast cu aceste alterari care produc mutatii punctiforme, derivatii acridinei, cauzeaza mutatii prin schimbarea cadrului de citire. Molecula de acridina contine un inel pirimidinic si doua inele benzenice. Molecula sau unul dintre derivatii sai se insera in dublul helix al ADN, inlocuind ambele baze ale unei perechi. Modificarea deformeaza helixul si produce despiralizarea partiala a catenelor de ADN. Despiralizarea permite ca una sau mai multe baze sa fie adaugate sau eliminate. Rezulta astfel, o mutatie prin schimbarea cadrului de citire.
Mutatiile prin schimbarea cadrului de citire, constau in insertia sau deletia uneia sau catorva baze in molecula de ADN (un numar diferit de multiplu de 3). Ele determina schimbarea cadrului de citire, pentru ca dincolo de situsul mutatiei, toti codonii isi schimba secventa de baze. A II-a schimbare a cadrului de citire, de aceiasi marime, dar in sens invers, restabileste cadrul initial de citire a informatiei genetice. O astfel de mutatie se numeste supresoare a primei mutatii si pentru ca are loc in aceiasi gena, fenomenul se numeste supresie intragenica.
Colorantii de acridina, in special proflavina, bromura de etidiu, induc mutatii ale cadrului de citire (fig. 117, 118). Proflavina este o molecula planara, cu dimensiuni similare cu ale unei perechi purina-pirimidina si se intercaleaza intre bazele ADN. Mutagenii care schimba cadrul de citire sunt agenti de intercalare, dar nu toti agentii de intercalare schimba cadrul de citire.
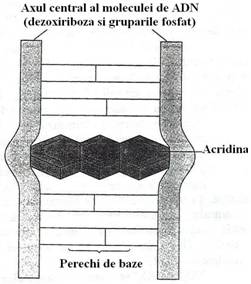
Fig. 117. Insertia acridinei in dublul helix ADN poate produceo mutatie prin schimbarea cadrului de citire.
Bromura de etidiu (colorant)
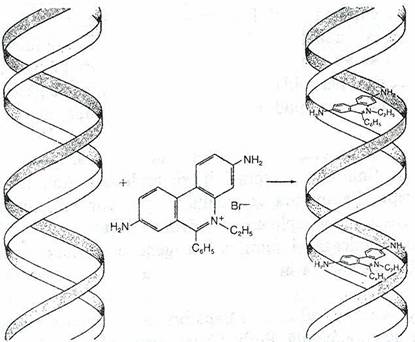
ADN-dublu catenar Moleculele de colorant intercalate intre baze confera flurescenta moleculei de ADN
Fig. 118. Bromura de etidiu se intercaleaza intre bazele ADN
(dupa Brock,1988).
Mutatiile punctiforme ale ADN (tranzitii, transversii sau cele produse prin schimbarea cadrului de citire) modifica secventa de baze in ARNm. O mutatie de sens gresit da nastere unui codon ce specifica un alt aminoacid decat cel codificat original. Efectul aminoacidului nou asupra polipeptidului, depinde de pozitia aminoacidului si de natura schimbarii. Polipeptidul poate sa ramana nemodificat, dar alteori functia sa se poate pierde complet sau mutatia poate genera o proteina cu stabilitate termica alterata. La o temperatura relativ scazuta (permisiva), proteina functioneaza normal, iar la temperaturi mai mari (nepermisive), proteina este nefunctionala (mutanta termosensibila). Daca rezultatul este moartea celulei, mutatia se numeste conditionat letala, adica este letala numai la temperatura nepermisiva.
Mutatiile punctiforme pot schimba un codon sens (care specifica un aminoacid), la un codon care specifica terminarea sintezei proteinei (UAG, UAA, UGA). Se sintetizeaza o proteina incompleta, in locul celei normale. Astfel de codoni mutanti se numesc nonsens, deoarece nu codifica incorporarea unui aminoacid.
Generarea unui codon nonsens intr-o gena proximala a unui operon, poate induce un efect polar asupra celorlalte gene ale operonului, situate distal fata de situsul codonului nonsens. Codonul nonsens semnifica stoparea traducerii ARNm si peptidul nascent este eliberat din ribosomi. Uneori, ribosomii se pot reatasa la ARNm, distal fata de situsul nonsens, la un alt situs de legare. Daca numarul ribosomilor care se reasociaza cu ARNm, este mai mic decat normal, rezulta fenomenul de polaritate a traducerii. Uneori, efectul polar asupra traducerii este puternic, adica genele situate distal fata de situsul mutatiei nonsens sunt foarte putin exprimate.
Mutatiile punctiforme sunt dependente de replicarea ADN, pentru consolidare.
Mutatiile majore produse prin modificarea unui numar mare de baze se datoreaza deletiilor, duplicatiilor, inversiilor si aditiilor. Mutatiile majore pot sa implice secvente de ADN ce depasesc marimea unei gene, iar efectele lor sunt mult mai ample decat ale mutatiilor punctiforme. Ele se datoreaza erorilor de replicare, evnimentelor de recombinare intre secventele omologe de ADN sau actiunii mecanismelor de reparare. Deletiile sunt rezultatul actiunii mecanismelor de transpozitie a elementelor genetice transpozabile (EGT).
Erorile de replicare a ADN (erori de copiere) se datoreaza alunecarii unei catene a ADN in raport cu cealalta, in timpul replicarii. Dupa ce s-au separat tranzitoriu, cele doua catene se realiniaza incorect, datorita formarii unei bucle pe catena matrita. Reluarea copierii la o secventa ce precede bucla produce o duplicatie, iar reluarea copierii in aval de bucla, determina o deletie in catena nou sintetizata.
Deletiile si inversiile pot fi rezultatul recombinarii intra- si inter-repliconi, pentru ca recombinarea necesita duplicatii de secvente. Recombinarea intre secvente omologe, cu aceiasi orientare intr-o molecula de ADN, duce la pierderea unei copii a secventei duplicate.
Radiatiile UV sunt mutagene. ADN absoarbe radiatiile cu lungimea de unda de circa 260 nm. Majoritatea celulelor bacteriene mor, dar cele care supravietuiesc sufera mutatii cu o frecventa inalta. Cea mai frecventa leziune indusa de radiatiile UV, este formarea dimerilor pirimidinici (fig. 119). Intre pirimidinele adiacente, pe aceiasi catena se formeaza legaturi covalente, astfel incat ele sunt reunite printr-un inel ciclobutan. Se formeaza in special dimeri T-T, dar si C-C sau T-C. Formarea dimerilor deformeaza molecula de ADN si interfera cu imperecherea normala a bazelor. Replicarea ADN se intrerupe la dimerii T-T si este reluata distal, la oarecare distanta. Astfel, in catena nou sintetizata, apare o discontinuitate, care, daca nu este reparata, devine letala.
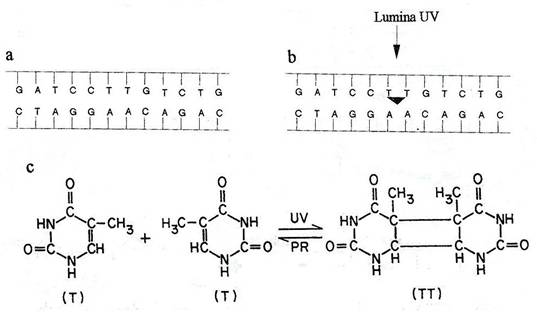
Fig. 119. Modificari induse de radiatiile UV im molecula de ADN. a. Secventa bazelor in molecula de ADN . b, c. Sub actiunea luminii UV se formeaza dimeri de timina (dupa Russell, 1998).
Radiatiile ionizante (γ si x) au energie inalta si actioneaza prin ruperea legaturilor chimice ale moleculelor. Se formeaza radicali liberi, atomi foarte reactivi, molecule sau ioni ce ataca alte molecule ale celulei, inclusiv molecula de ADN. Efectul radiatiilor ionizante consta in primul rand, in radioliza apei prin care se genereaza radicali OH (OH.). Radicalul OH. este capabil sa mobilizeze protoni de la C'4 al dezoxiribozei sau de la bazele pirimidinice. Se labilizeaza legaturile fosfodiesterice cu posibila aparitie a ruperilor mono- sau dublu catenare.
Mutatiile consecutive repararii ADN Supravietuirea bacteriilor in mediile ostile se datoreaza partial, sistemelor de supraveghere ce recunosc leziunile aparute in structura ADN si activeaza o cascada de evenimente moleculare ce restabilesc integritatea structurala si functionala a moleculei originale, asigurand stabilitatea genetica.
ADN este singura macromolecula ale carei leziuni sau defecte pot fi corectate prin actiunea unor sisteme enzimatice. Cele mai multe modificari ale moleculei de ADN sunt temporare pentru ca sunt imediat corectate sub actiunea unui set de procese denumite generic "repararea ADN". Mai putin de 1 din 1000 de schimbari ale bazelor ADN produc o mutatie permanenta, restul fiind eliminate prin procesele de reparare. Unele leziuni ale ADN (dimerii pirimidinici) nu pot fi reparate si molecula de ADN nu se replica. O celula care nu isi replica ADN, moare.
Mecanisme de reparatie genetica
La bacterii, cateva procente din capacitatea de codificare este destinata functiilor de reparare a ADN. In absenta mecanismelor de reparare, mutatiile spontane ar schimba rapid secventa de ADN. Structura dublu helicala a ADN este ideala pentru reparare. Cand una dintre catene este lezata, catena complementara pastreaza o copie intacta a informatiei si este folosita ca matrita pentru a restabili secventa initiala de nucleotide.
Fiecare celula dispune de sisteme multiple de reparare a ADN, fiecare cu propriul set de enzime. Actiunea diferitelor mecanisme este dependenta de de tipul de leziune. Cele mai multe sisteme de reparare folosesc catena intacta ca matrita pentru repararea catenei lezate.
Reparatia genetica se face prin mai multe mecanisme:
a) Reversia directa a leziunii, fara sinteza unor noi legaturi fosfodiesterice. In aceasta categorie intra fotoreactivarea si raspunsul reparator adaptativ;
b) Mecanismul reparator prin excizie si resinteza, bazat pe incizia catenei lezate la sau langa situsul leziunii, excizia si resinteza secventei excizate, folosind ca matrita, catena complementara normala;
c) Reparatia prin recombinare dupa replicare implica schimbul de catene prin recombinare;
d) Reparatia prin sistemul inductibil "SOS" a leziunilor ce nu pot fi corectate prin sistemele constitutive.
Fotoreactivarea
Data fiind frecventa mare a mutatiilor induse de radiatiile UV, celulele bacteriene si-au elaborat mecanisme cu actiune prompta pentru eliminarea leziunilor specifice si restabilirea secventei normale. Daca celulele tratate cu radiatii UV sunt expuse actiunii luminii vizibile cu lungimea de unda de 300-400 nm, atat frecventa mutatiilor, cat si letalitatea scad foarte mult. Fenomenul se numeste fotoreactivare. Iradierea UV produce o leziune caracteristica in molecula de ADN: o legatura chimica stabila, respectiv a unui dimer de tipul ciclobutil, cu patru atomi de carbon intre doua baze pirimidinice adiacente (fig. 120).
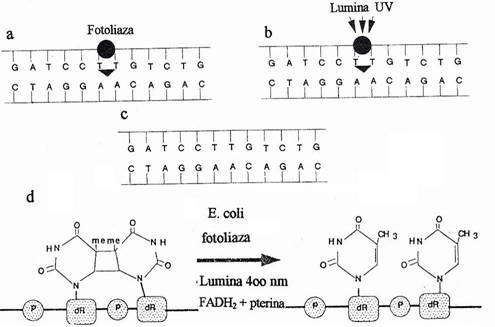
Fig. 120. Fotoreactivarea. a. O molecula de fotoliaza recunoaste dimerul si este activata de lumina cu lungimi de unda specifice. .b. Dimerul este clivat. c. Secventa ADN este restabilita. d. Reactia de fotoreactivare enzimatica a dimerilor pirimidinici de ciclobutan.
Dimerii pirimidinici se formeaza cel mai adesea (50%) intre doua resturi de T, ceva mai rar intre un rest de T si unul de C (40%) si foarte rar intre doua resturi de C (l0%). Lumina vizibila activeaza o fotoliaza, enzima ce hidrolizeaza inelul de ciclobutan si reverseaza reactia produsa de radiatia UV, fara erori, restabilind bazele monomere. Fotoreversia implica transferul de energie de la molecula de pterina, la FADH2 si transferul electronilor la dimerul pirimidinic.
Spre deosebire de mecanismul de reparare indus de lumina vizibila, care repara leziunile fara erori, celula bacteriana poseda mecanisme de reparare la intuneric, dar predispuse la erori, adica generatoare de mutatii, cu frecventa mare.
Raspunsul reparator adaptativ
Acest tip de raspuns se activeaza dupa expunerea prelungita a celulelor de E. coli, la concentratii mici de substante mutagene, ca de exemplu, N-metil-N'-nitro-nitrozoguanidina (MNNG) sau etil-metan-sulfonatul. Celulele bacteriene tratate cu concentratii mici de agenti alchilanti, care preced administrarea dozelor letale sau mutagene, devin rezistente la actiunea mutagena sau letala a unor concentratii de cateva sute de ori mai mari de substanta activa.
Raspunsul fiziologic al celulelor de E. coli, la actiunea agentilor metilanti sau etilanti s-a numit raspuns adaptativ.
Agentii metilanti sau etilanti produc leziuni ale moleculei de ADN, printr-un proces de alchilare, adica grefarea unei grupari alchil (metil, etil) pe un atom de C, N sau O (fig. 121).
Efectul MNNG consta in adaugarea unei grupari metil (-CH3), legata de un atom de O, in pozitia C6 a guaninei. Formarea leziunii O6-alchil-guanina este insotita de pierderea legaturilor de H care leaga G metilata de C de pe catena opusa.
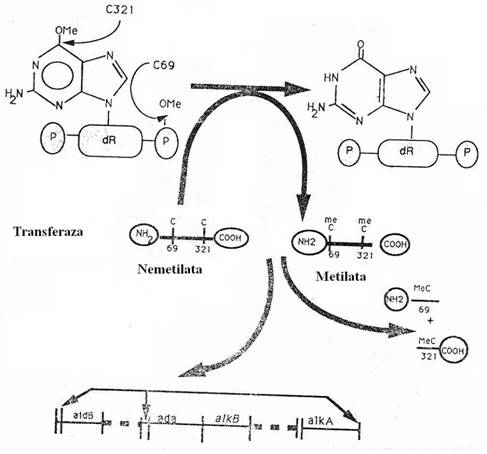
Fig. 121. Reversia leziunilor produse de agenti alchilanti la nivelul gruparilor fosfat si la nivelul guaninei. Gruparile alchil sunt transferate la resturile de cisteina ale transferazei, a carei sinteza este indusa de agentii alchilanti. Dupa metilare, enzima se inactiveaza (dupa Grossman, 1992).
Procesul
reparator este dependent de sinteza proteica de novo. Se sintetizeaza o metil-transferaza
- proteina
Repararea prin excizia bazelor, catalizata de glicozilaze si endonucleaze
Bazele modificate prin dezaminare pot fi excizate de un grup de enzime denumite ADN-glicozilaze. ADN-glicozilazele sunt enzime mici, cu specificitate inalta, foarte bine conservata in evolutie, atat ca structura, cat si ca mecanism de actiune, de la bacterii pana la om. Fiecare recunoaste un tip specific de baza alterata si catalizeaza eliminarea ei hidrolitica, prin clivarea legaturii N-glicozil a bazei cu deoxiriboza catenei. Sunt cel putin 6 tipuri de enzime: cele care elimina C dezaminata, A dezaminata, bazele alchilate sau oxidate, baze cu inele deschise sau baze la care dubla legatura C=C a fost convertita la legatura simpla C-C. Dupa excizia bazei, rezulta o deoxiriboza careia ii lipseste baza. Absenta bazei este recunoscuta de o endonucleaza AP (apurinic, apirimidinic) care cliveaza catena fosfodiesterica, leziunea este indepartata si reparata.
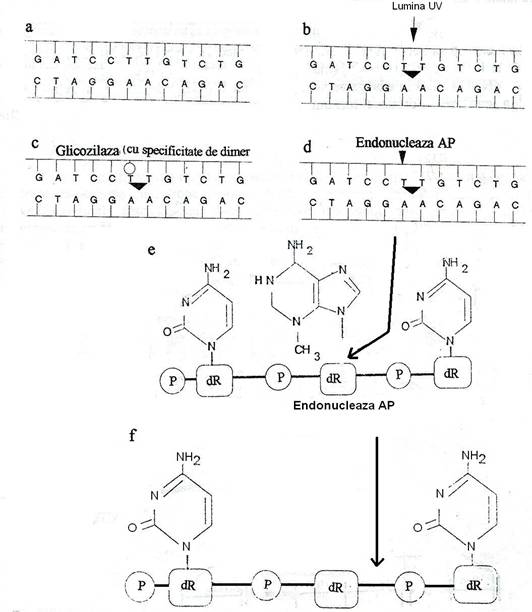
Fig. 122. Repararea prin excizia bazelor, catalizata de glicozilaze si endonucleaze. Dimerul format dupa expunerea bacteriilor la UV (a, b) este indepartat sub actiunea glicozilazei si endonucleazei AP (APurinic sau APirimidinic) specifice. Glicozilaza specifica, indusa de agentul lezional recunoaste dimerul pirimidinic si cliveaza legatura glicozidica dintre bazele lezate si dezoxiriboza, rezultand un situs apirimidinic sau apurinic. Endonucleza recunoaste situsurile AP si hidrolizeaza legaturile fosfodiesterice ale dezoxiribozei (c. d, e, f) (dupa Grossman, 1992).
Reparatia prin excizia si resinteza nucleotidelor
Mecanismul de reparatie prin excizie si resinteza, consta in excizia enzimatica a secventei lezate, urmata de completarea sa prin resinteza (fig. 123), utilizand ca matrita, catena complementara ramasa intacta. Spre deosebire de procesul fotoreactivarii, in cursul caruia dimerii de T sunt clivati in situ si rezulta monomerii corespunzatori, in reparatia la intuneric, dimerii sunt indepartati din molecula de ADN, prin excizia fragmentului in care s-au format.
Reparatia prin excizie si resinteza, a fost caracterizata la E. coli si este mediata de cel putin 6 produse genice ale sistemului uvr:
proteina UvrA, sub forma dimerica, se leaga de ADN in prezenta ATP;
ulterior, se asociaza proteina UvrB. Complexul Uvr2AB este translocat de la situsul initial al ADN, la situsul lezat, de o ATP-aza asociata cu UvrB, care este activata prin formarea complexului UvrA2B-ADN;
proteina UvrC face doua clivari in catena de ADN, la 7 nucleotide in directia 5' fata de situsul lezat si la 3-4 nucleotide in directia 3' fata de acelasi situs;
in prezenta proteinei UvrD (cu rol de helicaza), a ADN-polimerazei I si a substratului (deoxiribonucleozid-trifosfati), fragmentul lezat este eliberat, iar proteinele UvrA, UvrB, UvrC sunt recirculate.
Continuitatea catenei clivate se restabileste prin sinteza de novo, catalizata de ADN-polimeraza I, utilizand ca matrita, catena intacta, iar polinucleotid-ligaza restabileste continuitatea secventei.
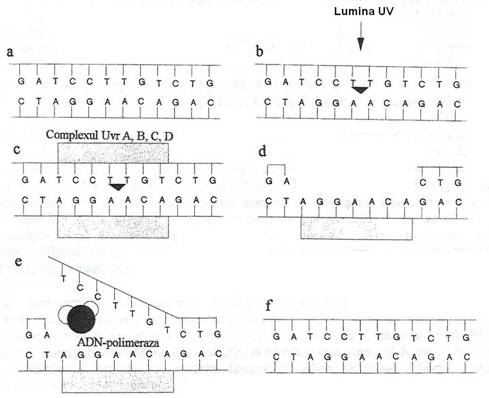
Fig. 123. Repararea nucleotidelor prin excizie si resinteza. Dimerii pirimidinici care se formeaza sub actiunea radiatiei UV sunt recunoscuti de complexul enzimatic UvrA, UvrB si UvrC (c). In prezenta UvrD, complexul cliveaza catena de ADN (d), iar AND-polimeraza resintetizeaza catena lezata, folosind ca matrita catena opusa (e, f).
Sistemul reparator inductibil "SOS"
Lezarea severa a ADN, care interfera cu capacitatea sa de replicare, determina activarea genelor care fac parte din reteaua de reglare "SOS" (fig. 124).
Activitatea reparatorie "SOS" este dependenta de produsele de sinteza a genelor rec A+ si lex A+. Cele doua gene codifica nu numai proteine reglatoare ale reparatiei ADN, ci au si alte functii:
inductia litica a unor fagi infectiosi pentru E. coli;
intarzierea diviziunii celulare, care duce uneori la formarea filamentelor etc.
Proteina Rec A are un rol esential in reparatia "SOS". Ea indeplineste doua functii:
functia de proteaza in activitatea reglatoare a sistemului;
functia de enzima de recombinare in reparatia postreplicativa.
Mutantele rec A- sunt defective pentru capacitatea de recombinare genetica in general si pentru exprimarea sistemului inductibil "SOS" si, din aceasta cauza sunt foarte sensibile la actiunea radiatiilor UV.
Substratul activitatii proteolitice a proteinei Rec A este represorul fagului lambda. Consecinta este evolutia lizogena a cuplului E. coli-fag lambda si inductia proteinei Lex A in cursul reparatiei "SOS".
Lezarea ADN intensifica sinteza proteinei Rec A. Ea trebuie sa dobandeasca activitate proteolitica, pentru a cliva proteina Lex A si represorul λ.
Functia de enzima de recombinare a proteinei Rec A, consta in stimularea formarii perechilor omologe intre doua molecule de ADN, adica de a alinia precis fragmentele de ADN care au secvente de baze complementare, pentru schimbul reciproc de baze prin recombinare.
Proteina Lex A (24 kDa), are functia de represie prin legarea de operatorul a circa l4 gene: rec A, uvr A, uvr B, uvr B, uvr D etc., precum si a propriei sale gene lex A. Astfel, proteina Lex A permite transcrierea ARNm si sinteza proteinelor codificate de aceste gene, la nivele foarte scazute in celulele neinduse. In celulele neinduse (normale), functia "SOS" este represata.
Cand celulele sunt expuse actiunii unor factori care produc leziuni importante ale ADN, ce nu pot fi reparate de nivelul scazut al enzimelor de reparatie prezente constitutiv, in molecula de ADN dublu catenara apar brese monocatenare, postreplicative. Moleculele de rec A prezente constitutiv in celula, se leaga de ADN monocatenar ramas intact in regiunea opusa breselor. Se produce un semnal inductor care activeaza functia de proteaza a proteinei rec A si odata cu aceasta, raspunsul "SOS". Proteina Rec A activata, cliveaza represorul Lex A in doua subunitati inactive si genele controlate de acest represor - uvr A, uvr B, uvr C uvr D si lex A sunt deblocate si sunt transcrise cu o rata crescuta, pentru sinteza proteinelor corespunzatoare.
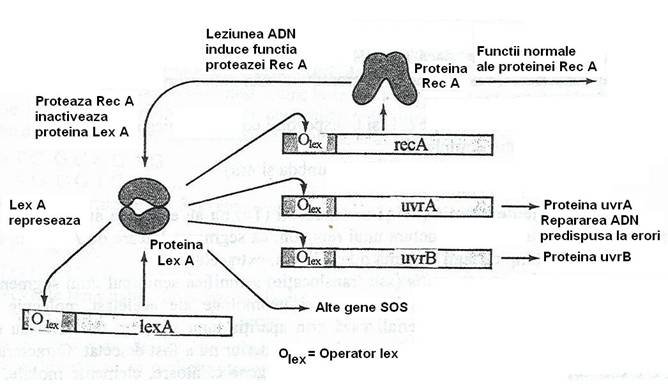
Fig. 124. Mecanismul raspunsului "SOS". Leziunea moleculei de ADN determina conversia proteinei Rec A intr-o proteaza care cliveaza proteina Lex A. Proteina Lex A, in conditii normale represeaza activitatile genei rec A si ale genelor de reparare uvr A si uvr B. Dupa inactivarea proteinei Lex A, aceste gene se activeaza (dupa Brock, 1988).
In absenta proteinei Rec A, proteina Lex A nu mai este clivata si continuarea sintezei sale reface rezerva celulara. Proteina represor Lex A se leaga de regiunea operator a genelor sistemului "SOS" si blocheaza transcrierea lor, inclusiv a genei lex A. Celula revine astfel, la starea normala, neindusa.
Repararea ruperilor dublu catenare. Ruperile dublei catene sunt produse de radiatia ionizanta, de agenti oxidanti, de erorile de replicare si de anumite produse metabolice ale celulei. Daca aceste leziuni nu sunt reparate, consecinta este ruperea cromosomului in fragmente mai mici. Sunt 2 mecanisme de reparare a ruperilor dublei catene:
reunirea si legarea neomologa a capetelor ADN, cu pierderea uneia sau mai multor nucleotide la situsul de legare. Mecanismul este comun in celulele mamiferelor;
legarea omologa, activa in celulele diploide: mecanismele de recombinare transfera secventa de nucleotide din dubla catena intacta, la situsul rupturii dublei catene.
Semnificatia biologica a sistemelor de reparare. Existenta mai multor mcanisme de raspuns la leziunile moleculei de ADN este consecinta actiunii agentilor fizici si chimici, care pot afecta structura si functia ADN.
Numarul leziunilor inactivatoare a unui genom, cu radiatii UV sau x creste odata cu complexitatea genomului.
Procesele reparatorii sunt avantajoase intr-un mediu in care actioneaza factori care induc mutatii cu o frecventa mare, pentru mentinerea stabilitatii genetice a organismului, dar pe de alta parte nu permit aparitia si exprimarea unor mutatii, esentiale pentru evolutia sistemelor vii.
Rata evolutiei unei specii ar putea fi cuplata cu capacitatea de discriminare a sistemelor de reparatie a ADN si cu eficienta actiunii lor.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3307
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved