| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Structura cromozomului la eucariote
Materialul genetic este reprezentat dintr-un genom nuclear si un genom extranuclear (reprezentat din genomul mitocondrial si, in cazul celulelor vegetale, si din genom cloroplastic).
Genomul nuclear cuprinde un set de molecule lineare de ADN, fiecare reprezentand un cromozom. Fara exceptie, toate eucariotele au cel putin 2 cromozomi care intotdeauna sunt lineari. Singura variatie care se inregistreaza este cea referitoare la numarul de cromozomi, care insa nu este corelata cu caracteristicile biologice sau cu pozitia evolutiva a organismului respectiv. Astfel, la Saccharomyces cerevisiae (drojdia de bere - un eucariot unicelular, inferior) numarul haploid de cromozomi este 16, de 4 ori mai mare decat la musca (Musca domestica). De asemenea, numarul de cromozomi nu este corelat nici cu dimensiunea genomului; de exemplu, la salamandra genomul este de 30 ori mai mare decat la om, dar are jumitate din numarul haploid de cromozomi de la om.
Cromozomii eucarioti sunt separati de citoplasma prin membrana nucleara si, ca urmare, transcrierea si traducerea sunt separate in timp si spatiu una de cealalta; in contrast, la procariote, cele doua procese se de sfasoara aproximativ simultan.
Nucleul este format din 4 componente: nucleolema, carioplasma, cromonemata (singular, cromonema) si nucleol. Cromonemata este o substanta cromatica despiralizata in interfaza si condensata in mitoza. Din aceasta substanta se diferentiaza cromozomii.
Nucleolul este o substructura nucleara cu rol in biogeneza ribozomilor. In aceasta zona a nucleului are loc sinteza de ARN ribozomal prin transcrierea genelor corespunzatoare.
La eucariote ADN este complexat cu o clasa de proteine specializate, cu caracter bazic, denumite histone. Acest complexpoarta numele de cromatina (denumirea are la baza tinctorialitatea fata decoloranti bazici).

Heterocromatina se clasifica in:
heterocromatina constitutiva - este caracteristica zonelor centromerice si telomerice
heterocromatina facultativa - este caracteristica unuia din cromozomii X la femelele de mamifer si constituie cromatina sexuala; apare datorita necesitatii compensatiei de doza a genelor esentiale prezente pe cromozomul X
heterocromatina de citodiferentiere - apare prin inactivarea selectiva a unor gene in timpul proceselor de citodiferentiere
Eucromatina este caracteristica genelor active transcriptional, in timp ce heterocromatina are rol preponderent reglator si structural.
1 Clase de secvente ADN la eucariote
secvente unice, o copie per genom haploid, corespunzatoare genelor codificatoare pentru lanturi polipeptidice, exceptie facand genele ce codifica histone, care se gasesc in catevasute de copii per genom haploid (este necesara o sinteza rapida intr-un timp scurt corespunzazoare perioadei S de replicare a ADN).
secvente moderat repetitive: sunt intr-un numar de 102 - 103 copii per genom haploid si codifica pentru ARN ribozomal, ARN de transfer si pentru histone.
secvente inalt repetitive: ajung pana la 106 copii per genom haploid
In general, eucariotele contin mult mai mult ADN decat procariotele. O celula de om contine de peste 1000 de ori mai mult ADN decat o celula de Escherichia coli. Astfel, ADN-ul dintr-o celula de om, aflat in stare decondensata atinge o lungime de 4 cm si nu poate incapea intr-un nucleu (de diametru 5 m) decat in forma condensata.
Forma cromozomilor dintr-o celula eucariota se modifica in timpul ciclului celular.
2 Proteinele cromozomale
Principalele proteine ce complexeaza ADN-ul au greutate moleculara mica, o cantitate mare de aminoacizi cu caracter bazic si se numesc histone. Au fost descrise 5 clase de histone: H1 H2A, H2B, H3 si H4.
Histona H1 este alcatuita din 3 domenii structurale distincte:
un capat amino-terminal cu 39 aminoacizi bazici
o regiune globulara centrala cu un diametru de 2,8 nm, cu aminoacizi cu caracter acid; interactioneaza cu alte proteinexi-terminal ce contine 40% lizina (cu puternic caracter bazic) si are afinitate de interactiune cu molecula de ADN.
Histonele H2A si H2B au cantitate mai redusa de lizina. H2A are leucina, iar H2B are serina si prolina. Au doar 2 domenii, dintre care unul interactioneaza cu ADN.
Histonele H3 si H4 sunt bogate in arginina si, in mod similar cu H2A si H2B, au tot doar 2 domenii functionale.
In general, histonele prezinta o structura inalt conservata in lumea vie. Cea mai heterogena este histona H1, acest parametru scazand de la H1 catre H4.
Genele pentru histone sunt organizate in unitati repetitive dispuse in tandem. De exemplu, la D.melanogaster unitatea repetitiva este formata din genele pentru:
H1 - H3 - H4 - H2A - H2B
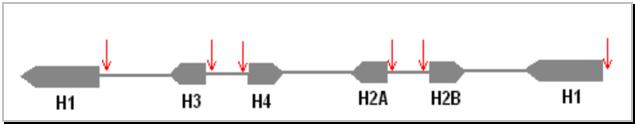
Figura 3.5 Organizarea genelor pentru histone la D.melanogaster.
![]() regiuni
reglatoare ale genelor (situsuri hipersensibile la nucleaza)
regiuni
reglatoare ale genelor (situsuri hipersensibile la nucleaza)
3 Nivelele de organizare ale cromatinei
Elementul structural de baza al cromatinei este nucleozomul care este format dintr-un octamer histonic central si o histona de legatura (linker). Nucleozomul are 145 perechi de baze corespunzatoare la aproximativ 2 ture de spira suprarasucite negativ in jurul miezului histonic. Miezul histonic prezinta domeniul hidrofob carboxi-terminal de interactiune cu celelalte proteine spre partea centrala, iar domeniul amino-terminal se gaseste la suprafata de interactiune cu ADN.
Intre 2 nucleozomi adiacenti se interpune ADN linker de 60 perechi de baze si este complexat cu histona H1.
Cromatina extinsa apare ca un sirag de margele cu un diametru de 11 nm. Prin intermediul histonei H1 implicata in superspiralizare interactioneaza 6 - 10 nucleozomi pentru a forma solenoidul de 30 nm. Acesta este unitate de baza a condensarii materialului genetic in interfaza.
Forma superioara de
condensare este reoprezentata de cromozomii metafazici, perioada in care
materialul genetic atinge un nivel maxim de condensare de 1400 nm.
Procesul de condensare a cromatinei este realizat prin atasarea ei la o clasa
de proteine denumite "proteine scaffold" (proteine de esafodaj). Atasarea
se realizeara cu formarea unor bucle (domenii) care au fiecare 100 kpb,
comparabile cu cele de la cromozomul bacterian.
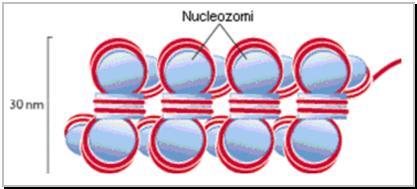
Figura 3.6 Reprezentarea schematizata a structurii nucleosomilor (dupa Strachan, 1999).
In organizarea materialului genetic nuclear la organisme eucariote intervin si proteine non-histonice. Acestea se subimpart in mai multe clase:
Ø enzime ale metabolismului ADN: ADN polimeraze, ADN ligaze, ADN topoizomeraze, terminal-ADN-nucleotidil-transferaze
Ø enzime ale metabolismului ARN: ARN polimeraze, poliA polimeraze, enzime de splicing, RNaze
Ø enzime ce modifica histonele: protein-kinaze, metilaze, acetilaze, esteraze, proteaze
Ø proteine structurale: proteinele fusului de diviziune, proteinele matricei nucleare, proteinele "scaffold", proteinele particulelor RNP ("RiboNucleoProtein")
Ø proteinele HMG (High Mobility Group) ce intervin in modificarea structurii cromatinei in vederea functionarii genelor in replicare si transcriere:
o HMG 1 si HMG 2 - proteine omoloage cu g.m. 29 kd;
o HMG 14 si HMG 17 - proteine omoloage cu g.m. 10-12 kd
Proteinele HMG inlocuiesc histona H1 determinand, alaturi de topoizomeraze, relaxarea helixului ADN in initierea transcrierii genelor.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4887
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved