| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Transformarea genetica
Transformarea genetica este o modalitate de transfer a informatiei genetice de la bacterie donoare la una receptoare, prin intermediul unui fragment de ADN (o fractiune a ADN total al celulei donoare), eliberat prin mecanisme fizice (plasmoliza), prin extractie chimica sau pe cale enzimatica.
Procesul transformarii genetice s-a descris la Streptococcus pneumoniae, Haemophilus sp., Bacillus subtilis, Neisseria sp., Rhizobium sp., Salmonella sp., Yersinia sp., Azotobacter sp. etc. si s-a demonstrat ca are loc atat intraspecific cat si interspecfic si chiar intre genuri diferite de bacterii.
Transformarea* propriu-zisa este un eveniment rar si este rezultatul integrarii fragmentului de ADN exogen, in genomul celulei receptoare prin procesul recombinarii si a exprimarii fenotipice a noilor determinanti genetici pe care ii contine.
*Fenomenul transformarii genetice a fost descoperit de Griffith (l928). El studia variabilitatea caracterelor de cultura la pneumococ (Str. pneumoniae), o bacterie care se prezinta sub forma unui diplococ, capsulat sau necapsulat. Pe baza compozitiei chimice si a specificitatii serologice a polizaharidelor capsulare, exista circa 80 de tipuri distincte. De exemplu polizaharidele capsulare de tip II sunt polimeri de acid galacturonic, galactoza, N-acetil-glucozamina si fucoza, iar cele de tip III sunt polimeri de glucoza si acid glucuronic. Sinteza acestor polizaharide este codificata de gene diferite. Celulele capsulate de pneumococ sunt virulente, iar cele necapsulate sunt nevirulente (dupa inocularea intr-un organism sensibil nu produc simptome de imbolnavire). Celulele capsulate cresc pe suprafata unui mediu solidificat, sub forma unor colonii netede (S, Smooth), iar cele necapsulate, produc colonii rugoase (R, Rough). Prin cultivarea tulpinilor capsulate si virulente (de tip S) in prezenta serului imun specific, se obtin variante necapsulare, avirulente, care cresc sub forma coloniilor R. Mutantele S - R apar cu o frecventa mult superioara mutantelor R - S. Modificarea tipului de colonie se face totdeauna in cadrul aceluiasi tip biochimic de polizaharid, fara modificarea tipului serologic.
Pentru a-si verifica ipoteza, Griffith a inoculat un numar mic de celule vii dintr-o tulpina R a tipului serologic II, avirulente, in amestec cu o cantitate mare de celule ale tulpinii S de tip serologic III, omorate prin caldura. Inoculate separat, nici una din cele doua tulpini bacteriene nu produce moartea animalelor. Amestecul celular produce infectia mortala la soarece, iar din sange se izoleaza pneumococi vii de tip III. Aparent, aceasta experienta a confirmat ipoteza de lucru a autorului: aparitia pneumococilor capsulati si virulenti ar fi rezultatul transferului polizaharidului de pe suprafata celulelor capsulate omorate, pe suprafata celulelor vii ale tulpinii R.
Experientele lui Griffith au fost reluate de Avery si colab. (l944). Ei au demonstrat ca la baza procesului de transformare genetica, sta transferul unor determinanti genetici, prin intermediul unui fragment de ADN, care confera celulei receptoare, capacitatea de a codfica polizaharidele capsulare. Sinteza materialului capsular determina aparitia corelativa a celorlalte particularitati: tipul coloniei (S sau R), virulenta. In acelasi mod pot fi transferati si alti determinanti genetici: cei care determina rezistenta la un antibiotic, sau care codifica sinteza unei enzime cu rol in metabolismul celular.
In experientele lui Griffith, deoarece mutanta R II de pneumococ, trece intr-o mutanta S III este evident ca un fragment de ADN al mutantei S III a fost transferat si s-a exprimat in celulele mutantei R II, permitand sinteza materialului capsular de tip serologic III si chiar activarea sintezei materialului de tip II.
Cercetarile diferitelor variante serologice patogene de pneumococ au evidentiat ca virulenta este o expresie cantitativa a gradului de patogenitate. In timp ce patogenitatea este o caracteristica de specie, gradul de virulenta a bacteriilor patogene, poate sa varieze de la avirulent pana la hipervirulent, trecand prin trepte intermediare.
Procesul de transformare genetica a fost studiat pentru reprezentantii mai multor genuri. Pentru ca transformarea sa aiba loc, sunt necesare cateva conditii:
ADN transformant trebuie sa aiba gr. mol. intre l-20 kDa;
fragmentul de ADN trebuie sa fie dublu catenar. ADN monocatenar nu este incorporat in celula sau incorporarea este ineficienta;
numarul celulelor transformate creste odata cu concentratia ADN in mediu, pana la nivelul de saturare (l-20 mg ADN/ml cultura bacteriana competenta, ceea ce corespunde la l00-200 fragmente de molecule de ADN/celula);
sa existe un grad de omologie genetica intre ADN transformant si o anumita secventa a cromosomului celulei receptoare. Astfel se explica frecventa superioara a recombinarilor genetice pentru transformarile intraspecifice si scaderea frecventei lor, pe masura ce se accentueaza diferentele de structura genetica;
acceptarea ADN si transformarea sunt conditionate de o anumita stare fiziologica a celulei receptoare, denumita "stare de competenta", cu o durata variabila.
Produsele genice necesare transformarii sunt clasificate in doua grupe;
a) proteinele genelor timpurii, cu rolul de a determina starea de competenta;
b) proteinele tarzii necesare pentru legarea, prelucrarea ADN si asamblarea mecanismelor de translocatie a ADN.
Competenta defineste o stare particulara de permeabilitate crescuta a structurilor suprafetei celulare, care permite legarea si inglobarea unui fragment de ADN exogen, intr-o forma rezistenta la actiunea DN-azei.
Starea de competenta limiteaza foarte mult frecventa procesului de transformare, pentru ca, desi ADN este adeseori disponibil (din celulele lizate intr-o cultura), transformarea este conditionata de starea fiziologica a celulelor acceptoare.
Starea de competenta poate fi naturala (fiziologica) sau artificiala (indusa). Competenta naturala, la unele bacterii, este permanenta, iar la altele este temporara.
La bacteriile Gram pozitive (B. subtilis, Str. pneumoniae), competenta naturala este temporara si este indusa de sinteza unor factori difuzibili, denumiti factori de competenta, a caror concentratie este proportionala cu densitatea celulara(''quorum sensing''* ).
*Quorum sensing este definita ca fenomenul de reglare a expresiei genice dependent de densitatea celulara. Bacteriile isi coordoneaza reglarea expresiei genice, producand molecule-semnal difuzibile: aminoacizi, AMPc, oligopeptide, dipeptide ciclice, derivati ai acizilor grasi etc. Acestea se acumuleaza extracelular si interactioneaza specific cu o molecula receptoare, pentru a induce modificari fiziologice, nelegate de metabolismul propriu. Moleculele semnal induc un raspuns dupa ce au atins o concentratie limita.
Competenta este o stare fiziologica temporara si reversibila, se instaleaza la sfarsitul fazei de crestere exponentiala si este asociata cu scaderea ratei sintezei ADN si cu sinteza unor proteine specifice localizate in membrana citoplasmatica, care determina o stare fiziologica particulara a structurilor suprafetei celulare.
In faza de competenta, numai l0% din celulele unei populatii devin competente pentru transformare, probabil ca o consecinta a unei reglari individuale la nivel celular.
Alteori, starea de competenta depinde de stari particulare ale invelisului celular, consecinta a sintezei unor proteine de membrana cu rol de receptori pentru ADN, si a unor autolizine. S-a descris la Str. pneumoniae, B. subtilis, H. influenzae, A. vinelandii, Neisseria sp., Pseudomonas sp., Rhizobium sp., Xanthomonas sp.
Bacteriile Gram negative nu sintetizeaza factori de competenta, starea fiind reglata prin mecanisme metabolice interne si este indusa in conditii de deficit nutritional, asociate cu incetinirea sau limitarea cresterii. Toate celulele unei culturi devin competente si starea este mentinuta in medii care limiteaza cresterea.
La Neisseria sp. si la alti gonococi, starea de competenta este constitutiva si permanenta, nefiind dependenta de stadiul culturii si nici de sinteza factorilor de competenta.
Competenta artificiala este indusa de :
concentratiile relativ mari de CaCl2 sau cloruri de Mg, Ba, Rb, Sr si amestecurile lor;
tratamentul celulelor cu agenti chelatori (EDTA) ;
tratamentul cu enzime (muramidaze sau peptidaze), ce duc la formarea sferoplastilor sau protoplastilor ;
fuziunea protoplastilor cu lisosomi purtatori de ADN;
expunerea celulelor la socuri electrice (electroporare);
bombardarea celuleor cu particule mici ce transporta ADN in citoplasma (transformarea biolostica).
Celulele de B. subtilis devin competente in faza stationara, in prezenta ionilor de Mg2+ si Ca2+. De obicei, se folosesc combinatii ionice de CaCl2 (30 mM) si MgCl2 (26 mM).
O metoda eficienta de inducere a starii de competenta, este electroporarea, ce consta in permeabilizarea invelisurilor celulare si usurarea inglobarii ADN exogen, sub actiunea stimulilor electrici.
Mecanismul molecular al transformarii genetice
Mecanismul molecular al transformarii genetice a fost descris la B. subtilis, Streptococcus sp., Haemophilus sp. Transformarea genetica este produsa numai de moleculele de ADN dublu catenare, lineare, in timp ce ADN monocatenar, ARN, hibrizii ADN-ARN sau ADN circular inchis, nu sunt legate pe suprafata celulelor competente. Procesul transformarii poate fi impartit in trei etape:
legarea moleculei de ADN pe suprafata celulei si fragmentarea ei;
inglobarea (transportul ADN prin invelisurile celulei);
stabilizarea ADN transformant, fie ca replicon autonom, fie prin recombinare cu un replicon rezident.
Mecanismul transformarii este asemanator la bacteriile Gram pozitive si la cele Gram negative, dar exista diferente ale etapelor de legare si inglobare, datorate diferentelor structurale majore ale peretelui celular.
Intr-o prima etapa, fragmentul de ADN exogen este adsorbit reversibil, consecinta a unei interactiuni laxe. Peretele celulelor Gram pozitive are o sarcina neta pozitiva, iar ADN are o sarcina electrica negativa. Din aceasta cauza, adsorbtia ADN este rapida. Faza adsorbtiei reversibile dureaza 4-5 secunde. Molecula de ADN poate fi indepartata prin spalare rapida.
Faza de legare "stransa" sau de adsorbtie ireversibila, corespunde ancorarii ferme a moleculei de ADN, la cateva situsuri din cele circa 50 ale celulei receptoare.
La B. subtilis, in legarea ADN exogen este implicata o proteina complexa de 75 kD, asociata membranei, sintetizata sub actiunea factorului de competenta. Proteina de legare este formata din doua polipeptide echivalente ca marime, una dintre ele avand activitate endonucleazica. Deoarece are rol decisiv in transportul ADN exogen in celula a fost denumita translocaza. Ambele faze ale legarii sunt nespecifice, deoarece celulele de B. subtilis pot sa lege fragmente de ADN de provenienta straina (de exemplu, ADN de mamifer).
Transportul ADN prin invelisul celular (inglobarea) este putin inteles. Patrunderea ADN exogen in celula receptoare se face sub forma monocatenara. Studiile experimentale cu molecule marcate radioactiv au evidentiat o injumatatire a radioactivitatii in interiorul celulei, ceea ce sugereaza ca transportul ADN se face sub forma monocatenara. {n timpul transportului, catena complementara este degradata complet. Inglobarea implica incizia unei catene a ADN exogen, la sau langa situsurile de legare la membrana, in prezenta ionilor bivalenti (Mg2+, Ca2+) si este catalizata de nucleaza din complexul de 75 kD. Urmeaza incizii succesive monocatenare, astfel incat una dintre catenele de ADN exogen este degradata complet.
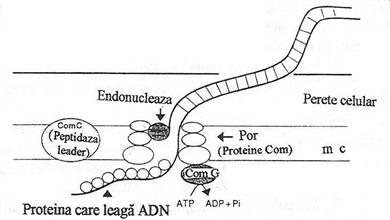
Fig. 129. Modelul molecular al transformarii genice la bacteriile Gram pozitive. Canalul transmembranar este probabil format de proteinele de competenta. In timpul trecerii prin membrana citoplasmatica, o catena a ADN dublu catenar exogen este degradata de o endonucleaza asociata membranei (dupa Dreiseikelmann, 1994).
Inglobarea ADN monocatenar in celula receptoare pare a fi rezultatul actiunii endonucleazei. Transportul se face, probabil, printr-un canal membranar, format prin modificarea conformatiei spatiale a nucleazei. La B. subtilis, inglobarea ADN necesita forta proton-motrice si nu hidroliza ATP.
ADN monocatenar este protejat de actiunea nucleazelor in timpul transportului, prin formarea unui complex cu o proteina specifica de legare, sintetizata in intervalul de competenta.
La bacteriile Gram negative (Haemophilus sp., N. gonorrhoeae), ADN exogen este legat si transportat in transformasomi (vezicule membranare de transport) cu diametrul de 20 nm, alcatuiti in special, din componentele membranei externe. In transformasom, ADN este protejat de actiunea DN-azei exogene si de endonucleazele celulare de restrictie.
La iesirea din vezicula de transport, o catena a ADN exogen este degradata complet.
ADN inglobat de celulele competente, poate fi izolat sub forma fragmentelor monocatenare de circa l0 kb.
Moleculele suprahelicale si chiar unele relaxate nu sunt transportate in citoplasma. Structura spiralizata a plasmidelor sau absenta capetelor libere impiedica translocatia ADN din transformasom in citoplasma. Astfel se explica efocienta scazuta a transformarii, cu ADN plasmidial.
Stabilizarea ADN monocatenar transformant, este consecinta unui proces de integrare prin recombinare. ADN monocatenar este protejat de actiunea nucleazelor, de o proteina specifica sintetizata in faza de competenta, proteina SSB (Single Strand Binding).
Integrarea ADN monocatenar exogen in cromosomul celulei receptoare, este catalizata de enzime si este dependenta de proteina Rec A* (produsul genei rec A), printr-un proces de recombinare. Rezultatul final al recombinarii, este o molecula hibrida (heteroduplex). Fragmentul de ADN monocatenar exogen se aliniaza pe baza complementaritatii bazelor, cu o regiune omologa a cromosomului. Se formeaza un complex ADN exogen-ADN receptor. Recombinarea propriu-zisa are loc prin deplasarea fizica si inlocuirea uneia dintre catenele ADN receptor. Deplasarea catenei este insotita concomitent, de degradarea ei si a altor molecule de ADN exogen transformant. Daca ADN exogen nu are omologie cu ADN cromosomal, ADN exogen este degradat complet.
Activitatea enzimatica unica a proteinei Rec A consta in imperecherea moleculelor omologe de ADN, atat monocatenare cat si dublu catenare si favorizarea schimbului reciproc al catenelor de ADN intre moleculele omologe. Schimbul genetic implica, suplimentar, activitatea altor enzime: ADN-helicaze, DN-aze, ATP-aze, topoizomeraze, ADN-polimeraza si ADN-ligaza.
Proteina Rec A este o ATP-aza, dependenta de ADN. In vitro, proteina Rec A favorizeaza formarea perechilor omologe intre moleculele de ADN adecvate. Daca nu sunt constrangeri topologice care sa impiedice formarea heteroduplexului, va rezulta o legare plectonemica a moleculelor (catenele sunt legate intr-un ADN dublu catenar). Daca sunt constrangeri topologice (cele 2 catene pot fi circulare) se vor forma perechi paranemice (se formeaza perechi de baze, dar catenele nu sunt topologic imperecheate).
Pentru toate reactiile omologe de imperechere, catalizate de Rec A, este necesar ADN monocatenar. Pentru complexele paranemice, ADN mc este esential. Necesitatea ADN mc in procesele de imperechere dependente de proteina Rec A, explica necesitatea actiunii helicazelor si nucleazelor in recombinarea genetica. Helicazele produc ADN mc prin despiralizarea ADN dc. Nucleazele produc ADN mc prin degradarea unei catene a ADN dc.
Reactia de schimb intre ADN cromosomal si ADN exogen este conditionata de proteina Rec A. Actiunea proteinei Rec A consta in alinierea celor doua molecule de ADN, pe baza complementaritatii nucleotidelor, intr-o reactie dependenta de ATP. Alinierea este urmata imediat de formarea "buclei D", in care ADN monocatenar invadeaza ADN dublu catenar rezident. Recombinarea parcurge urmatoarele trei etape:
l. Polimerizarea proteinei Rec A pe molecula de ADN monocatenar. In vitro, proteina Rec A si ADN monocatenar formeaza un filament nucleoproteic evidentiat la microscopul electronic.
2. Sinapsa. In prezenta ATP, nucleoproteina formeaza un complex cu ADN cromosomal. Interactiunea initiala nu se produce intre secventele de bazele omologe. Cele doua molecule de ADN se deplaseaza una fata de alta, astfel incat secventele omologe vin in contact direct si se aliniaza pe baza omologiei nucleotidelor.
3. Schimbul postsinaptic. Catenele omologe, aliniate dar neimperecheate, se leaga instabil. Proteina Rec A deplaseaza o catena a moleculei dublu catenare si catalizeaza integrarea catenei exogene, cu consum de energie, furnizata prin hidroliza ATP. Proteina Rec A are actiune de helicaza si produce despiralizarea ADN dublu catenar. Integrarea ADN este polarizata in directia 5' ---- 3'.
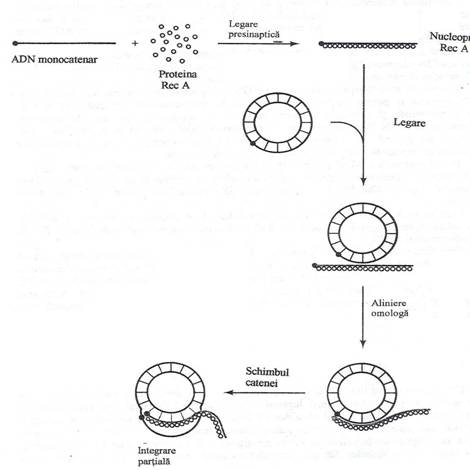
Fig. 130. Etapele procesului de recombinare a secventelor omologe, dependenta de proteina Rec A.
La Str. pneumoniae, rolul proteinei Rec A pare a fi indeplinit de SSB. Catena inlocuita este sectionata. Nucleazele sectioneaza capetele libere ale ADN exogen si endogen, iar ligaza repara inciziile. Se formeaza un heteroduplex, in care unele baze sunt imperecheate gresit. Rezultatul recombinarii, adica mentinerea sau eliminarea ADN exogen in genomul celulelor progene, depinde de repararea imperecherilor gresite.
Unii markeri transforma celulele cu o eficienta foarte inalta. Imperecherile de baze gresite ori sunt corectate eficient si aduse la nivelul secventei de baze a ADN exogen, ori corectarea este nesemnificativa. Pentru markerii care au eficienta transformanta scazuta, mecanismele de reparare a bazelor imperecheate gresit sunt foarte active. Sunt indepartate bazele ADN exogen si celula pastreaza fenotipul conferit de ADN propriu. Dovada pentru validitatea acestei concluzii a fost adusa de izolarea mutantelor defective pentru repararea bazelor imperecheate gresit. La aceste mutante, toti markerii de transformare au eficienta foarte inalta.
Fenomenul recombinarii este rar, avand loc in circa l% din totalul evenimentelor de inglobare a ADN exogen. De cele mai multe ori, ADN exogen nu se poate recombina datorita fenomenului de restrictie, un adevarat mecanism de imunitate moleculara, prin care ADN strain este recunoscut si eliminat.
Exista doua tipuri de ADN, care dupa inglobarea in celula competenta, nu necesita integrarea: ADN plasmidial si fagic. Neavand omologie a bazelor fata de ADN cromosomal, nu se integreaza, ci raman ca repliconi in stare fizic autonoma.
Studiile de transformare s-au facut cu ADN cromosomal, pentru markerii de prototrofie si cu ADN plasmidial pentru markerii de rezistenta la antibiotice. Se inregistreaza aparitia coloniilor prototrofe si respectiv a celor rezistente la antibiotice, pe mediile adecvate. Transformarea cu ADN fagic se inregistreaza sub forma plajelor de liza pe o panza de celule sensibile si nu depinde de prezenta secventelor de baze omologe. Este o transformare independenta de capacitatea celulei de a face recombinare omologa.
Transformarea cu ADN plasmidial sau fagic are loc cu o frecventa foarte mica si necesita cooperarea unui mare numar de copii moleculare, echivalent la circa l0 000 echivalenti ai genomului fagic si l000-l0 000 molecule de ADN plasmidial, pentru un eveniment de transformare.
Dimensiunea medie a ADN integrat in celulele de B. subtilis s-a estimat la 8-l0 kb. Fiecare celula poate integra l0 molecule de 8-l0 kb fiecare, ceea ce coresunde la circa l00 de gene de dimensiuni medii.
Transformarea genetica, descoperita in laborator, este un fenomen care se desfasoara cu mare probabilitate si in conditii naturale, in mediile in care se realizeaza populatii bacteriene foarte dense si unde au loc procese de liza celulara, prin care se elibereaza fragmente de ADN: liza fiziologica, liza fagica sau sub actiunea substantelor antibiotice. Fragmentele de ADN sunt preluate de alte celule prin mecanismul transformarii, asigurand o variabilitate genetica cu o frecventa semnificativa.
Functiile inglobarii ADN :
variatia proprietatilor antigenice (de exemplu, ale pilinei) este o strategie pentru invazia bacteriilor patogene;
protectia celulelor fata de fagi;
inglobarea ADN ca o cale de dobandire a nutrientilor. Produsele rezultate din degradarea ADN heterolog raman disponibile metabolismului celular si sunt folosite ca precursori ai sintezei ADN sau ca sursa de C, N si P;
repararea ADN: ADN exogen homolog ar putea fi folosit pentru repararea prin recombinare, a leziunilor ADN cromosomal al receptorului, induse de stresul genotoxic. Ipoteza presupune implicit, ca celulele competente supravietuiesc mai bine decat cele necompetente.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2215
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved