| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
DOCUMENTE SIMILARE |
|||||||||||
|
|||||||||||
Cai de semnalizare induse de concentratia crescuta de glucoza asociata diabetului in celulele peretelui vascular
Desi, in ultimii ani, au fost efectuate multe studii care au aratat ca nivelele crescute de glucoza pot induce disfunctia diferitelor cascade de semnalizare, mecanismele prin care hiperglicemia determina aparitia bolilor cardiovasculare nu sunt pe deplin elucidate. Dat fiind ca exista numeroase studii care au demonstrat ca stresul oxidativ, mediat in principal de radicalii liberi indusi de hiperglicemie, contribuie la dezvoltarea si progresia diabetului si complicatiilor acestuia, este normal sa ne gandim ca mediatorii activarii cailor de semnalizare in diabet sunt in principal speciile reactive de oxigen (ROS). In timp ce ROS generate in conditii fiziologice fuctioneaza ca molecule de semnalizare in mecanismele de aparare ca fagocitoza, functia neutrofilelor si vasorelaxare indusa de stresul produs de fluxul de sange, generarea excesiva a stresului oxidativ are consecinte patologice, afectand proteinele, lipidele si ADN.
Ideea ca ROS ar pute functiona ca mesageri secundari de semnalizare a aparut prima data in anii 1970s cand s-a demonstrat ca H2O2 exogen mimeaza actiunea insulinei si ca insulina si factorii de crestere stimuleaza productia celulara de H2O2 [Mukherjee et al., 1978]. Ulterior, au aparut dovezi care arata ca ROS endogene participa la cascadele de semnalizare in multe tipuri celulare [Forman et al., 2002; Forman & Torres, 2002]. Astfel, diferiti antioxidanti si inhibitori ai sistemelor generatoare de ROS inhiba caile de semnalizare mediate de diversi agonisti, printre care glucoza crescuta. Tintele majore ale ROS includ factori de transcriptie, protein tirozin fosfataze, protein tirozin kinaze (PTK), protein kinaze activate de mitogen (MAPK), canale ionice, fosfolipide (figura 13) [Droge, 2001]. Toate aceste tinte sunt de asemenea reglate de concentratiile crescute de glucoza, prin intermediul ROS.
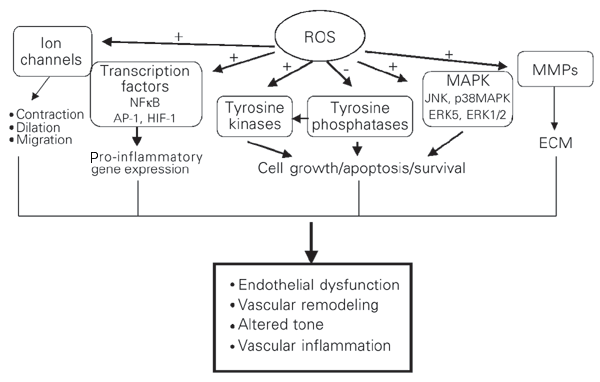
Figura 13. Cai de semnalizare redox-dependente. Speciile reactive de oxigen modifica activitatea tirozin kinazelor ca: Src, Ras, JAK2, Pyk2, PI3K, si EGFR, precum si a MAPK, in particular p38MAPK, JNK si ERK5. ROS pot inhiba activitatea protein-tirozin fosfatazelor, contribuind in plus la activarea protein tirozin kinazelor. De asemenea, ROS influenteaza expresia genica si proteica prin activarea factorilor de transcriptie NFkB, activator protein-1 (AP-1) si factorul ce induce hipoxia (HIF-1). ROS stimuleaza canale ionice de Ca2+ si K+ ducand la modificari ale concentratiei cationilor. Activarea acestor cai sensibile-redox de catre glucoza crescuta duce la diferite raspunsuri celulare care contribuie la aparitia bolilor cardiovasculare asociate diabetului. (-, efect inhibitor; +, efect stimulator; ECM, matrice extracelulara; MMPs, metaloproteaze matriceale; dupa Touyz et al., 2002)
IV.1. Factori de transcriptie
Factorii de transcriptie au fost primele proteine semnalizatoare identificate ca sensibile-redox. Factorul de transcriptie kB (NF-kB), care este activat de glucoza crescuta in celulele vasculare, este prototipul factorilor de transcriptie sensibili-redox. Desi diabetul si hiperglicemia induc multiple modificari in metabolism si semnalizare, studiile existente sugereaza ca activarea factorului de transcriptie NF-kB poate fi o trasatura semnificativa a complicatiilor diabetice. Recent, NF-kB activat a fost asociat cu placile aterosclerotice la pacientii cu diabet de tip2 [Cipollone et al., 2003]. Diverse tipuri celulare ca celule musculare netede [Yerneni et al., 1999], celule endoteliale [Chen et al., 2003; Li et al., 2003; Golovchenko et al., 2000], pericite [Romeo et al., 2002], celule mesangiale [Ha et al., 2002] prezinta activitate crescuta a factorului de transcriptie NF-kB cand sunt activate de concentratii de glucoza crescuta. ROS influenteaza acivitatea NF-kB prin modificarea oxidativa a reziduurilor cisteinice, prin degradarea IkB si prin declansarea cascadelor de semnalizare a caror tinta este NF-kB [Droge, 2001]. NF-kB regleaza transcriptia multor gene implicate in inflamatie si crestere, incluzand interleukine, molecule de adeziune si proto-oncogene [Thannickal & Fanburg, 2001]. Alti factori de transcriptie activati de ROS sunt AP-1 si HIF-1. AP-1 este un factor de transcriptie complex format prin homo sau heterodimerizarea membrilor familiilor c-Jun si c-Fos si influenteaza diferentierea si cresterea celulelor vasculare. ROS regleaza activitatea AP-1 prin intermediul a numeroase mecanisme si tinte, incluzand glutationarea-S a unei singur reziduu cisteinic conservat, reglarea redox reversibila prin tioredoxina si proteina nucleara Ref1 [Xanthoudakis S & Curran,1992] si prin reglarea cascadei c-Jun N-terminal kinazei (JNK). ROS produs de glucoza crescuta stimuleaza calea p38 MAP kinaza, care in plus, activeaza factorul de transcriptie AP-1. Activarea AP-1 duce la transcriptia genelor inflamatoare, avand ca efect cresterea productiei endoteliale de IL-8 (figura 14). Eliberarea chemokinei IL-8 de celulele endoteliale mediaza recrutarea si adeziunea monocitelor la endoteliu in inflamatie si complicatiile vasculare ale diabetului [Srinivasanet al., 2003].

Figura 14. Activarea p38MAPK de glucoza
IV. 2. Proteine tirozin fosfataze si proteine tirozin kinaze
In prezent, tintele moleculare directe cel mai bine stabilite ale ROS sunt protein tirozin fosfatazele (PTP). Fosforilarea protein tirozinelor este un mecanism major pentru modificarea post-translationala a proteinelor si joaca un rol critic in reglarea proliferarii, diferentierii, migrarii si transformarii celulare. Nivelul de fosforilare a tirozinelor in celula este controlat de balanta dintre PTK (protein tirozin kinaze) si PTP [Anderson et al., 2001]. Prin defosforilarea proteinelor substrat ale PTK, PTP contracareaza efectele activitatii PTK. Astfel, PTP pot fi considerate ca reglatori negativi ai procesului de semnalizrae initiat de PTK. Expunerea celulelor la doze scazute de oxidanti induce o crestere a fosforilarii tirozinice datorita inactivarii PTP.
Protein tirozin fosfataze. PTP sunt o familie diferita structural de enzime receptor si non-receptor care sunt reglatori critici pentru multe cai de semnalizare [Anderson et al., 2001]. Datorita structurii lor particulare, PTP sunt susceptibile la oxidare si la inactivarea de catre ROS. Toate enzimele PTP poseda un domeniu conservat de 230 aminoacizi care contin o cisteina reactiva (aceasta putand fi reglata redox ) care catalizeaza hidroliza reziduurilor fosfotirozinice ale proteinei prin formarea unui intermediar cisteinil fosfat [Denu & Tanner, 1998]. Aceasta cisteina formeaza tiol fosfatul, un intermediar in reactia de defosforilare a PTP. Oxidarea acestui reziduu cisteinic la acid sulfenic de catre H2O2 face ca PTP sa devina complet inactiv. Dat fiind ca oxidarea PTP este ireversibila, PTP exista in doua forme: o stare activa cu o cisteina redusa si o stare inactiva cu o cisteina oxidata. Activarea sau inactivarea PTP este reglata de semnale extracelulare ca hiperglicemie [Wu X et al., 2005] si EGF, iar H2O2 joaca rol major ca mesager secundar in acest proces [Denu & Tanner, 1998]. Transducerea semnalului insulinei in adipocite este insotit de de consumarea H2O2 care faciliteaza semnalizarea insulinei, prin inhibarea PTP dependente de tiol. PTP sunt reglatori negativi ai semnalizarii insulinei. Cum hiperglicemia este asociata cu cresterea ROS, s-a aratat ca nivelele crescute de glucoza pot amplifica fosforilarea proteinelor stimulate de insulina. Activitatea PTP endogena bazala si activitatea PTPB1, o PTP implicata in reglarea negativa a semnalizarii insulinei, sunt reduse in conditii de glucoza crescuta, iar reducerea lor ulterioara de catre semnalizarea prin insulina este mai mare in mediu cu glucoza crescuta, comparativ cu mediulcu concentratie scazuta de glucoza [Wu X et al., 2005]. De asemenea, s-a demonstrat ca inactivarea PTP 1B de catre EGF este dependenta de oxidarea reversibila a cisteinelor de catre H2O2. Studii recente sugereaza ca PTP 1B poate fi reglata mai eficient de O2- decat de H2O2 [Barrett et al., 1999]. Peroxinitritii inhiba rapid si ireversibil PTP sustinand rolul acestor ROS in daunele facute de stresul oxidativ. In afara de fosfatazele solubile, receptorii PTP (RPTP) pot fi de asemenea modulati de stresul oxidativ [Blanchetot et al., 2002].
Protein tirozin kinaze. Fosforilarea tirozinei este una din modificarile covalente cheie care are loc in organismele multicelulare, ca rezultat al comunicarii intercelulare in timpul embriogenezei si dezvoltarii tesuturilor adulte. Enzimele care faciliteaza aceste modificari sunt protein tirozin kinazele (PTK), care catalizeaza transferul unei grupari fosfat de la ATP la reziduurile tirozinice ale sbstraturilor proteinelor. Fosforilarea reziduurilor tirozinice moduleaza activitatea enzimatica si creaza situsuri de legare pentru recrutarea proteinelor semnalizatoare. In celula sunt prezente doua tipuri de PTK: PTK receptori (transmembranare) si PTK nonreceptori. Pentru ca PTK sunt componente critice ale cailor de semnalizare, activitatea lor catalitica este reglata strict. Semnalizarea necontrolata a tirozin kinazelor poate duce la raspunsuri inflamatoare si boli ca ateroscleroza, cancer si psoriazis.
Tirozin kinazele sunt de asemenea tinte ale speciilor reactive de oxigen [Droge, 2001; Thannickal & Fanburg, 2001; Lee et al., 1998]. In conditii patologice asociate cu stresul oxidativ, ROS pot activa direct receptorii suprafetei celulare, amplificand procesul de generare a O2-. Tirozin kinazele care nu sunt receptori ca Src, JAK2, STAT, p21Ras, Pyk2 si Akt, sunt stimulate ca raspuns la glucoza crescuta (prin intermediul ROS) si sunt implicate in remodelarea cardiovasculara si lezarea vasculara
MAP kinazele. MAP kinazele sunt o familie de serin/treonin kinaze care participa la caile clasice de transducere a semnalului, asociate cu diferentierea celulara, cresterea si moartea celulei [Torres & Forman, 2003]. Printre principalele MAP kinaze, ERK1/2, p38MAP kinaza si JNK sunt cele mai studiate si mai bine caracterizate. ERK1/2, fosforilata de MEK1/2 (MAP/ERK kinaza) este kinaza cheie in semnalizarea cresterii, in timp ce JNK si p38 MAP kinaza, fosforilate de MEK4/7 si respectiv MEK3/6, influenteaza supravietuirea celulara, apoptoza, diferentierea si inflamatia. ERK5, reglata de MEK5, este implicata in sinteza proteica, progresia ciclului celular si cresterea celulara. In celulele vasculare, toate MAPK sunt reglate, mai mult sau mai putin, de glucoza crescuta [Liu et al., 2000; Igarashi et al., 1999; Takaishi et al., 2003]. Activarea crescuta a MAP kinazelor a fost demonstrata in hipertensiune, ateroscleroza si diabet, si pare sa fie un mecanism major care contribuie la lezarea vasculara asociata cu aceste boli [Torres & Forman, 2003; Touyz et al., 2002]. MAP kinazele sunt reglate de cascade de fosforilare si sunt puternic activate de ROS. Mecanismele prin care ROS activeaza MAP kinazele nu sunt clare, insa, posibile tinte sunt MAP kinazele fosfataze (MKP). Similar cu PTP, MPK au o cisteina sensibila - redox, esentiala, conservata care confera activitate catalitica. Inhibarea MKP de catre ROS, prin modificari oxidative, duce la activarea MAP kinazelor [Thannickal & Fanburg, 2000]. De fapt, scaderea activitatii fosfatazei a fost legata de cresterea activitatii ERK1/2 vasculare in hipertensiune [Begum et al., 1998]. Alte procese prin care ROS influenteaza MAP kinazele pot fi declansate de activatori up-stream, ca tirozin kinaza Src, GTPaza Ras si protein kinaza C [Forman et al., 2002; Forman & Torres, 2002]. Kinaza p38 este un membru al familiei de serin/treonin protein kinaze activate de mitogen care este activata prin dubla fosforilare a Thr180 and Tyr182 de catre MAP kinaza kinaza. In celulele endoteliale calea p38 MAP kinaza este activata de stimuli ce induc stres, incluzand ROS, hiperglicemie si cytokine proinflamatoare [Surapisitchat et al., 2001]. Activarea p38 MAPK regleaza inducerea genelor inflamatoare si activarea altor cai de semnalizare inflamatoare, inclusiv NF-kB [Evans et al., 2002]. Datele existente pana in prezent demonstreaza ca fosforilarea si activarea caii p38 MAPK sunt implicate in reorganizarea actinei in citoscheletul celulelor endoteliale si inducerea expresiei moleculelor de adeziune, ducand la adeziunea si migrarea leucocitelor [Landry et al., 1995; Rousseau et al., 1997]. Astfel, activarea caii p38 in endoteliul vascular prin nivele crescute, cronice, de glucoza, poate contribui semnificativ la dezvoltarea complicatiilor vasculare in diabet. Inhibarea caii p38 MAPK, in celulele endoteliale, blocheaza producerea de interleukina-8 si adeziunea monocitelor la celulele endoteliale izolate de la soarecii diabetici. Aceste studii demonstreaza ca activarea cai p38MAP kinza in celulele endoteliale declanseaza evenimentele de semnalizare proinflamatoare care duc la recrutarea si adeziunea monocitelor.
Sisteme de transport al calciului. In plus la influentarea cailor de semnalizare asociate cu cresterea celulara si inflamatia, ROS moduleaza concentratia intracelulara de calciu ([Ca2+]i), determinant major al contractiei vasculare. Superoxidul si H2O2 induse de concentratia mare de glucoza cresc [Ca2+]i in celulele musculare netede si celulele endoteliale [Dragomir et al., 2004; Lounsbury et al., 2000]. Aceste efecte au fost atribuite mobilizarii calciului indusa de inozitolul trifosfat dependent redox, cresterii influxului de Ca2+ si scaderii activarii Ca2+-ATPazei [Gao & Lee, 2001]. Canalele de K+ din membrana plasmatica a CMN vasculare care controleaza o hiperpolarizare necesara relaxarii, sunt deschise printr-un mecanism asociat cu oxidarea tiolului de catre ROS [Droge, 2001]. Recent, studiile noastre au aratat ca [Ca2+]i in celulele endoteliale activate de glucoza crescuta este mai mare fata de [Ca2+]i din celulele control, nivelele de ROS fiind de doua ori mai mari in celulele activate de glucoza [Dragomir et al., 2004]. Aceste date sugereaza ca in plus la disfunctia vasodilatarii indusa de endoteliu (datorita anihilarii crescute a NO de catre O2-), modificarile Ca2+ induse de stresul oxidativ ar putea contribui la alterarea tonusului vascular.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1599
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved