| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Ribosomii sunt particule ribonucleoproteice, localizate in citoplasma, care la microscopul electronic au forma sferica, cu diametrul de 20 nm.
Pe baza constantei de sedimentare* (S) se disting urmatoarele categorii de ribosomi: a) ribosomi 80S, in citoplasma celulelor eucariote; b) ribosomi mitocondriali si cloroplastici, intre 55S la mamifere si 75S la plantele superioare; c) ribosomii archaea, de 70S, asemanatori din punct de vedere functional cu ribosomii 80S ai eucariotelor, datorita absentei sensibilitatii la streptomicina si cloramfenicol si prin sensibilitatea la toxina difterica; d) ribosomii eubacteriilor, de 70S.
Simbolul S semnifica viteza de sedimentare a unei particule sau a unei molecule prin tehnica ultracentrifugarii. Constanta de sedimentare (viteza cu care o molecula se deplaseaza intr-un camp gravitational) este dependenta de doua proprietati fizice ale moleculei:
gr. mol. (M): cu cat gr. mol. creste, cu atat creste si viteza de sedimentare;
aspectul moleculei (o molecula aerodinamica se deplaseaza mai repede decat una sferica). O molecula mai compacta intampina o forta de frecare mai mica si se deplaseaza mai repede.
Raportul dintre viteza de deplasare a moleculei si forta centrifuga aplicata se numeste coeficient de sedimentare(S).
S = Viteza/Forta centrifuga
Valoarea lui S pentru o molecula data este aceiasi in diferite solutii.
Pentru majoritatea macromoleculelor, valoarea lui S este cuprinsa intre 1 x l0-13 secunde si l00 x l0-l3 secunde. In onoarea lui Svedberg, cel care a inventat centrifuga, l0-13 secunde este echivalentul unui svedberg sau S.
Numarul ribosomilor in celula bacteriana este corelat cu activitatea ei fiziologica: este mic in celulele in repaus, dar creste foarte mult in celulele fiziologic active (in medie 20000 ribosomi/celula, cu variatii intre l5-l00 000).
Ribosomii sunt structuri dinamice, calitate ce se reflecta in capacitatea lor de a se disocia in doua subunitati, de 30 S si 50S si de a se reasocia. Disocierea si reasocierea sunt corelate cu variatia concentratiei ionilor de Mg2+: cresterea concentratiei ionilor favorizeaza asocierea, iar scaderea concentratiei lor produce disocierea. Circa l0% din numarul total de ribosomi sunt asamblati, liberi in citoplasma. Ei sintetizeaza proteinele structurale. Alti l0% se gasesc sub forma subunitatilor disociate, iar restul de 80% sunt polisomi.
Ribosomii au doua localizari: liberi in citoplasma sau atasati fetei interne a membranei citoplasmatice. La nivelul celor atasati se sintetizeaza proteinele de export. Studiul ribosomilor a beneficiat de tehnici din domeniul fizicii (metoda dispersiei neutronilor), care, in asociatie cu tehnicile de chimie au permis intelegerea structurii si functiei ribosomilor.
La microscopul electronic, s-a demonstrat ca ribosomii au forma complexa, cea sferica perceputa in mod obisnuit fiind rezultatul examinarii cu sisteme optice cu putere redusa de rezolutie. Subunitatea 50S are o forma asemanatoare cu aceea a unui fotoliu, iar subunitatea 30S se aseamana cu o haltera asimetrica, asezata orizontal pe bratele si spatarul fotoliului. Intre cele doua subunitati ramane un spatiu prin care trece ARNm (fig. 29).
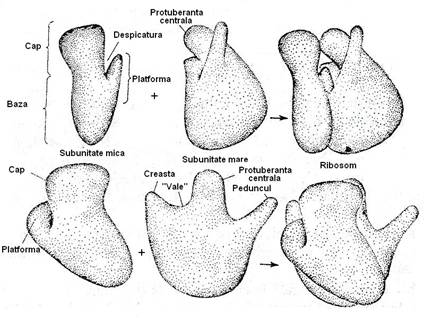
Fig. 29. Reprezentarea schematica a subunitatilor ribosomale 30S si 50S.
Din punct de vedere biochimic, ribosomii bacterieni contin circa 55 de tipuri de molecule proteice si trei tipuri de molecule de ARNr.
Studiile privind structura functionala a ribosomilor s-au facut cu doua metode foarte sensibile: difractia cu neutroni si imunoelectronomicroscopia
Subunitatea mica 30 S cuprinde 2l tipuri de molecule proteice (S1-S21), in ordinea descresterii marimii, cu gr. mol. intre 60 kDa si 8 kDa si o molecula de ARNr l6S, alcatuita din circa l6 000 nucleotide.
Subunitatea mare, 50S contine 34 tipuri de molecule proteice (Ll - L34, L= Large), cu gr. mol. cuprinsa intre 9-28,5 kD si doua molecule de ARNr, de 23 S si respectiv 5 S. Cele doua tipuri de molecule de ARN provin prin clivarea unui precursor comun, de 30 S.
ARNr este transcris din 7 operoni (rrn), fiecare cu doi operoni in tandem (P1 si P2), din care sunt transcrise copiile 16S, 23S si 5S.
Sinteza ARNr este controlata de concentratia aminoacizilor in celula: rata sintezei ARNr este dependenta de rata de aprovizionare cu aminoacizi, corelata direct cu capacitatea ribosomilor de a consuma aminoacizii in sinteza proteinelor. Daca un singur aminoacid lipseste, sinteza proteinelor ribosomale este stopata si celula nu mai asambleaza ribosomi.
Cele 55 de tipuri de proteine ribosomale se gasesc intr-un singur exemplar (o singura molecula din fiecare tip). Unele au rol structural, fiind esentiale pentru asamblarea ribosomului, altele au rol functional, permitand legarea ARNm in procesul traducerii si sintezei lantului proteic. La E. coli, in fiecare subunitate ribosomala, raportul ARN-proteine este 2/l, iar la alte bacterii, raportul este 2/3.
Moleculele componente au o distributie fixa, riguroasa in structura ribosomului. ARNr este pliat intr-o structura tridimensionala ce formeaza regiunea centrala a ribosomului si determina aspectul sau. Proteinele sunt localizate in general la suprafata ribosomului, in depresiunile pe care le creeaza ARN pliat. Unele proteine contin domenii globulare, localizate la suprafata, ce trimit extensii in regiunea centrala a ribosomului. Interactiunile fixe ale componentelor conditioneaza procesele de autoasamblare a ribosomilor.
Rata sintezei proteinelor ribosomale este controlata negativ de proteinele-r libere: acumularea proteinelor-r libere diminua rata traducerii, scade timpul de injumatatire al ARNm si al proteinelor-r. Astfel scade rata sintezei proteinelor-r.
Asamblarea ribosomilor. Experimental, componentele ribosomale se disperseaza si se autoasambleaza dupa restabilirea conditiilor de mediu, pentru a produce ribosomi activi. S-a reconstituit subunitatea 30 S, dar reasamblarea subunitatii 50 S este mai complexa deoarece este dependenta de temperatura (60o) si proteinele se denatureaza la aceasta temperatura. In studiile experimentale s-au utilizat ribosomi de Bacillus stearothermophilus, care are proteine termostabile, rezistente la 60o. Ulterior s-a reasamblat subunitatea 50 S de la E. coli. Reasamblarea urmeaza o cale specifica: anumite proteine se leaga de ARN si complexul este recunoscut succesiv de alte proteine, pana ce structura devine completa. Ribosomii reconstituiti sunt functionali (fac sinteza proteica).
Rolul proteinelor pare a fi de stabilizare a ARN, dar ele permit schimbarea configuratiei ARNr, necesare catalizei sintezei proteinelor.
Ribosomii reprezinta componenta esentiala a sistemului de traducere a informatiei genetice. Ei sunt adevaratele "fabrici" de proteine ale celulei. Ribosomii au rolul de a mentine atat molecula de ARNm, cat si complexul aminoacil-ARNt, intr-o orientare corespunzatoare pentru a permite atat citirea mesajului, cat si formarea legaturilor peptidice.
Ribosomii se asociaza in polisomi (poliribosomi), adica grupari functionale formate din 4-50 unitati ribosomale. Dimensiunile polisomilor variaza in functie de lungimea ARNm. Polisomii sintetizeaza concomitent mai multe molecule proteice pe aceiasi molecula de ARNm.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1573
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved