| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Viroizii
Viroizii reprezinta o categorie specifica de agenti infectiosi subvirali, patogeni exclusiv pentru plante, caracterizati printr-un genom alcatuit din ARN pur, prin absenta capsidei proteice si a stadiului de virion.
Denumirea de "viroid" (cu aspect de virus) este neadecvata, deoarece genomul lor nu este niciodata protejat de un invelis proteic.
Viroizii s-au descoperit accidental (Diener, l97l), cu ocazia incercarilor de caracterizare a agentului patogen al bolii tuberculilor fusiformi la cartof, considerata pana atunci ca fiind de origine virala. Viroizii cunoscuti in prezent, au luat numele maladiei pe care o produc: viroidul tuberculilor fusiformi la cartof, al nanismului si marmorarii clorotice a crizantemelor, al nanismului la hamei etc. Cel mai studiat este viroidul care produce boala tuberculilor fusiformi la cartof.
Morfologie si structura moleculara. Viroizii sunt molecule de ARN pur. La microscopul electronic, ei apar sub forma unor structuri lineare, ca niste bastonase, usor curbate, lungi de circa 50 nm, cu o grosime de 2,0 - 2,5 nm, izolate sau grupate in agregate compacte. Greutatea lor moleculara este cuprinsa intre 75 - l25 kDa.
Structura moleculara a viroidului care produce boala tuberculilor fusiformi la cartof, este cu totul neobisnuita, fiind reprezentata de molecule de ARN mononatenar, circulare, inchise covalent, alcatuite din 359 ribonucleotide, a caror structura primara (secventa) a fost determinata cu exactitate.
Structura secundara, de dubla helice defectiva, este de asemenea foarte caracteristica. Datorita gradului ridicat de complementaritate intramoleculara, un numar mare de baze (244, respectiv 68%) sunt legate in perechi. Ele formeaza regiuni dublu catenare (rezultate din imperecherea intracatenara a unei secvente de 4-5 baze), care alterneaza cu regiuni mai scurte monocatenare (2-3 baze), sub forma unor bucle interne si terminale, corespunzatoare regiunilor lipsite de baze complementare, care raman separate. Acest tip de structura, numita "in ac de par", impune anumite constrangeri topologice, datorita carora molecula de ARN sufera o pliere tridimensionala. Dupa denaturare cu formaldehida, la 63o, legaturile de H dintre bazele complementare se rup si la microscopul electronic se observa molecule circulare monocatenare cu lungimea de l40 nm si molecule lineare monocatenare cu lungimea medie de ll0 nm.
Datorita dimensiunilor mici, viroizii au rezistenta deosebita la agentii fizici (radiatii) si chimici.
Mecanismul replicarii ARN viroidal nu este cunoscut (fig. 108). Cantitatea de informatie genetica a ARN viroidal este foarte mica (teoretic suficienta numai pentru a codifica o proteina alcatuita din ll9 aminoacizi). Experientele cu extracte acelulare de plante si bacterii au evidentiat ca ARN viroidal nu are capacitatea de codificare, datorita absentei situsurilor de legare ribosomala (absenta codonului initiator AUG) si datorita structurii secundare stabile si circularitatii moleculei. ARN viroidal nu functioneaza ca ARNm. Absenta capacitatii de codificare a ARN viroidal este confirmata de faptul ca din celulele vegetale infectate nu s-a purificat nici o proteina straina de proteinele plantei, care sa sugereze originea viroidala.
Replicarea viroizilor este, in consecinta, total dependenta de celula, reprezentand o forma aparte de parazitism absolut, distincta de tipul celei descrise pentru virusuri.
Au fost propuse mai multe modele ipotetice, bazate pe cunostiintele actuale de biologie moleculara, implicand activitatea unor enzime diferite.
Modelul lui Branch si Robertson (l984) este cel mai probabil (fig. 106), deoarece se bazeaza pe evidentierea concomitenta, in celulele infectate, a doua tipuri de molecule de ARN, respectiv, molecule tipice de ARN viroidal circulare (notate conventional "+" ) si molecule de ARN multimere lineare, cu o secventa complementara (notate "-").
Modelul considera ca moleculele multimere de ARNc sunt intermediari de replicare, iar forma circulara a viroizilor maturi, sugereaza ca replicarea s-ar face dupa modelul cercului rotativ, sub controlul ARN-polimerazei II celulare, a carei legare de extremitatile viroizilor a fost evidentiata prin microscopie electronica. Rezulta molecule lineare multimere de sens antigenomic. Acestea sunt transcrise in molecule multimere complementare de sens genomic, ulterior prelucrate prin clivaj la situs specific. Dupa legarea extremitatilor se formeaza molecule circulare mature. In mod surprinzator, activitatile enzimatice de clivare si legare a extremitatilor, se desfasoara in absenta proteinelor enzimatice si se se aseamana cu mecanismul autoexciziei intronilor din ARN premesager al organitelor. ARN are activitate enzimatica - ribozimica.
Deoarece, in mod normal, ARN-polimeraza II celulara foloseste ca matrita o molecula de ADN, autorii considera ca, datorita structurii lor unice, prezenta viroizilor in celule ar fi interpretata gresit de enzima, ca o molecula de ADN. Dupa acesti autori, viroizii ar fi agenti similari ADN (ADN-like).
Dupa alti autori, replicarea moleculei de ARN viroidal este autocatalitica. O molecula de ARN care se replica autocatalitic se numeste ARN-replicaza.
Replicarea viroizilor are loc in nucleul celulelor si, ca urmare, infectiozitatea maxima este asociata cu nucleul si, in special, cu cromatina celulelor infectate.
In functie de faza infectiei, numarul viroizilor variaza intre 200 si l0 000/celula.
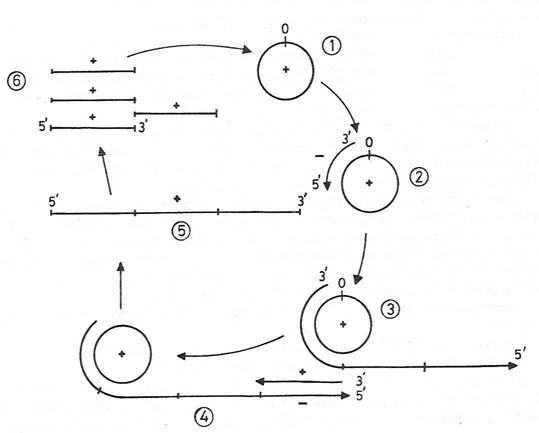
Fig. 106. Replicarea viroizilor dupa modelul lui Bransch si Robertson (1984). Viroidul matur (+) (1) are rol de matrita (2) pentru sinteza unor molecule multimere de ARN complementar (3). Acestea sunt transcrise in molecule multimere (+) (4), reprezentate de genomuri viroidale concatemere (5). Clivarea lor riguroasa (6) are ca rezultat formarea moleculelor monomere de ARN viroidal, cu secvente terminale caracteristice. Circularizarea lor genereaza ARN viroidal progen.
Patogenitatea viroizilor
Desi cantitatea de informatie genetica pe care o aduc in celula este foarte mica, viroizii produc mai mult de l0 maladii grave ale plantelor de cultura sau ornamentale, cu importanta economica. Intre acestea, cele mai studiate sunt: boala tuberculilor fusiformi la cartof, nanismul si marmorarea clorotica a crizantemelor, nanismul hameiului, boala cadang-cadang a cocotierului si o boala caracteristica a portocalilor si lamailor, insotita de distrugerea progresiva a scoartei si scaderea masiva a productiei de fructe.
Infectia experimentala reproduce tabloul manifestarilor patologice la plante din aceiasi specie, datorita replicarii excesive a acestor entitati moleculare cu caracter infectios. ARN viroidal care produce boala tuberculilor fusiformi la cartof, se propaga la un numar foarte mare de plante din familia Solanaceae, dar si la plante din alte familii, unde se replica fara sa produca simptomele imbolnavirii.
Viroizii produc infectii persistente: la cartof, simptomele apar in perioada formarii tuberculilor, iar la tomate, la 2-3 saptamani de la infectie. In frunzele de tomate si in petiol, dupa infectia cu ARN viroidal al tuberculilor fusiformi de cartof se produce o hipertrofie marcata a nucleului. Se intensifica curentii citoplasmatici. In tesutul infectat, infectiozitatea este asociata cu fractia sa nucleara, ceea ce denota ca ARN viroidal este asociat cu nucleul si in special cu cromatina celulelor infectate.
Mecanismul producerii bolilor viroidale este greu explicabil, deoarece ARN viroidal nu codifica sinteza unor proteine noi. |esuturile plantelor infectate nu contin proteine noi, in raport cu plantele sanatoase, dar prezinta variatii cantitative foarte importante ale unor proteine normale. Este probabil ca viroizii determina o perturbare a mecanismelor de reglare a exprimarii anumitor gene ale celulei. Viroizii nu ar actiona prin exprimarea propriei lor informatii genetice, ci prin perturbarea functiei genelor celulare, a carei consecinta ar fi alterarea mecanismelor normale de reglare a functiilor celulei.
In unele cazuri, s-a observat o scadere pronuntata a cantitatii de gibereline in tesuturile plantelor infectate. Deoarece acidul giberelic poate regla evenimentele nucleare ale transcrierii, precum si metabolismul unor specii de ARN, replicarea viroizilor si/sau patogenitatea lor ar putea fi influentate prin intermediul acestei clase de mediatori hormonali.
Tulburarile de crestere, frecvent asociate cu infectiile viroidale, pot fi astfel explicate ca rezultat al unui dezechilibru al sintezei si activitatii hormonilor de crestere: viroizii prezenti in nucleul celulelor, ar actiona ca molecule anormale de reglare, care interfera cu cele care asigura in mod normal reglarea genelor ce codifica hormonii de crestere.
Transmiterea viroizilor pe orizontala, de la o planta bolnava, la plantele sanatoase se realizeaza in mod obisnuit, pe cale mecanica (de exemplu, prin altoire sau prin alte leziuni mecanice), probabil prin intermediul nucleilor celulari sau al unor fragmente de cromatina, care contin ADN viroidal. Conservarea infectiozitatii viroizilor in cursul transmiterii, in fazele extracelulare, adica pastrarea infectioziattii lor in absenta invelisului proteic (capsidal) este datorata localizarii lor intranucleare, la adapost de atacul enzimatic.
Transmiterea viroizilor pe verticala, se face prin polenul infectat, al plantelor de tomate sau cartof.
Originea viroizilor este necunoscuta. Au fost propuse mai multe ipoteze.
l. Ipoteza originii virale a fost formulata in doua variante:
a) Viroizii ar fi entitati virale foarte primitive, care nu au ajuns la gradul de complexitate structurala si genetica, necesare pentru a induce in celula, reactiile metabolice noi, de biosinteza, pentru a codifica sinteza proteinelor specifice, capabile sa asigure propria lor replicare;
b) Viroizii ar reprezenta entitati virale degenerate, a caror biosinteza si replicare a fost initial determinata de infectia cu ARN viral, care a suferit o reducere importanta a genomului (inclusiv a genelor care codifica proteinele capsidei) si a capatat o autonomie totala.
2. Ipoteza originii celulare considera viroizii ca provenind din acizii nucleici ai celulei:
a) Viroizii ar putea proveni din transcrierea unor gene cromosomale patologice, sub influenta unor factori inca nedeterminati, sau ar fi derivati dintr-o specie de ARN cu greutate moleculara mica, de genul celor prezente in nucleu si nucleol, implicate in mod normal in reglarea unor functii nucleare si in transcrierea ARN;
b) Viroizii ca derivati ai elementelor genetice transpozabile (EGT Ipoteza se bazeaza pe observatia ca viroizii contin secvente care se aseamana cu cele prezente la extremitatile EGT. Viroizii ar fi evoluat din EGT sau din provirusuri retravirale, din care, in cursul transcrierii la ARN, ar fi disparut unele secevente de baze;
c) Ipoteza originii viroizilor prin circularizarea intronilor, se bazeaza pe faptul ca genele celulelor eucariote, ca si cele ale virusurilor animale, au o structura discontinua, in sensul ca sunt formate din secvente cu rol de codificare (exoni), care prin transcriere sunt regasite in structura ARN matur si secvente intercalate (introni), care sunt clivate in etapa de prelucrare a ARN premesager.
In cursul procesului de biosinteza a proteinelor, intreaga gena (exonii si intronii) este transcrisa la o molecula mare de ARN premesager. Ulterior, acesta sufera un proces de maturare, prin care intronii sunt indepartati de o enzima speciala, iar exonii sunt "lipiti" sau inaditi cap la cap, pentru a forma o molecula de ARN matur, care trece in citoplasma si este tradusa intr-o secventa polipeptidica.
Sectionarea intronilor este controlata de o molecula mica de ARN (ARN U1), prezenta in nucleii celulelor vegetale, partial omolog cu intronii, care se leaga de extremitatile acestora, asigurand excizia corecta. Final, intronii se circularizeaza sub controlul unei ARN-ligaze). Circularizarea impiedica degradarea enzimatica, confera posibilitatea de replicare necontrolata si de infectiozitate, prin mobilitate intercelulara si intre organisme.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1940
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved