| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
DOCUMENTE SIMILARE |
|
ANGLIAVANDENIAI (KARBOHIDRATAI, SACHARIDAI)
Angliavandeniai – polifunkciniai junginiai, aktyviai dalyvaujantys žmogaus, gyvűnř ir augalř gyvybinëse funkcijose.
Angliavandeniai sudaro 80 sausosios augalř masës ir 2 sausosios gyvűnř masës.
Gyvűnř organizmai nesugeba sintetinti karbohidratř. Tik augalai sugeba juos sintetinti iš atmosferos CO2 ir H2O, vykstant sudëtingai reakcijai – fotosintezei:
![]()
Gyvűnai karbohidratř gauna su augaliniu maistu ir suvartoja energijai išgauti:
![]()
Tokius junginius vadinti karbohidratais pasiűlë Šmitas (1844 m.) pagal analizës duomenis Cn(H2O)m.
Karbohidratai skirstomi á monosacharidus, oligosacharidus (2 …10 monosacharidř fragmentř) ir polisacharidus (daugiau 10 monosacharidř fragmentř).
8.1. Monosacharidř struktűra, izomerija ir savybës
Monosacharidai – kietos, gerai tirpios vandenyje, dažniausiai saldžios medžiagos.
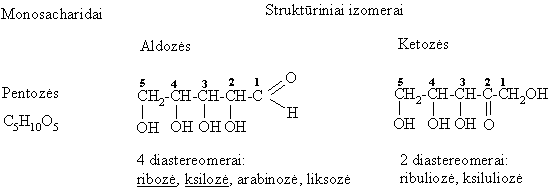
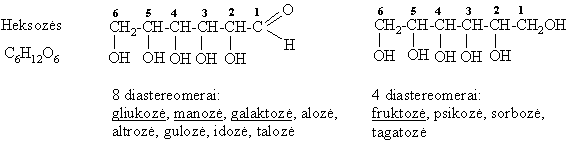
Pagal C atomř skaičiř monosacharidai skirstomi á triozes (3 C), tetrozes (4 C), pentozes (5 C), heksozes (6 C) ir t.t.
Nesisteminiai monosacharidř pavadinimai baigiasi galűne – ozë
Monosacharidř molekulëse yra okso- ir hidroksigrupiř, todël jie yra polihidroksialdehidai (aldozës) arba polihidroksiketonai (ketozës).
Monosacharidams kaip polifunkciniams junginiams bűdinga ávairi izomerija.
Struktűrinë (funkciniř grupiř) izomerija
![]()
Erdvinë izomerija
Monosacharidai turi vieną arba kelis chiralinius centrus ir yra optiškai veiklios medžiagos.
Monosacharidř erdviniai izomerai, nesantys vienas kito veidrodinis atspindys bei besiskiriantys fizikinëmis savybëmis, vadinami diastereomerais. Kiekvienas monosacharidř diastereomeras turi trivialřjá pavadinimą. Iš aldopentoziř reikšmingiausi diastereomerai yra ribozë ir ksilozë, o iš heksoziř organizme dažniausiai aptinkami šie diastereomerai: gliukozë, manozë, galaktozë (aldoheksozës) ir fruktozë (ketoheksozë).
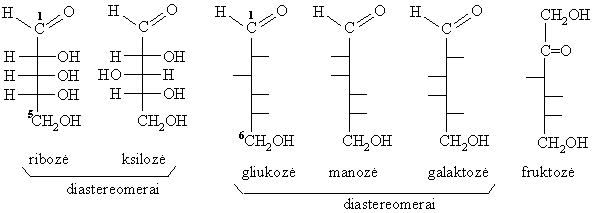
Kiekvienas diastereomeras gali egzistuoti dviejř optiniř izomerř, vadinamř D- ir L-enantiomerř, formoje. Jeigu gamtinës aminorűgštys yra L-formos, tai gamtiniai monosacharidai yra D-formos.
Žiedo grandinës tautomerija (oksociklotautomerija)
Išvardintř gamtiniř monosacharidř C-4 arba C-5 (fruktozës atveju C-5 arba C-6) padëties hidroksigrupei nukleofiliškai jungiantis prie oksogrupës (žiűr. 62 psl.) susidaro hemiacetalinis tiltelis ir naujas, vadinamasis hemiacetalinis arba glikozidinis, hidroksilas.

Ciklinëje struktűroje C-1 atomas tampa chiraliniu centru, vadinamu anomeriniu anglies atomu, arba anomeriniu centru. Hemiacetalinë gliukozë yra dviejř diastereomeriniř a- ir b-formř, vadinamř anomerais. a-Anomero hemiacetalinë hidroksigrupë Fišerio projekcijoje yra toje pačioje pusëje kaip ir paskutinio chiralinio centro hidroksigrupë (laisva arba áeinanti á ciklą ir nusakanti priklausomybę D- arba L-genetinei eilei). b-Anomere jos stovi priešingose pusëse.
Pentoziř ir heksoziř tirpaluose pusiausvyra iš esmës nusistovi tarp penkiř tautomerř. Cikliniai tautomerai virsta vienas kitu tik per necikliná tautomerą. Reiškinys vadinamas oksociklotautomerija.
Konformacijos
Piranozës egzistuoja këdës konformacijoje, kurioje kuo daugiau stambiř pakaitř yra ekvatorinëse padëtyse.
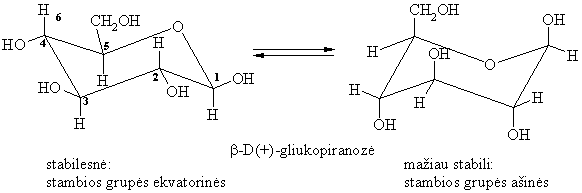
Enantiomerai (optiniai izomerai)
Monosacharidř erdviniai izomerai, esantys vienas kito veidrodinis atspindys, vadinami enantiomerais (optiniais izomerais). Enantiomerams bűdingos identiškos fizikinës ir cheminës savybës, išskyrus vieną. Jie suka poliarizuotą šviesos plokštumą vienodos reikšmës, tačiau priešingo ženklo kampu.
Kiekvienas monosacharido diastereomeras (ciklinis ar neciklinis) turi du enantiomerus: dešiniojo sukimo (+) ir kairiojo sukimo (-). Enantiomerai tradiciškai skirstomi á dvi D- ir L-genetines eiles. D-eilei priskiriami tie enantiomerai, kuriř paskutiniojo chiralinio centro (labiausiai nutolusio nuo oksogrupës) konfigűracija yra tokia pati, kaip ir konfigűracinio standarto D-glicerolio aldehido. L-eilës enantiomerř paskutinio asimetrinio C konfigűracija sutampa su L-glicerolio konfigűracija.
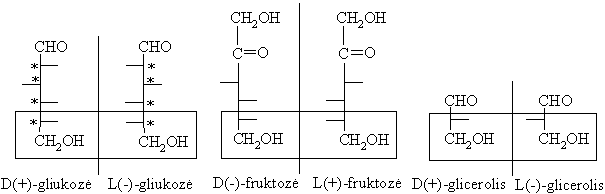
Realus dešinysis (+) arba kairysis (-) sukimo kampas priklauso nuo visř molekulës chiraliniř centrř, o ne vien nuo paskutiniojo asimetrinio C atomo konfigűracijos ir nustatomas eksperimentiškai.
Daugumoje atvejř biologiniu aktyvumu pasižymi tik vienas monosacharidř enantiomeras (D-eilës).
Mutarotacija
Ištirpinus a-D-gliukopiranozę vandenyje, tirpalo sukimo kampas mažëja ir, pasiekęs +52.5 C, toliau nebekinta. b-Formos sukimo kampas didëja, kol pasiekia taip pat ribinę +52.5 C reikšmę. Šis reiškinys, t.y. poliarizacijos plokštumos sukimo kampo kitimas, buvo pavadintas mutarotacija. Jos esmë ta, kad tirpale vienas anomeras virsta kitu, ir procesas vyksta tol, kol nusistovi pusiausvyra.

Monosacharidř dariniai
Gamtoje aptinkamř monosacharidř dariniř molekulëse vietoje vienos ar keliř hidroksigrupiř gali bűti tokie pakaitai: H, NH2, SH ir kt.
Deoksisacharidai
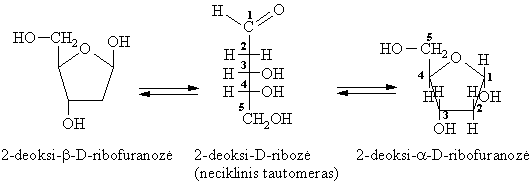
Deoksisacharidř cheminës savybës panašios á monosacharidř.
Aminosacharidai (glikozaminai)
Aminosacharidai pagal aminogrupës padëtá skirstomi á 2-amino-, 4-amino- ir 2,6-diaminosacharidus. Gamtoje labiausiai paplitusios N-acetildariniř pavidale 2-aminogliukozë ir 2-aminogalaktozë.
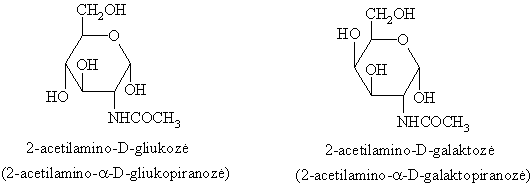
Aminosacharidams bűdingos monosacharidř ir aminř savybës.
Monosacharidř ir jř dariniř cheminës savybës
Monosacharidams bűdinga oksociklotautomerija. Cheminiuose virsmuose gali dalyvauti kiekvienas tautomeras atskirai arba ávairiř tautomerř visuma.
Rűgštinës-bazinës savybës
Monosacharidai yra polihidroksiliniai alkoholiai. Panašiai kaip glicerolis (pKa 14.0) vandeniniuose tirpaluose jie nepasižymi rűgštinëmis savybëmis. Tačiau alkoholiř tirpaluose veikiami šarminiř metalř alkoksidais monosacharidai sudaro druskas – sacharatus, kurios vandeniniuose tirpaluose visiškai hidrolizinasi:
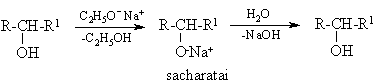
Veikiamos šarmř (netgi vandeniniř tirpalř) protonř donorais gali bűti CH-grupës, esančios greta karbonilgrupës (žiűr. epimerizacija, 142 psl).
Monosacharidř esteriai su sieros arba fosforo rűgštimis – stiprios rűgštys. Bet kurio monosacharido fosfatai R-OPO(OH)2 yra divandenilinës rűgštys ir disocijuoja laipsniškai; pK1a 0.6 –1,6 ir pK2a 5.5 – 6.5. Biosistemose pagal pirmąją pakopą jos yra visiškai jonizuotos, o kai pH = 7 didžia dalimi jonizuotos ir pagal antrąją pakopą. Monosacharidř monosulfatai ROSO3H – dar stipresnës rűgštys (pKa < 0.4), todël biologinëse terpëse jř sulfogrupë visada jonizuota. Monosacharidř fosfatř ir sulfatř anijonai yra susikaupę ląstelës vidaus skysčiuose ir, skirtingai negu monosacharidai, nepraeina per ląstelës membraną.
Aminosacharidai R-NH2 dëka aminogrupës pasižymi bazinëmis savybëmis.
Kompleksodara
Monosacharidai pasižymi polihidroksiliniř alkoholiř savybëmis. Jie reaguoja su Cu(OH)2 sudarydami kompleksinius junginius:
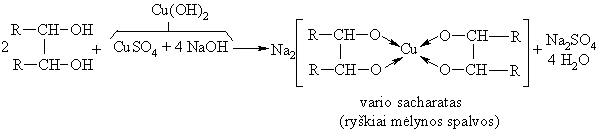
Monosacharidř fosfatai ir sulfatai yra žymiai aktyvesni ligandai už monosacharidus. Jie sudaro mažai patvarius kompleksus su K+ ir Na+ katijonais ir patvaresnius su Mg2+ bei Ca2+ katijonais. Susidarant kompleksiniams junginiams aktyvinami biosubstratai. Antra vertus, tokie biosubstratai reguliuoja mineraliniř druskř apykaitą organizme.
Elektrofilinës-nukleofilinës reakcijos
Elektrofilinë-nukleofilinë reakcija yra monosacharidř žiedo grandinës tautomerijos (oksociklotautomerijos, žiűr. 136 psl.) pagrindas. Cikliniř hemiacetaliř glikozidinë hidroksigrupë aktyviai reaguoja su acilinimo bei alkilinimo reagentais (žiűr. alkoholiř O-acilinimą bei O-alkilinimą, 51 psl.).
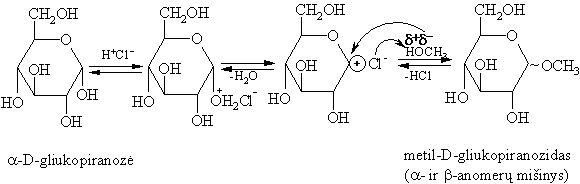
Glikozidai, skirtingai negu eteriai, lengvai hidrolizinasi rűgštinëje terpëje.
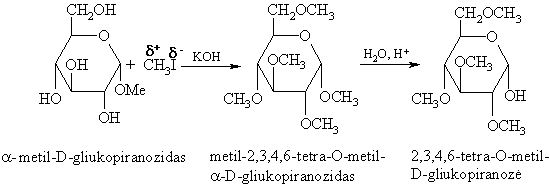
Glikozidř hidroksigrupës yra O-nukleofilai ir reaguoja su stipriais alkilinimo reagentais.
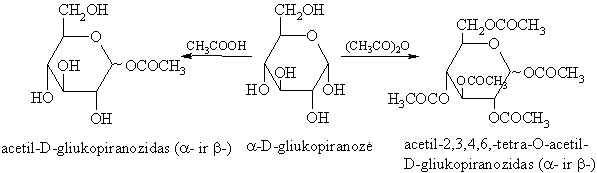
Monosacharidř hidroksigrupës, veikiamos karboksirűgštimis arba jř anhidridais, acilinasi.
Biosistemose monosacharidai, reaguodami su adenozintrifosfatu (ATP), fosforilinasi:
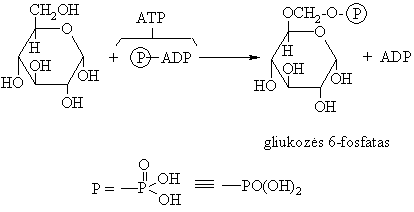
Oksidacijos-redukcijos reakcijos
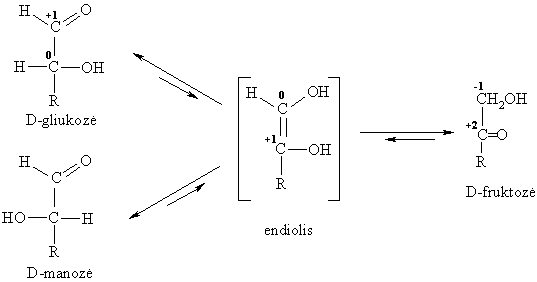
Monosacharidř C atomř oksidacijos laipsniř reikšmës kinta nuo –1 iki +2. Todël jie lengvai dalyvauja oksidacijos-redukcijos reakcijose, áskaitant ir vidinę molekulinę dismutaciją.
Monosacharidř epimerizacija
Epimerais vadinami diastereomerai, besiskiriantys tik vieno C atomo konfigűracija molekulëje. Jř virtimo vienas kitu cheminis procesas vadinamas epimerizacija.
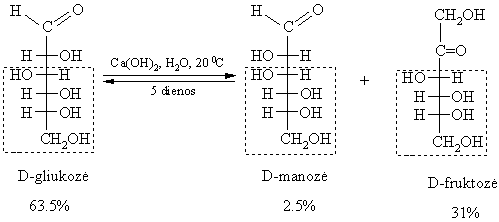
Epimerizacija šarminëje terpëje:
Organizme fermentas fosfogliukoizomerazë iš gliukozës 6-fosfato epimerizacijos virsmu gamina fruktozës 6-fosfatą (2-oji gliukozës katabolizmo stadija).
Aldolinis C3-C4 skilimas
Gyvűnř organizmuose, veikiant fermentui aldolazei D-fruktozës 1,6-difosfatas skyla sudarydamas dihidroksiacetono fosfato ir D-glicerolio aldehido 3-fosfato mišiná.
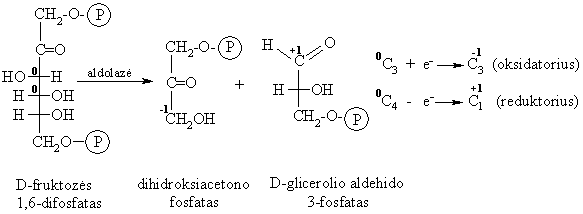
Ši reakcija yra 3-oji gliukozës katabolizmo stadija. Augalř ląstelëse vyksta atvirkštinë reakcija (viena iš fotosintezës stadijř), kurios metu susidaro D-gliukozës 1,6-difosfatas.
Monosacharidř oksidavimas
Švelnűs oksidatoriai, pvz. bromo vanduo, oksiduoja oksogrupę, o stiprűs, pvz. konc. HNO3, oksiduoja ne tik oksogrupę, bet ir pirminę hidroksigrupę.
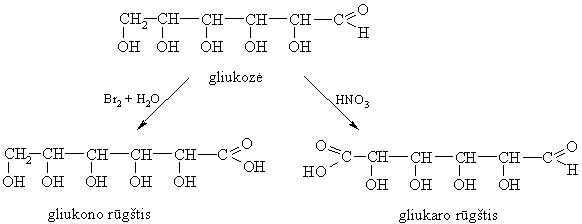
Oksiduojant glikozidus
oksiduojasi tik pirminë hidroksigrupë, virsdama karboksigrupe.
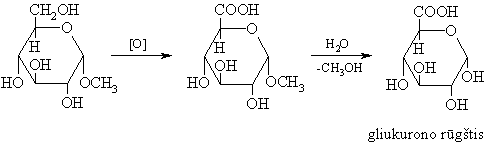
Organizme gliukurono rűgšties glikozidinë hidroksigrupë nukleofiliškai keičiama svetimkűnio –OR arba –NHR grupëmis ir taip dalyvauja jo šalinimo iš organizmo procese.
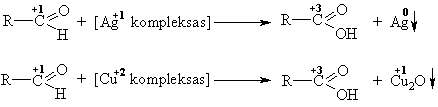
Šarminëje terpëje oksogrupę oksiduoja metalř katijonai (Ag+1, Cu+2), kurie redukuojasi iki laisvo metalo (Ag) arba mažesnio oksidacijos laipsnio jono (Cu+1).
Monosacharidř hidrinimas
Reduktoriai hidrina
monosacharidus iki polihidroksialkoholiř. Hidrinantis fruktozës
oksogrupei, antrasis ketozës molekulës anglies atomas
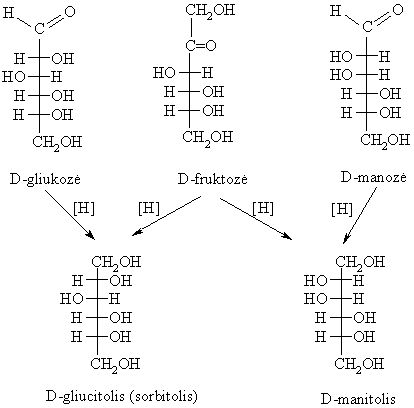
Daugiahidroksiliniai alkoholiai sudaro esterius su acto rűgštimi, kurie taikomi praktikoje. Pvz., D-gliucitolio heksanitratas (nitrosorbitas) ir D-manitolio heksanitratas taikomi kraujagyslëms plësti. Pramonëje iš gliukozës gaminama askorbo rűgštis (vitaminas C) – stiprus reduktorius. Oksiduodamasi virsta dehidroaskorbo rűgštimi. Šis procesas yra grážtamas:
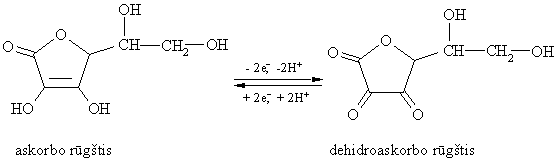
j10 = 0.08 V
Organizme konjuguota oksidacinë redukcinë sistema – askorbo rűgštis ir dehidroaskorbo rűgštis – yra efektyvus radikaliniř oksidaciniř redukciniř procesř, pasireiškiančiř organizmo patologinëse bűsenose, priešnuodis.
Glikolizë – gliukozës katabolizmas
Dauguma karbohidrtř virškinamąjame trakte hidrolizinasi iki gliukozës ir fruktozës, kurios patenka á ląstelę. Tolimesnis monosacharidř virtimas piruvo rűgštimi vyksta glikolizës proceso, susidedančio iš 10 reakcijř, metu.
Glikolizës procese išskiriami trys etapai: I etapas – heksoziř kitimas, II etapas – trioziř kitimas, III etapas – oksokarboksirűgščiř kitimas.
I etapas
Gliukozës fosforilinimas (aktyvinimas)
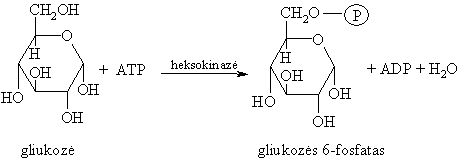
Tai elektrofilinë-nukleofilinë reakcija, endenerginë ir negrážtamoji.
Gliukozës 6-fosfato izomerinimas
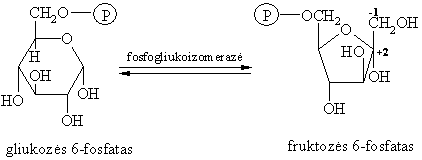
Tai vidinës molekulinës dismutacijos reakcija.
Fruktozës 6-fosfato fosforilinimas
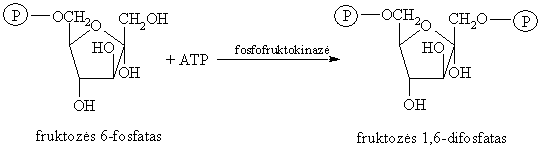
Tai elektrofilinë-nukleofilinë reakcija, negrážtamoji ir pati lëčiausia iš visř glikolizës reakcijř.
Fruktozës 1,6-difosfato aldolinis skilimas
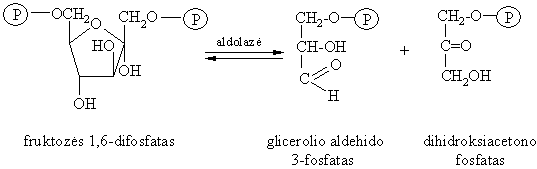
Tai vidinës molekulinës dismutacijos (oksidacijos-redukcijos) reakcija.
II etapas
Dihidroksiacetono fosfato izomerizavimas (epimerizavimas)

Tai vidinës molekulinës dismutacijos (oksidacijos-redukcijos) reakcija.
Oksidavimas ir fosforilinimas
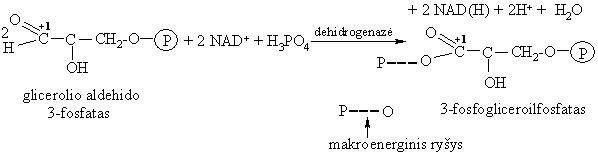
Tai tarpmolekulinës dismutacijos ir fosforilinimo reakcijos. Susidariusi anhidridinë jungtis – makroenerginë.
3-Fosfogliceroilfosfato hidrolizë
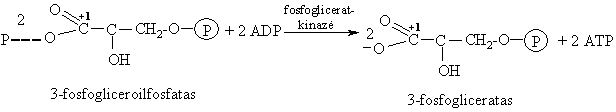
Tai elektrofilinë-nukleofilinë reakcija, ekzoenerginë.
3-Fosfoglicerato izomerizavimas

Tai elektrofilinë-nukleofilinë reakcija.
III etapas
2-Fosfoglicerato dehidratavimas
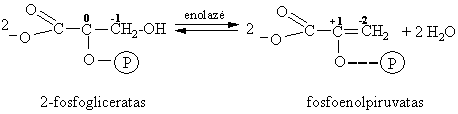
Tai vidinës molekulinës dismutacijos (oksidacijos-redukcijos) reakcija susidarant makroenerginiam ryšiui.
Rűgštinë fosfoenolpiruvato hidrolizë iki piruvato

Tai vidinës molekulinës dismutacijos (oksidacijos-redukcijos) reakcija, negrážtamoji, egzoenerginë.
Taigi, dviejose I etapo reakcijose sunaudojama viena ATP molekulë, o III etape – išsiskiria dvi ATP molekulës. Vadinasi, glikolizës metu ląstelë pasipildo energija.
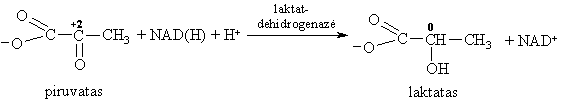
Tolimesni piruvato virsmai organizme priklauso nuo sąlygř, kuriomis jie vyksta. Anaerobinëmis sąlygomis raumenyse ir audiniuose piruvatas gali redukuotis á laktatą:
Kita jo kitimo kryptis – oksidacinis dekarboksilinimas:
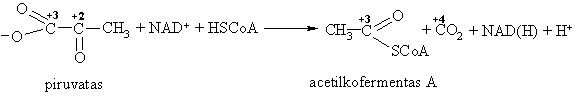
Susidaręs acetilkofermentas A mitochondrijoje patenka á Krebso ciklą, kuriame acetilgrupë oksiduojasi iki CO2 ir H2O. Esant tokiai glikolizës baigčiai iš kiekvienos gliukozës molekulës išsiskiria 38 molekulës ATP. Todël angliavandeniai ir yra vienas iš pagrindiniř ląstelës energijos šaltiniř.
Monosacharidř
katabolizmui bűdingos fermentinio skilimo, vykstančio
Alkoholinis (anaerobinis procesas):
![]()
Sviestarűgštis (anaerobinis procesas):
![]()
Pienrűgštis (anaerobinis procesas):
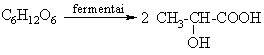
Citrinrűgštis (aerobinis procesas):
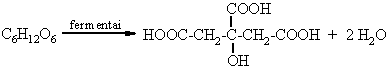
Šios rűgimo rűšys yra svarbios ávairiř maisto produktř gamyboje.
8.2. Disacharidř struktűra ir savybës
Gamtiniř disacharidř molekulës sudarytos iš dviejř vienodř arba skirtingř monosacharidř likučiř, sujungtř O-glikozidiniais ryšiais.
Svarbesni gamtiniai disacharidai yra: sacharozë (cukriniř runkeliř, cukranendriř cukrus), maltozë (salyklo cukrus), laktozë (pieno cukrus), celobiozë (celiuliozës dalinës hidrolizës junginys).
Hidrolizinant praskiestomis mineralinëmis rűgštimis arba fermentais, jie skyla á dvi monosacharido molekules:
![]()
![]()
![]()
![]()
![]()
Disacharidai skirstomi á du tipus pagal glikozidinio ryšio susidarymo pobűdá: redukuojantys ir neredukuojantys.
Redukuojantys disacharidai (glikozilglikozës)
Redukuojantys disacharidai sudaryti sureagavus vieno monosacharido hemiacetaliniam (glikozidiniam) hidroksilui su kito monosacharido alkoholiniu hidroksilu (dažniausiai 4 arba 6 padëties). Šio tipo disacharido molekulę sudarančioje vienoje monosacharido liekanoje yra laisva glikozidinë hidroksigrupë, galinti sudaryti tautomerinę pusiausvyrą su oksogrupe.Todël jie pasižymi aldoziř savybëmis: redukuoja vará ir sidabrą iš jř kompleksiniř junginiř (iš čia ir kilo pavadinimas redukuojantys), lengvai oksiduojasi iki biono rűgščiř, dalyvauja mutarotacijoje.
Gamtiniai redukuojantys disacharidai yra: maltozë, laktozë ir celobiozë.
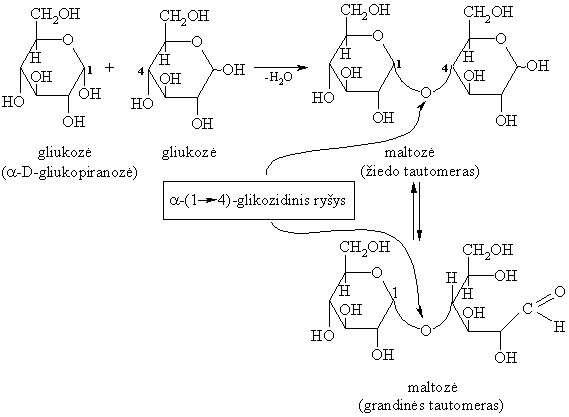
Žmogaus organizmas ásisavina maltozę.
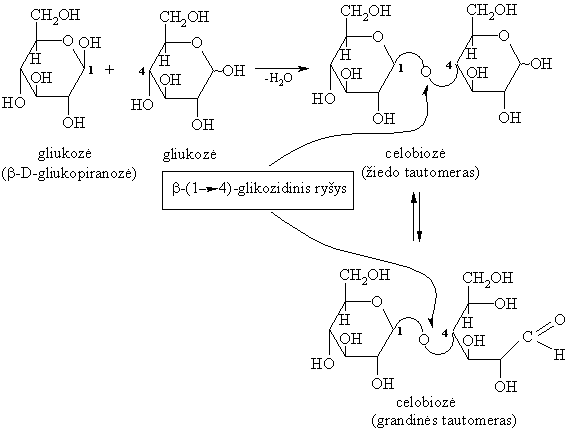
Celobiozę ásisavina žoliaëdžiř organizmas.
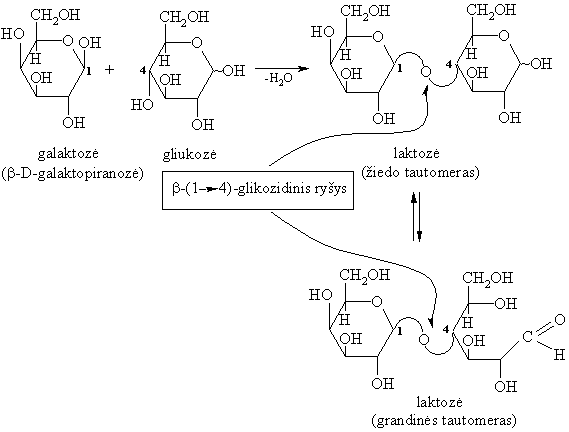
Kai kurie žmonës neásisavina laktozës, kadangi jř organizme trűksta fermento laktazës.
Redukuojantys disacharidai dalyvauja beveik visose reakcijose, kurios bűdingos monosacharidams.
Neredukuojantys disacharidai (glikozilglikozidai)
Šio tipo disacharidai sudaryti sureagavus abiejř monosacharidř glikozidinëms hidroksigrupëms. Jie neturi laisvos glikozidinës hidroksigrupës, todël jiems nebűdinga mutarotacija, jie neredukuoja oksidatoriř (iš čia kilo pavadinimas neredukuojantys).
Gamtinis neredukuojantis disacharidas yra sacharozë.
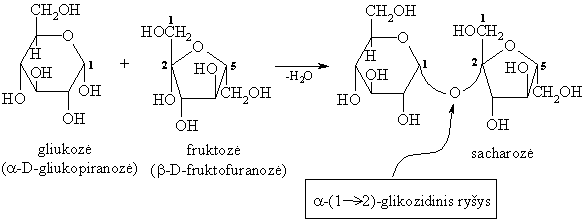
Sacharozë – augaluose labiausiai paplitęs disacharidas. Ypač daug jo yra cukriniuose runkeliuose (14-18%) ir cukranendrëse (15-20%).
8.3 Polisacharidř struktűra ir savybës
Polisacharidai (poliglikozidai) yra gamtinës stambiamolekulës medžiagos, susidariusios iš daugelio glikozidiniais ryšiais susijungusiř monosacharido likučiř.
Polisachariduose dažniausiai esti (1 4) ir (1 6) glikozidiniř ryšiř (dar bűna 1 3 arba 1 2). Polisacharidř molekulës bűna linijinës arba šakotos, susidariusios iš vienodř arba skirtingř monosacharidř. Pirmosios grupës polisacharidai vadinami homopolisacharidais, o antrosios – heteropolisacharidai.
Homopolisacharidai
Svarbiausi homopolisacharidai yra krakmolas, glikogenas, celiuliozë ir chitinas.
Krakmolas
Krakmolas yra augalř rezervinis homopolisacharidas. Krakmolas sudarytas iš dviejř homopolisacharidř: amilozës (10-20%) ir amilopektino (80-90%).
Amilozës makromolekulës yra linijinës struktűros, beveik nešakotos. Jose D-gliukopiranoziniai likučiai (200-1000) susijungę a-(1 4)-glikozidiniu ryšiu.
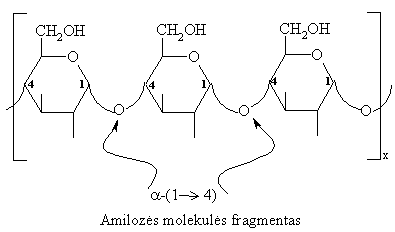
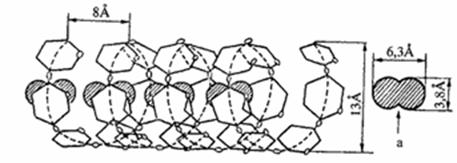
Rentgeno struktűrinës analizës duomenimis, amilozës linijinës makromolekulës susisukusios á spiralę, kurios kiekvieną viją sudaro 6 gliukozës likučiai. Vijos viduje susidaro 5Å skersmens ertmë, á kurią gali ásiterpti kitř junginiř, pvz., jodo arba alkoholio, molekulës. Pakliuvęs á tokią ertmę, jodas sąveikauja su amilozës vandenilio atomais ir sudaro mëlynos spalvos kompleksiná junginá. Reakcija su jodu daroma krakmolui atpažinti.
Amilozës makromolekulës spiralinë struktűra: užtušuota – ásiterpusiř junginiř molekulës.
Amilopektino makromolekulës skiriasi nuo amilozës didesniu polimerizuotumu ir šakotumu. Linijinës jř atkarpos palyginti trumpos, susideda iš 20-30 gliukozës likučiř. Pačios atkarpos jungiasi tarpusavyje a-(1 6)-glikozidiniu ryšiu.
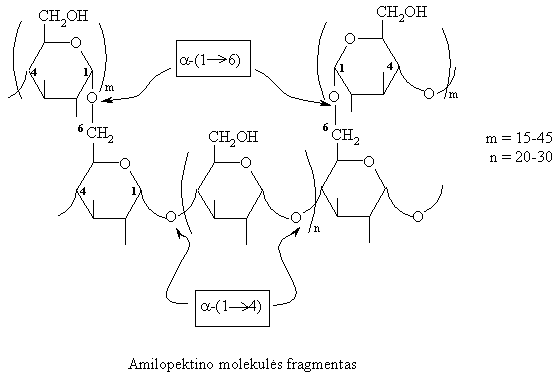
Amilopektino makromolekulës atšakos yra spiralës formos, todël su jodu sudaro violetinës spalvos kompleksiná junginá.
Krakmolą kaitinant arba hidrolizinant (rűgštinë arba fermentinë hidrolizë), jis skyla á paprastuosius sacharidus – dekstrinus, maltozę, pagaliau – gliukozę.
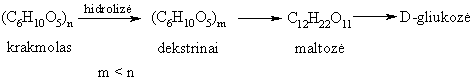
Virškinamajame trakte krakmolą hidrolizina fermentai, galintys skaldyti a-(1 4)- ir a-(1 6)-glikozidinius ryšius.
Krakmolas yra vienas iš pagrindiniř maisto produktř.
Glikogenas
Tai rezervinis gyvuliř organizmo polisacharidas. Su maistu patekę karbohidratai biocheminiř pokyčiř metu virsta glikogenu ir kaupiasi kepenyse (6-7%) ir raumenyse (iki 1%). Glikogeno makromolekulës panašiai kaip amilopektino sudarytos iš a-D-gliukopiranozës, tik dvigubai šakotesnës. Linijinës atkarpos dažniausiai sudarytos iš 10-12 gliukozës likučiř, susijungusiř tarpusavyje a-(1 4)-glikozidiniu ryšiu. Atšakos prisijungę a-(1 6)-glikozidiniu ryšiu.
Glikogeno makromolekulës neprasiskverbia pro membraną ir pasilieka ląstelëje (rezerve) tol, kol organizmui neprisireikia energijos. Glikogeno funkcija – kaupti gliukozę lengvai ásisavinama ląstelëms forma. Glikogenas labai lengvai hidrolizinasi rűgščiř ir fermentř veikiamas. Molekulës šakotumas palankus fermentinei veiklai, nes fermentas g-amilazë skaido makromolekulę nuo jos galo. Tad, juo daugiau polisacharidas turi atšakř, juo greičiau já fermentai suardo. Pagal tuos pačius dësnius vyksta ir glikogeno sintezë ląstelëje.
Celiuliozë
Tai labiausiai paplitęs polisacharidas, savo kiekiu užimąs neginčijamai pirmą vietą tarp kitř organiniř junginiř (apie 1011 t kasmet jos pagamina augalai biosintezës bűdu). Ji yra pagrindinë augaliniř ląsteliř apvalkalëliř sudedamoji dalis.
Celiuliozë yra stambiamolekulë medžiaga, sudaryta iš b-D-gliukopiranoziř, kurios yra susijungusios linijinëje grandinëje b-(1 4)-glikozidiniais ryšiais ir viena kitos atžvilgiu pasisukusios 180 kampu.
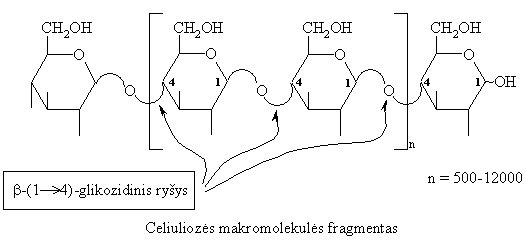
Celiuliozë - mechaniškai
atspari, chemiškai inertiška, todël ji yra
Žmogaus organizmas neturi celiuliozę skaldančiř fermentř, todël jos nevirškina (žolëdžiř gyvűnř organizmai virškina celiuliozę).
Celiuliozës hidrolizë vyksta pakopomis.
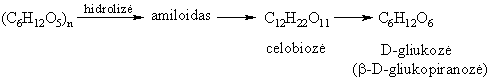
Chitinas
Tai polisacharidas, sudarytas iš N-acetil-D-gliukozamino likučiř, sujungtř b-(1 4)-glikozidiniais ryšiais. Chitino makromolekulë nešakota, erdviniu išsidëstymu panaši á celiuliozę.
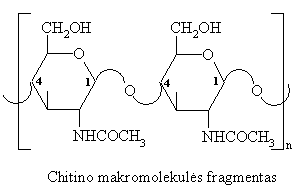
Iš chitino sudaryti vëžiagyviř kiautai, vabzdžiř išorinë danga. Chitinas – tai grybř struktűrinis polisacharidas.
Heteropolisacharidai
Jř yra jungiamajame audinyje (odoje, kremzlëse, sausgyslëse, sąnariř skystyje, kauluose).
Hialurono rűgštis
Hialurono rűgštis sudaryta iš disacharidiniř (D-gliukurono rűgšties ir N-acetil-N-gliukozamino) likučiř, sujungtř b-(1 4)-glikozidiniais ryšiais. Gliukurono rűgštis su N-acetil-O-gliukozaminu susijungusi b-(1 3)-glikozidiniu ryšiu.
Hialurono rűgšties makromolekulës fragmentas:
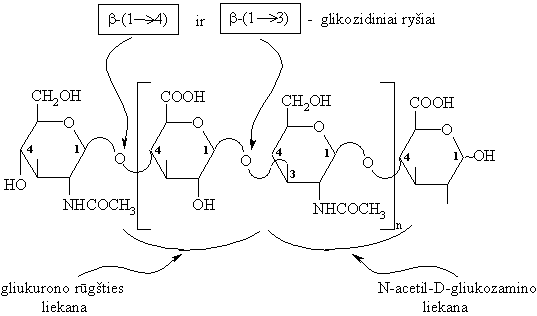
Hialurono rűgštis biosistemose egzistuoja anijonř formoje.
Hialurono rűgšties randama akies stiklakűnyje, bambos virkštelëje, sąnariř skystyje ir kt. Klampűs hialurono rűgšties tirpalai apsaugo jungiamąjá audiná nuo mikrobř prasiskverbimo, sąnarius (amortizatorius).
Chondroitinsulfatai
Jie sudaryti iš disacharidiniř N-acetilintř chondrozino likučiř, sujungtř b-(1 4)-glikozidiniais ryšiais. Chondrozinas sudarytas iš D-gliukurono rűgšties ir D-galaktozamino, susijungusiř b-(1 3)-glikozidiniu ryšiu.
Chondroitinsulfato makromolekulës fragmentas:
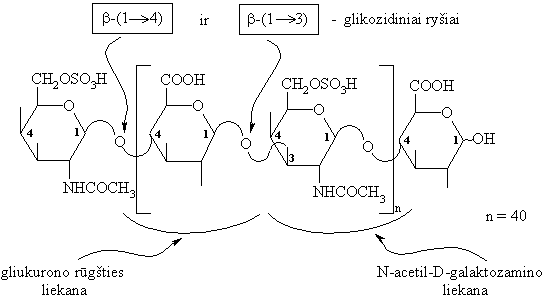
Chondroitilsulfatř (jungiamojo audinio polisacharidř) yra odoje, kremzlëse, sausgyslëse ir kt.
Proteoglikanai
Tai mišrűs biopolimerai,
sudaryti iš polisacharido (iki 95%) ir baltymo. Jř molekulës
susijungę kovalentiniais (O- ir N-glikozidiniais) ir tarpmolekuliniais ryšiais.
Proteoglikanus linkę sudaryti hialurono rűgštis bei chondroitinsulfatai.
Glikoproteinai
Tai mišrűs biopolimerai, sudaryti iš baltymo (iki 90%) ir oligosacharidř (3-25 monosacharido likučiai). Jř molekulës susijungę O- arba N-glikozidiniais ryšiais.
Glikoproteinai – ląsteliř membranř baltymai, apsauginiai baltymai (imunoglobulinai), hormonai, fermentai, plazmos baltymai, nulemiantys kraujo grupę.
Išmokę šá skyriř turite žinoti:
monosacharidř ir jř dariniř struktűrą bei ávairias izomerijos rűšis;
monosacharidř ir jř dariniř rűgštines-bazines, kompleksodarines, elektrofilines-nukleofilines ir oksidacines-redukcines savybes;
gliukozës katabolizmo – glikolizës reakcijas;
disacharidř ir polisacharidř struktűros ir savybiř ypatumus.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 6717
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved