| CATEGORII DOCUMENTE |
| Bulgara | Ceha slovaca | Croata | Engleza | Estona | Finlandeza | Franceza |
| Germana | Italiana | Letona | Lituaniana | Maghiara | Olandeza | Poloneza |
| Sarba | Slovena | Spaniola | Suedeza | Turca | Ucraineana |
DOCUMENTE SIMILARE |
|
NUCLEÓTIDOS Y ÁCIDOS NUCLEICOS
Los nucleótidos son un grupo de macromoléculas que participan en los procesos de transmisión y expresión de la información genética. Existen 2 tipos de ácidos nucleicos, el ác. ribonucleico (RNA o ARN) y el ác. desoxirribonucleico (DNA o ADN).
El nombre de los ácidos nucleicos hace referencia a su característica de ácidos, pues tienen ácido fosfórico, y la primera vez que se localizaron, fue en el núcleo de células eucariotas, pero sabemos hoy que también pueden existir en las mitocondrias, cloroplastos, y citoplasma, y también en todo procariota y virus.
Desde el punto de vista químico, los ácidos nucleicos son polinucleótidos, polímeros de nucleótidos que están unidos por un enlace característico, el fosfo-di-ester, que es un ácido fosfórico que se une por dos enlaces éster sucesivamente a dos grupos alcoholes.
Los nucleótidos están formados por tres componentes, que son el ácido orotfosfórico, una pentosa y una base nitrogenada.
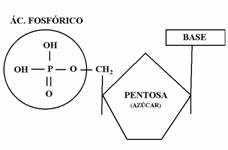
Pentosas
En el caso de los nucloeótidos de ARN (ribonucleótidos), la pentosa es la ribosa (ver temas de azúcares). En el ADN (desoxirribonucleótidos), las pentosas son la 2-desoxirribosa.
Ahora hablaremos de carbonos “prima” (1’, 2’, 3’) para diferenciarlos de los C’s de las bases nitogenadas (1, 2, 3).
Bases nitrogenadas:
Hay dos tipos, las púricas y las pirimidínicas, según de qué anillo deriven, la purina o la pirimidina.
Los sustituyentes en el anillo determinan las bases.
Púricas: adenina y guanina
Pirimidínicas: timina (en ADN solamente),uracilo (solo en ARN) y citosina (en ambos).
Además de estas bases nitrogenadas, que son mayoritarias, existen otras minoritarias que algunas de ellas forman parte de solo unos tipos determinados de ácidos nucleicos, y otras que son productos intermediarios en el metabolismo de éstos.
Existen análogos sintéticos usados en terapia anticancerosa y antivírica.
Las bases nitrogenadas pueden existir en 2 formas que se llaman tautoméricas (ceto y enol). La primera predomina a pH fisiológico. En las “enol”, el doble enlace del oxígeno se transforma en sencillo. Que la base esté en una u otra forma influye en su capacidad para formar puentes de hidrógeno. Esto es de gran relevancia a la hora de estudiar ls estructura y función de los ácidos nucleicos, ya que estas bases nitrogenadas se emparejan entre sí por puentes de hidrógeno. En el caso del ARN esta molécula es monocatenaria pero pueden existir regiones bicatenarias por apareamientos de pares de bases complementarias.
Las bases se unen siempre una púrica con una pirimidínica, unión fundamental porque las primeras son grandes, con dos anillos, y las segundas pequeñas, con un anillo, lo que nos garantiza que el par de bases tiene el tamaño constante.
Se unen siempre G + C por 3 puentes de hidrógeno, una unión un poco más fuerte que la A + T (en ADN) o A + U (en ARN), formada por 2 puentes de hidrógeno.
En estos puentes de hidrógeno participan los grupos ceto. Si las bases están en forma enol, esto influye, y puede dar lugar a la formación de puentes de hidrógeno incorrectos.
Unión entre los constituyentes:
La pentosa y la base nitrogenada se unen por
un enlace N-glucosídico, que se establece entre el C![]() de la pentosa (el anomérico), con el N
de la pentosa (el anomérico), con el N![]() de las pirimidinas, o bien con el
de las pirimidinas, o bien con el ![]() de las purinas. El compuesto resultante es un “nucleósido”.
Entorno a este enlace hay posibilidad de giro, y esto hace que en los ácidos
nucleicos puedan existir diferentes conformaciones según la posiciónes
relativas que ocupan la pentosa y la base nitrogenada.
de las purinas. El compuesto resultante es un “nucleósido”.
Entorno a este enlace hay posibilidad de giro, y esto hace que en los ácidos
nucleicos puedan existir diferentes conformaciones según la posiciónes
relativas que ocupan la pentosa y la base nitrogenada.
De entre estas conformaciones están la SIN y la ANTI, que dependen de cómo esté colocada la base nitrogenada con respecto a la pentosa (base hacia fuera de la pentosa o hacia adentro).
Si a un nucleósido le añadimos un ácido fosfórico mediante un enlace éster, tenemos un nucleótido. El ác. se une por enlace éster a las posiciones 5’ o 3’ (a los OH’s). Aquí, al unirse una única molécula de ácido fosfórico, el resultante es un nucleósido monofosfato.
Denominación: (según la base y la pentosa)
Si el azúcar es la ribosa, y la base nitrogenada la adenina, el nucleósido resultante es la adenosina. El símbolo es la inicial de la base nitrogenada. Si agregamos un ácido fosfórico, obtenemos un nucleótido, ácido adenílico, uridílico, guanílico . Se representan como los anteriores con la misma inicial y con una “p” minúscula a la izquierda, lo que quiere decir que el ácido fosfórico está unido al C 5’ (también se pone 5’-AMP, y normalmete se suele decir AMP cuando es el 5’). Si el ácidos fosfórico se une en la posición 3’, para diferenciarlo, colocamos la “p” en el lado derecho (o 3’-AMP).
Si la pentosa es una desoxipentosa, se añade “desoxi-“ a la palabra (desoxiadenosina, ácido desoxiadenílico).
Hemos mencionado los nucleótidos monofostato. Existen también nucleótidos di y trifosfato, y momofosfato cíclicos.
Si al grupo fosfato en posición 5’ le añadimos otro ácido fosfórico, tenemos un nucleótido difosfato, que lo indicaremos poniendo en la nomenclatura dos “p’s” minúsculas o una D (ADP). Si añadimos otro ácido fosfórico a la molécula obtenemos un nucleótido trifosfato, como el ATP.
Los enlaces entre fosfato y fosfato son “anhídridos”.
Entre los monofosfato cíclicos está el AMP cíclico, en el que la molécula de fosfato se une a los carbonos 5’ y 3’ por dos enlaces éster.
Los nucleótidos mono, di, trifosfato y lo monofosfato cíclicos son importantes porque, además de formar los ácidos nulceicos, intervienen de forma libre en reacciones metabólicas y reacciones de intercambio de energía en la célula.
Los nucleótidos se pueden representar así:
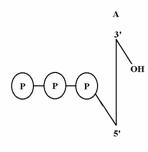
Los nucleótidos se unen por enlaces fosfodiester para formar los polinucleótidos, y se establecen por un enlace éster en el grupo OH 3’ de la ribosa, a al OH 5’ de otra pentosa. En este momento tendríamos un dinucleótido. Aumentando el número de nucleótidos tendríamos trinucleótidos, tetranucleótidosoligonulceótidos, polinucleótidos, ácidos nucleicos.
En los polinucleótidos tenemos un esqueleto
de uniones azúcar-fosfato constante para el ADN y ARN (fosfato – pentosa,
fosfato – pentosa.) siempre en el mismo sentido (5’ Lo característico de la
secuencia es como van situadas las bases de estos nucleótidos (G, A, T, U,
C) desde el nucleótido 5’ fosforilado, hasta el 3’ libre (con OH).
Un ejemplo sería: ![]() CGUA
CGUA![]() . Si solo pusiésemos CGUA sobreentenderíamos lo demás.
. Si solo pusiésemos CGUA sobreentenderíamos lo demás.
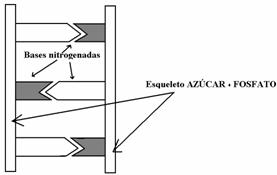
Por apareamiento con puentes de hidrógeno se unen 2 bases complementarias, formando cadenas dobles como el DNA. Las cadenas de DNA son un poco más estables que las de ARN, sobre todo, respecto a la hidrólisis alcalina. Esto se debe a que en el DNA la pentosa es la desoxirribosa, y en esta no pueden formarse anillos por no tener OH sino H .
Que el ADN contenga timina en vez de uracilo también afecta a la estabilidad de este ácido nucleico, porque pueden producirse desaminaciones. Por ejemplo, la desaminación de citosina da uracilo.
Otra diferencia existente entre el ADN y al ARN es que en la composición de los ARN existen unas relaciones de bases nitrogenadas que no encontramos en los ADN’s. En ADN, la concentración de Timina es igual a la de Adenina, y la [G] = [C].
El contenido de G y C varía de una especie a otra pero no en individuos de la misa especie. En el ARN la composición es más variable.
Estructura secundaria (3D) de los polinucleótidos
DNA:
La mayor parte de las cadenas de ADN son bicatenarias, con excepción de algunos virus. Estas dos cadenas discurren antiparalelas:
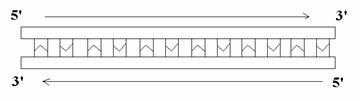
Dentro de ellas se encuentran los pares de bases unidos por puentes de hidrógeno de manera que siempre se juntan una púrica con una pirimidínica. El ancho es de 11
La forma mayoritaria es la de DNA-B o de “wattson y Crick” (1953 premio nobel). En esta, la estructura bicatenaria se enrolla en una doble hélice dextrógira , en la que se puede observar que cada vuelta de hélice se corresponde a 10 pares de bases. Hay una distancia de 3,4 entre cada par de bases, con lo que una vuelta de hélice tiene 34 El ancho de la doble hélice tiene, también constante, 2nm.
Hay dos tipos de surcos, los “surco mayor” y “surco menor”. Aquí, los pares de bases están en el interior y los fosfatos en el exterior (cargados negativamente).
Estas moléculas, en disolución, presentan estructuras largas, flexibles, de aspecto filamentoso, que se vuelve más compacto cuando la molécula reacciona con otros componentes de la célula, como proteínas.
Además de esta forma de ADN-B existen otras formas isoméricas. Las más estudidadas son el ADN A y el Z.
Del ADN-A todavía no se conoce bien su función. Se piensa que el DNA puede pasar de una forma a otra al interactuar con componentes celulares. Se cree que el DNA adopta la forma A cuando está formando híbridos. Con menos del 75% de humedad adopta la forma A.
EL DNA-A presenta 11 nucleótidos por vuelta. Es una hélice más corta y ancha que la del ADN-B. Aquí los pares de bases no aparecen perpendiculares al eje mayor de la hélice como aparecía en la hélice ADN-B. Los surcos mayor y menor están menos diferenciados en tamaño. Es también dextrógira.
El DNA-Z es levógiro, contiene 12 nucleótidos por vuelta. Es una molécula más delgada y solo hay un único y profundo surco. Los grupos fosfato se disponen en zig-zag.
Se ha observado también la formación de triplex, 3 hélices, e icluso cuartetos. Se dan solo en determinadas secuencias, y posibilitan la formación de diferentes puentes de hidrógeno.
El tamaño de las moléculas de ADN es diferente según el organismo del que proceda. Las más pequeñas constan de miles de pares de bases, y las grandes, de millones de pares.
Se suele hablar del tamaño en kilobases (![]() bases).
bases).
Forma
Algunas son lineales, como los cromosomas eucariotas. Otras, con los extremos unidos por enlaces fosfo-di-éster, forman un anillo, como los cromosomas de bacterias, ADN mitocondrial, de ciertos virus, y de los plásmidos (ADN en el citoplasma que se replica autónomamente).
Los ADN circulares se superenrrollan sobre sí mismos, y a estos se les denomina ADN’s superenrrollados
También pueden asociarse como vitaminas para enrollarse más.
La eschierichia coli, pequeña, tiene un cromosoma de lngitud 1mm, y está superenrrollado y asociado a prtoteínas y a ciertos componentes de la membrana plasmática.
El ADN lineal no puede superenrrollarse. En eucariotas se pliega alrededor de proteínas y así es como se empaqueta para formar la cromatina.
La estructura de la cromatina es muy variable, pudiendo encontrarse desde estados poco compactos (fibras de 10 nm de diámetro) hasta estados muy empaquetados (durante la metarase). Entre estos grados, existen intermedios.
La cromatina está formada por las proteínas “histonas”, ficas en aminoácidos básicos (cargadas positivamente) a pH fisiológico lo que les permite interactuar con los grupos fosfato (cargados negativamente) del DNA.
En células de mamíferos existen 5 tipos: ![]() ,
, ![]() ,
, ![]() ,
, ![]() y
y ![]() .
.
Las cuatro últimas están formando un octámero, con dos unidades de cada una. Alrededor de éste, da dos vueltas la fibra del ADN, formando así los nulceosomas, unidad repetitiva de la fibra de cromatina.
El ADN que une los nucleosomas es el
“espaciador”, que se une al siguiente espaciador por una molécula de ![]() .
.
Esta fibra se pliega para dar lugar a una fibra más gruesa, de 30 nm, en la que cada 6 nucleosomas dan una vuelta de hélice (se enrollan). Así se consigue empaquetar la cromatina unas 50 veces.
En la metafase el empaquetamiento es de 100 veces más. Esto se alcanza por interacción con otras proteínas.
El DNA mitocondrial es mucho más sencillo y es circular.
Debido a que las uniones del ADN son puentes de hidrógeno, es posible separar las dos cadenas rompiendo las uniones con un aporte bajo de energía. Esto produce la desnaturalización o fusión del ADN, y puede realizarse por un aumento de pH o de temperatura.
La temperatura a la que se desnaturaliza el
ADN al 50% es la ![]() o temperatura de
fusión, y es una función directa del contenidos de pares citosina-guanina, pues
se enlazan por 3 puentes de hidrógeno. A mayor concentración de
guanina-citosina mayor temperatura.
o temperatura de
fusión, y es una función directa del contenidos de pares citosina-guanina, pues
se enlazan por 3 puentes de hidrógeno. A mayor concentración de
guanina-citosina mayor temperatura.
Este proceso es reversible. Si volvemos a temperatura ambiente, el dúplex vuelve a formarse, denominándose a esto “renaturalización”
La desnaturalización da lugar al “efecto hipercrómico”, que consiste en que cuando se desnaturaliza, aumenta su absorbancia en el ultravioleta.
Lo que absorbe la UV, en realidad, son las bases nitrogenadas, y cuando se separan las hebras, estas quedan expuestas, captando así mejor la UV.
Existen métodos como la absorbancia para determinar ácidos nucleicos, o como la transformación de estos en compuestos coloreados, que pueden ser determinados por colorímetros. Un coloreador es el “orcinol”, específico para ribosa, o la difenilamina, para la desoxirribosa.
El DNA también se puede identificar por fluorescencia con el bromuro de “eidio”, que se intercala entre bases y emite una coloración característica.
Secuencias palindrómicas
Son aquellas secuencias que se pueden leer igual en un sentido u otro. Por ejemplo:
5’-AAATTT-3’ = 3’-TTTAAA-5’
Son regiones con un tipo de simetría binaria importantes porque se forman estructuras secundarias características en los ácidos nucleicos a los cuales se unen proteínasque regulan la expresión genética, pero porque también porque hay encimas de restricción que rompen los enlaces fosfo-di-éster, y algunas reconocen estas secuencias y son dianas de algunas encimas.
![]()
ARN
Estructura de los ácidos ribonucleicos
Son moléculas monocatenarias de tamaño mucho más pequeño. Estas tienen entre 70 y 10.000 nucleótidos.
Aunque la molécula es monocatenaria, pueden existir regiones bicatenarias, que se forman por emparejamiento antiparalelo de bases en la misma cadena, formando lazos, orquillasimportantes para la función de ese ARN.
Las cadenas de ARN pueden unirse con cadenas complementarias de ADN, dando lugar a la hibridación.
El RNA es más abundante que el DNA en la célula, y existen diferentes tipos de ARN’s:
ARNm (o mensajero),
ARNr (ribosómico),
ARNt (transferente),
ARNsn (nuclear pequeño),
ARN heterogéneo nuclear.
El más abundante es el rRNA, y representa el 80% del total de los RNA’s en la célula, seguido del transferente.
Además, se han identificado los denominados “ribozimas”, que son RNA’s que presentan por sí mismos función catalítica.
El ARNm:
Se caracteriza porque no presenta una configuración típica que lo diferencie de los demás, ya que en una célula puede existir al mismo tiempo miles de moléculas de ARNm con una secuencia o estructura diferente.
El mensajero contiene la información que dirige el ensambjaje de los aminoácidos para formar los polipéptidos. Esta información está contenida en tripletes de bases denominados “codones”, específicos para cada aminoácido.
Los ARN mensajeros de procariotas y eucariotas son distintos. El de eucariotas es monogenico, contiene la información para un único polipéptido, mientras que en procariotas, este es poligénico, y puede codificar varios polipéptidos. También hay diferencias en el sistema del proceso de maduración del ARN.
El ARNt:
Los RNA transferentes son los más pequeños, porque contienen entre 70 y 95 nucleótidos. Existen en las célula vivas muchos diferentes (hasta 60).
La función de estos ácidos nucleicos es unir a los aminoácidos en la biosíntesis de proteínas, siguiendo el mensaje que le da el ARNm. Colaboran en el ensamblaje de los aminoácidos correctamente.
Esto lo sigue por el anticodón, un triplete de pares de bases complementaria al codón del ARNm.
Las moléculas del transferente son monocatenarias, pero existen regiones con apareamientos de bases intracatenarios. Esto se manifiesta porque cuando los transferentes se desnaturalizan, muestran efecto hipercrómico.
La estructura bidimensional, secundaria, de los transferentes es la estructura en “trébol”, que consta de un extremo abierto, tres brazos principales, y un brazo menor. A su vez, este trébol se pliega sobre sí, y a esta estructura se la denomina en “L”.
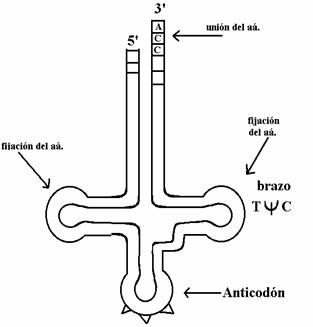
Todos los transferentes tienen en el brazo abierto los extremos 5’ y 3’. Todos acaban con CCA en el 3’, que es donde se une el aá. por enlaces covalentes durante la biosíntesis proteica.
El brazo ![]() C está asociado con la fijación del aminoacil tRNA
(RNA + aá.)
C está asociado con la fijación del aminoacil tRNA
(RNA + aá.)
El asa 2 es donde está el anticodón, y se caracteriza por tener en las posiciones adyacentes nucleótidos modificados.
El asa 3 contiene dihidrouracilo. Interviene también en la fijación al ribosoma del aminoacil tRNA.
Una característica de todos los ARNt es la
presencia de nucleótidos modificados. Hay una gran variedad. Los más comunes
son bases metiladas, nucleótidos con dihirouracilo, y también la
“pseudouridina”, representada por “![]() ”(fi). Aquí lo que varía frente al nucleótido normal es que
la base nitrogenada se une a la ribosa por el
”(fi). Aquí lo que varía frente al nucleótido normal es que
la base nitrogenada se une a la ribosa por el ![]() y no con el 1, lo
normal.
y no con el 1, lo
normal.
ARNr
El ARN ribosomal es el más abundante. Constituye aproximadamente el 65% de los ribosomas.
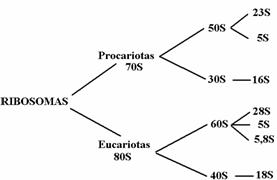
Los ribosomas intactos don diferentes en procariotas y eucariotas.
En procariotas, el tamaño es de 70S y está formado por dos subunidades, una mayor (de 50S) y una menor (de 30S). La unidad mayor o “pesada” contiene 2 tipos de RNA, de 23S y de 5S. La menor o ligera, solo posee un tipo de 16S.
En eucariotas el ibosoma es de 80S. La subunidad mayor, de 60S, contiene tres RNA’s, de 28S, 5S, y 5,8S, mientras que la pequeña, de 40S, solo tiene un tipo, de 18S.
Secuencia y estructura de los ARNr:
Sus secuencias están muy conservadas. Son muy parecidas en todas las especies en la escala evolutiva. También presentan estructuras secundarias determinadas por la formación de apareamientos de bases nitrogenadas intracatenarias.
Toma formas diversas según la secuencia. Estas estructuras secundarias influyen en la asociación con las proteínas.
ARNsn (small nuclear RNA)
Estos, se llaman así por su pequeño tamaño (entre 4S y 8S) y porque aparecen en el núcleo. Están en eucariotas, procariotas y virus.
Muchos de estos ARN existen en estado libre, y se asocian con proteínas para formar partículas de ribonucleoproteínas.
El más abundante es el U-snRNA, donde la “U” proviene de que es rico en uracilo y se ha encontrado en núcleos de células eucariotas. Se han descrito 6 tipos de estos U-snRNA. Además de ser ricos en U, también se caracterizan porque contienen pseudouridina, bases metiladas, y porque contiene un elemento estructural en el extremo 5’ llamado CAP, presente también en RNA mensajeros de células eucariotas. Este casquete 5’ consta de un nucleótido de G en el que la guanina está trimetilada, que se une por un enlace trifosfato 5’-5’ a 1 ó 2 moléculas de azúcar que están metiladas en posición 2’.
La función que se ha propuesto para los U-snRNA es en el proceso de maduración del RNAm en eucariotas. Estos participan en la escisión (eliminación) de los intrones, aquellas regiones que no se traducen) y en el empalme de los exones (aquellas que sí se traducen).
A algunos ARN se les ha encontrado función catalítica, lo que ha llegado a cuñar el término de ribozima, ARN con función encimática.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3027
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved