| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Aspirina imbunatateste homeostazia calciului, scade speciile reacive de oxigen si induce productia de NO in celulele endoteliale umane activate de glucoza crescuta
1. Efectul aspirinei asupra concentratiei de calciu intracelular si producerii NO in celulele endoteliale umane expuse la concentratie mare de glucoza
a. INRODUCERE SI OBIECTIVE
Aspirina este un medicament anti-inflamator nonsteroidal folosit pentru tratarea unor boli inflamatoare umane nesevere. In ultimii ani, diferite studii au descris actiuni independente de prostaglandina ale aspirinei ca blocarea factorului de transcriptie NF-kB [Kopp & Ghosh, 1994], scaderea cu 40% al riscului de aparitie a cancerului de colon [Kune et al., 1988; Thun et al., 1993; Thun et al., 1991], protectia celulelor endoteliale de stresul oxidativ [El Midaoui et al., 2002; Podhaisky et al., 1997] si efecte benefice in diabetul miellitus [Hundal et al., 2002]. Totusi, mecanismul de actiune al aspirinei asupra pacientilor cu diabet de tip 2 nu este studiat.
Disfunctiile endoteliale induse de diabet sunt bine cunoscute Cohen, 1993; Manduteanu et al.,1999; Simionescu et al., 1996]. Desi mecanismele exacte prin care concentratiile crescute de glucoza induc disfunctia endoteliului nu sunt cunoscute, s-a aratat ca in conditii de glucoza crescuta este afectata productia endoteliala a diferitilor mediatori (incluzand oxidul nitric). Astfel, in hiperglicemie endoteliul este afectat prin reducerea rezistentei periferice si a cresterii fluxului de sange, ducand mai intai la o crestere urmata de o scadere a activitatii endoteliale si la un risc crescut de ateroscleroza. De asemenea, atat in diabetul experimental cat si diabetul miellitus uman, relaxarea dependenta de endoteliu este diminuata [Costache et al., 2000; Diederich, et al., 1994]. Natura acestui eveniment nu este foarte bine inteleasa, insa un rol important il are homeostazia calciului si scaderea bioactivitatii oxidului nitric (NO), datorita unei cresteri a speciilor reactive de oxigen (ROS).
In disfunctia celulei endoteliale (CE) are loc o crestere a concentratiei intracelulare de calciu [Ca]2+ care poate produce intensificarea permeabilitatii, secretia citokinelor inflamatorii, inducerea transcrierii unor gene si reglarea secretiei de vasorelaxanti. S-a dovedit anterior ca CE expuse unor conditii de glucoza crescuta prezinta o concentratia intracelulara de calciu ([Ca]2+) mai mare datorita eliberarii de anioni superoxizi; mai mult, antioxidantii previn aceste alterari ale [Ca]2+ endotelial. Sunt cunoscute efectele benefice ale aspirinei in reducerea evenimentelor vasculare datorate inhibarii activitatii ciclooxigenazelor cat si a efectului antiinflamator ale carui mecanisme sunt mai putin cunoscute. In acest studiu, am urmarit efectul aspirinei asupra mobilizarii [Ca]2+ in CE umane expuse unor concentratii mari de glucoza cat si a eliberarii de oxid nitric (NO), stiind ca modificarile ale [Ca]2+ influenteaza producerea de NO.
Celule
EA 926 este o linie de celule endoteliale vasculare umana permanenta obtinuta prin hibridizarea HUVEC cu A549 (celule ce provin de la o carcinoma de plaman, umane). Aceste celule au fost obtinute prin amabilitatea Dr. Cora Edgell (Universitatea North Carolina) (Edgell CJ et al., 1983). Sunt crescute in DMEM continand 15% ser fetal de vitel.
Celulele au fost subcultivate in placi de cultura cu 6 godeuri.
Tratatrea celulelor
Celulele au fost cultivate in conditii normale de glucoza (5mM) si in conditii de glucoza crescuta (33mM glucoza). La confluenta, celulele aflate in 33mM glucoza au fost impartite in 2 loturi experimentale: un lot a fost incubat in continuare in 33mM glucoza iar celalalt lot a fost incubat in 33mM glucoza plus 1mM aspirina, 48 ore.
Masurarea Ca2+
Concentratia de calciu intracelulara ([Ca2+]i) a fost determinata folosin fura-2AM. Pe scurt, celulele aflate in diferite conditii experimentale au fost recoltate cu tripsina (0.06%), 2 minute, spalate si incubate in tampon (HBSS) fara calciu continand fura-2AM. Dupa incarcarea cu fura-2AM, 1 ora, la 37 C si l-a intuneric, celulele au fost spalate de doua ori si resuspendate in HBSS. Masuratorile de fluorescenta au fost facute cu ajutorul unui spectofotometru controlat termic si prevazut cu un program asistat de calculator (Shimadzu RF-5001 PC). Lungimile de unda de excitatie au fost setate 340 si 380 nm, iar emisia la 510nm. Eliberarea calciului din stocurile intracelulare a fost urmarita prin stimularea cu histamina (25mM). Pentru a urmari raspunsul datorat intrarii calciului din mediul extracelular, s-a adaugat 2,5mM calciu, celulelor aflate in cuva in tampon fara calciu. Concentratia intracelulara de calciu a fost calculata conform ecuatiei:
[Ca2+]i
(nM) = Kd ![]() ,
,
unde R este raportul fluorescentei la 340 si 380nm. Rmax este raportul de fluorescenta obtinut la adaugarea de 10% Triton si Rmin este raportul obtinut prin adaugarea ulterioara de 5mM EGTA. Kd este constanta de disociere a fura legat de calciu, de 224mM.
Dozarea spectrofotometrica a NOx (NO2- / NO3- ) in mediu de cultura
Am evaluat productia de NO in mediul de cultura al celulelor aflate in coditiile experimentale precizate mai sus, prin masurarea nivelelor de nitriti (NO2-), produsul oxidat al NO, prin reactia Griess. Principiu: reducerea enzimatica a nitratilor (NO3-) din proba, la nitriti (NO2-) si detectia nitritilor cu reactiv Griess (amestec 1:1 de 1% sulfanilamida / 1% N-naftiletilendiamina); se formeaza compusi azo colorati roz-violaceu care se citesc la 540 nm.
Protocolul experimental:
-la 100 ml mediu de cultura / blanc (tampon fosfat T.F 0.1M) / standard (se fac 3-4 standarde: 7.5, 15, 30, 60 mM NaNO3 dintr-un stoc de 120 mM NaNO3 in apa distilata), s-a adaugat:
ml NADPH 2mM
ml FAD 1 mM
ml nitrat reductaza 1,25 U/ ml din stoc 5 U/ 1ml 0.1 M TF
![]() ml tampon fosfat 0.1 M, pH 7.2 - 7.4
ml tampon fosfat 0.1 M, pH 7.2 - 7.4
A urmat incubarea 2 h, la temperatura camerei,la intuneric, apoi peste toate probele s-a adaugat:
ml 1% sulfanilamida, agitare (reactie de diazotare), apoi
ml 1% N-naftiletilendiamina (reactie de cuplare)
si s-au microfugat 5 min, iar supernatantele limpezi au fost citite la 540 nm fata de blanc.
Cantitatea de NO a fost normalizata la numarul total de celule ale fiecarei probe.
c. REZULTATE
1) Efectul concentratiei mari de glucoza asupra calciului intracelular [Ca2+]i in celulele endoteliale (CE) umane.
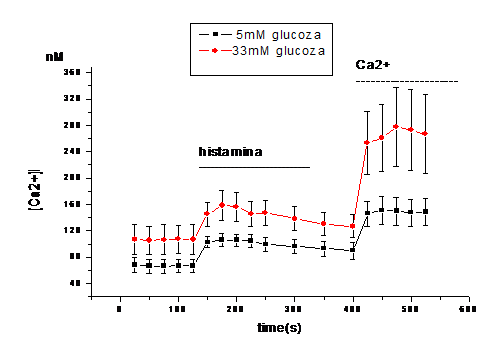
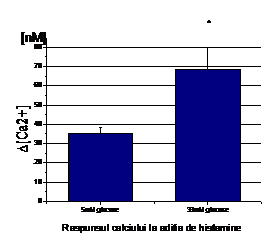
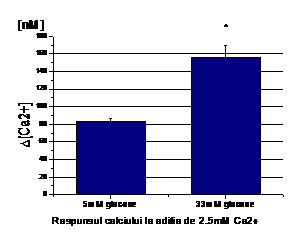
Expunerea celulelor endoteliale la 33mM D-glucoza timp de 6 zile duce la cresterea calciului liber citosolic comparativ cu celulele crescute in conditii normale de glucoza (5mM). Astfel, in absenta Ca2+ extracelular, concentatia bazala de calciu in celulele tratate cu 33mM glucoza este de 106.75 nM in timp ce in celulele control este de 66.88 nM, valori ce sunt stabile in timp (Figura 1, A).
Aditia histaminei (25 mM) induce o crestere tranzienta a [Ca2+]i in ambele conditii (5 si 33mM glucoza), care in timp, scade la o valoare de platou fara sa atinga insa nivelul bazal (Figura 1). Crestera maxima a [Ca2+]i ca raspuns la histamina este mai mare in celulele expuse la 33mM glucoza fata de control (~163.5 vs. ~99.75nM, Figura 1,B). Dupa cum se observa in figura 1, aditia ulterioara de calciu extracelular duce la o crestere a calciului care intra in celule, care este mai mare in celulele crescute in 33mM glucoza (~266.45 nM) decat in celulele control (~148.5 nM, Figura 1,C ).
Figura 1. Concentratia intracelulara de calciu in CE umane activate de glucoza crescuta
 a
a
B C
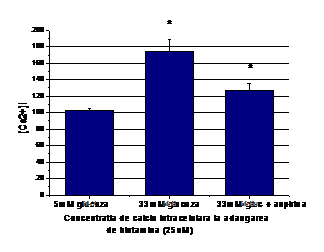
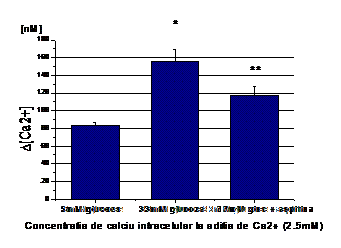
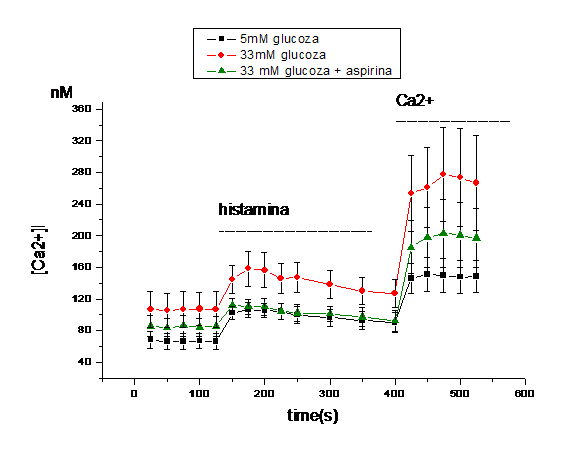
2) Efectul aspirinei asupra homeostaziei calciului celular indusa de concentratia mare de glucoza
Dat fiind faptul ca rezultatele obtinute de noi arata ca nivelele crescute de glucoza duc la o acumulare de calciu intracelular datorata atat calciului care iese din stocurile intracelulare (la adaugarea de histamina), cat si cat si calciului care intra din mediul extracelular, ne-am intrebat daca aspirina are efect aspra acestor procese. Rezultatele experimentelor arata ca incubarea celulelor crescute in 33mM glucoza timp de 48 ore cu 1mM aspirina reduce efectul glucozei asupra [Ca2+]i. Dupa cum se observa in figura 2 A, [Ca2+]i din celulele endoteliale crescute in 33mM glucoza si tratate cu aspirina este redus semnificativ fata de celulele crescute in glucoza 33mM in absenta de aspirina (de la 163 la 104 nM, Figura 2, B).
In plus, aspirina a influentat si calciul care intra din mediul extracelular, scazand acumularea de [Ca2+]i de la ~266 nM in CE crescute in 33mM glucoza, la ~197 nM pentru celulele crescute in 33mM glucoza dar tratate cu aspirina (Figura 2, C).
Figura 2. Efectul aspirinei asupra concentratia intracelulara de calciu in CE umane activate de glucoza crescuta
A
|
B
|
C
|
3) Efectul aspirinei asupra productiei de oxid nitric in celulele endoteliale expuse la concentratie mare de glucoza
Sinteza oxidului nitric (NO) endotelial depinde de calciul intracelular. Exista multe studii care arata ca hiperglicemia afecteaza productia de NO in vasculatura. In studiul nostru, am investigat o posibila relatie intre efectul aspirinei asupra mobilizarii calciului si productia de NO in CE crescute in mediu cu concentratie crescuta glucoza. In acest scop, supernatantul celulelor ce urmau sa fie supuse determinarii calciului intracelular a fost colectat si s-a determinat concentratia de nitriti eliberata de fiecare proba.
Figura 3. Eliberarea oxidului nitric (NO) in mediu de cultura al CE
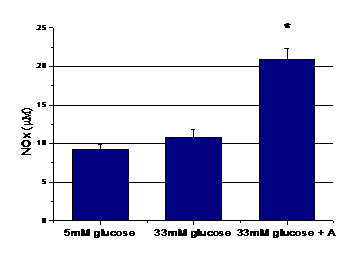
Dupa cum se observa in figura 3, productia de NO elibearata de CE crescute in 33mM glucoza este usor crescuta, fata de control (5mM glucoza), dar nu semnificativ. Tratarea celulelor crescute in 33mM glucoza cu aspirina pt 48 ore, induce o crestere semnificativa (cu 108%) a productiei de NO fata de celulele aflate numai in glucoza (Figura 3)
4)Viabilitatea celulara
Pentru a verifica daca rezultatele obtinute de noi sunt fiabile si nu datorate unui eventual efect citotoxic al aspirinei, am performat experimente de viabilitate a celulelor crescute in conditii normale de glucoza, activate cu glucoza crescuta si tratate cu aspirina. In conditiile experimentale testate, viabilitatea celulelor aflate in diferitele conditii de incubare a fost de peste 90% (figura 4), intre probe neexistand diferente semnificative din punct de vedere statistic
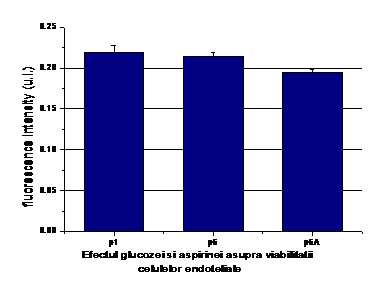
Figura 4. Viabilitatea CE aflate in
diferite conditii experimentale
d. DISCUTII SI CONCLUZII
Concentratia de calciu intracelulara este un reglator cheie al functiei celulare, modificarile homeostaziei calciului afectand functiile vasculare ca permeabilitatea, secretia unor citokine inflamatoare, expresia genica si proteica a diferitelor molecule [Hempel et al., 1997; Conboy et al., 1999]. Efectul glucozei crescute asupra raspunsului Ca2+ la diferiti agonisti este dependent de perioda de incubare cu glucoza, specia celulara si agonist [Pieper & Dondlinger, 1996]. Exista studii care au aratat ca atat expunerea la timp scurt cat si expunerea prelungita la glucoza crescuta modifica semnalizarea Ca2+. Astfel, incubarea celulelor endoteliale cerebrale bovine timp de 2 ore cu 23mM glucoza, nu modifica nivelele bazale de calciu (calciul liber citosolic), insa la adaugarea histaminei, calciul nu se modifica, celulele fiin incapabile sa-si elibereze calciul din stocurile intracelulare. De asemenea, la adaugarea calciului expracelular celulelor expuse la glucoza crescuta, calciul extracelular nu intra in celula, concentratia intracelulara de calciu ramanand nemodificata [Kimura et al.,1997]. In contrast, incubarea celulelelor aortice porcine cu glucoza crescuta (44mM) timp de 24 ore creste atat concentratia bazala de calciu, cat si semnalizarea calciului ca raspuns la agonisti (histamina, calciu extracelular), printr-un mecanism necunoscut [Graier et al., 1996; Wascher et al., 1994].
S-a aratat ca antioxidantii (vitamina E, probucolul, GSH si vitamina C) diminueaza raspunsul [Ca2+]i, crescut de concentratia mare de glucoza, prin reducerea nivelului anionilor superoxizi de oxigen [Graier et al., 1997]. De asemenea, au inceput sa apara dovezi care sugereaza ca aspirina are proprietati antioxidante [El Midaoui et al., 2002; Ghiselli et al., 1992; Podhaisky et al., 1997] si efecte favorabile in diabet [Hundal et al., 2002], insa mecanismele de actiune nu au fost inca elucidate. Astfel, in studiul nostru, am urmarit atat efectul aspirinei asupra mobilizarii calciului in CE activate de glucoza crescuta.
Rezultatele noastre demonstreaza ca atat eliberarea calciului intracelular din stocurile intracelulare (ca raspuns la histamina) cat si intrarea calciului extracelular in celula, sunt modificate in urma expunerii celulelor endoteliale la concentratie crescuta de glucoza. Tratarea cu aspirina a celulelor activate de glucoza produce restabilirea semnalizarii calciului.
Este foarte bine cunoscut ca sinteza oxidului nitric endotelial depinde de calciul intracelular si ca hiperglicemia afecteaza productia de NO in vasculatura. Si pentru ca aspirina a imbunatatit semnalizarea calciului, am urmarit daca productia de NO eliberata in mediul de cultura in conditii de glucoza crescuta, este modificata de aspirina. Astfel, in studiul nostru am obtinut ca productia de NO este crescuta smnificativ de aspirina.
Concluzia care se desprinde din prima parte a acestui capitol este ca aspirina, in plus la efectele cunoscute, imbunatateste atat raspunsul calciului cat si productia de NO, in celulele endoteliale umane expuse la concentratie mare de glucoza. Aceste rezultate sugereaza ca aspirina poate diminua disfunctia endoteliala indusa in conditii hiperglicemice.
2. Mecanismul de actiune al aspirinei implicat in imbunatatirea semnalizarii calciului in celulele endoteliale aflate in conditii patologice de glucoza. Evidentierea unei posibile proprietai de antioxidant al aspirinei.
a. INTRODUCERE SI OBIECTIVE
Aspirina este un medicament antiinflamator larg raspandit al carui mecanism de actiune acceptat este ca inhiba sinteza de prostaglandine, prin blocarea ciclooxigenazelor (COX-1 si COX-2). Asa cum am amintit mai sus, in ultima vreme, mai multe studii au descris actiuni independente de prostaglandine ale aspirinei, printre care efecte benefice in diabetul miellitus [Hundal et al., 2002]. Astfel, s-a demonstrat ca tratamentul cu aspirina previne dezvoltarea hipertensiunii si reduce semnificativ rezistenta la insulina in sobolanii hraniti cu concentratii mari de glucoza. S-a sugerat ca aspirina produce aceste efecte prin proprietatile sale antioxidante, gasindu-se ca aceasta previne cresterea productiei de anioni superoxizi (O2-) in aortele acestor sobolani. Totusi, mecanismul de actiune al aspirinei la pacientii cu diabet de tip 2 nu este clar.
Datele obtinute in prima pare a acestui proiect, folosind un sistem in vitro de celule endoteliale (EA926) care au fost crescute in conditii normale si patologice de glucoza arata ca homeostazia calciului este afectata de concentratii mari de glucoza. Insa, tratarea celulelor crescute in concentratii mari de glucoza timp de 48 ore cu aspirina (1mM) amelioreaza efectele glucozei reducand concentratia de calciu intracelulara ([Ca2+]i) catre valorile obtinute la celulele crescute in conditii normale de glucoza.
Se stie ca in diabet, excesul de glucoza duce la o crestere a productiei de specii reactive de oxigen (ROS) prin cateva cai, printre care autooxidarea glucozei, activarea NADPH oxidazei, glicarea proteinelor neenzimatice. Acumularea de anioni superoxizi este un eveniment ce candideaza la patogeneza vasculara si daunele endoteliale in diabet, si se aduc din ce in ce mai multe evidente pentru rolul important pe care il au ROS in semnalizarea celulara [Lyle AN, Griendling KK., 2006; Zmijewski et al., 2005; Tao et al., 2005]. Astfel, cantitati crescute de O2- inactiveaza NO producad peroxinitriti si produce o crestere a concentratiei de calciu intracelular ca raspuns la histamina.
In studiul nostru, pentru a evidentia mecanismul de actiune al aspirinei de imbunatatire a raspunsului calciului am urmarit efectul acesteia asupra productiei de specii reactive de oxigen in conditii de glucoza crescuta, analizand astfel proprietatea de atioxidant a aspirinei. Dat fiind faptul ca NADPH oxidaza este una din sursele cele mai importante de producere a ROS si ca activarea ei este dependenta de protein kinaza C am testat activitatea NADPH oxidazei si expresia PKC in conditii de glucoza crescuta. De asemenea am urmarit efectul aspirineie asupra acestor doua molecule implicate in productia de specii reactive de oxigen.
b. MATERIALE SI METODE
Celule
Celulele EA 926 - linie de celule endoteliale umane permanenta, au fost crescute in DMEM suplimentat cu 10% ser fetal de vitel si 100U/ml penicilina, 100mg/ml streptomicina intr-un incubator la 37 C, 5% CO2 si 95% aer.
Tratatrea celulelor
Celulele au fost cultivate in conditii normale de glucoza (5mM) si in conditii de glucoza crescuta (33mM glucoza). La confluenta, celulele aflate in33mM glucoza au fost impartite in 2 loturi experimentale: un lot a fost incubat in continuare in 33mM glucoza iar celalalt lot a fost incubat in 33mM glucoza plus 1mM aspirina, 48 ore.
Determinarea speciilor reactive de oxigen
Speciile reactive de oxigen din celule au fost determinate folosind forma diacetata a fluoroforului DCFH (DCFH-DA) care este preluat de celule si clivat la forma DCFH de catre esterazele intracelulare. In prezenta de H2O2, DCFH oste oxidat la DCF. Fluorescenta emisa a fost masurata la spectofluorimetru, folosind lemisie=527 nm si lexcit=516 nm. Dupa masuratorile de fluorescenta, celulele au fost recuperate si lizate pentru a se determina concentratia proteica totala prin metoda BCA (Sigma). Cantitatea de specii reactive de oxigen a fost exprimata ca unitate de fluorescenta per miligram proteina.
Masurarea anionilor superoxizi (O2-) eliberati in mediul de culura de celule
Anionii superoxizi au fost masurati cu ajutorul reactiei de reducere a ferricitocromului c. Celulele aflate in diferitele conditii au fost incubate cu ferricitocrom c (16mM) in prezenta sau absenta superoxid dismutazei (SOD, 400U/ml). Reactia rezultata a fost masurata la spectofotometru la 550nm dupa 30 minute. Diferentele de absorbtie in absenta si prezenta SOD au aratat direct extintia datorata O2-. Concentratia de O2- a fost calculata folosind coeficientul de extintie molar al formei reduse a ferricitocromului c, e
Testarea activitatii NADPH oxidazei
Producerea O2- dependenta de NADPH oxidaza a fost determinata din omogenate obtinute prin lizarea celulelor cu un tampon de liza ce contine inhibitori de proteaze. Fericitocromul c si NADPH au fost adaugate in prezenta sau absenta SOD (200 U/ml) si incubate la temperatura camerei timp de 30 minute. Reducerea fercitocromului c a fost evaluata prin citirea absorbantei la 550nm la un cititor ELISA. Rezultatele obtinute au fost exprimate in nanomoli O2- per miligram proteina.
Determinarea expresiei proteice a PKC
Prepararea lizatului celular si fractiilor subcelulare: Celulele au fost spalate de trei ori cu un tampon izotonic (pH:7.2), luate de pe placa intr-un tampon de omogenizare ce contine inhibitori de proteaze si sparte cu ajutorul unui omogenizator Dounce (pestle tip B). Lizatul obtinut a fost centrifugat la 100 000x g timp de 1 ora. Supernatantul a fost colectat ca fractia citosolica. Peletul a fost resuspendat in tampon omogenizator cu 1% Triton X-100, si centrifugat din nou la 100 000x g, 1 ora. Supernatantul a fost colectat ca fractie membranara.
Western blot: Ambele fractii colectate de la diferitele probe au fost supuse unei electroforeze SDS-PAGE.
Imunoblot: Dupa stingerea legarilor nespecifice realizata prin incubarea membranelor de nitroceluloza cu 5% lapte degresat in TBS continand 0,1% (TBS-T) Tween 20, acestea au fost ulterior prelucrate prin incubarea peste noapte, la 4C, cu anticorpul monoclonal anti- PKC, iar ulterior cu anticorpul secundar (dilutie 1: 5.000) conjugat cu peroxidaza din hrean (HRP, horseradish peroxidase). Spalarile dintre incubari s-au realizat cu TBS-T. Complexele Ag-Ac au fost vizualizate prin chemiluminiscenta, folosind un kit de ECL (bloturile au fost incubate cu substratul enzimatic chemiluminiscent Pierce, iar luminiscenta obtinuta a impresionat un film fotografic).
c. REZULTATE
1) Efectul aspirinei asupra speciilor reactive de oxigen (ROS) intracelulare induse de glucoza crescuta
Dat fiind ca se stie ca nivelele crescute de glucoza duc la cresterea speciilor reactive de oxigen (ROS) care la randul lor produc defecte in functionarea celulei, am urmarit efectul nivelelor crescute de glucoza asupra ROS, in sistemul nostru celular. De asemenea, se stie ca ROS interactioneaza cu NO si produce peroxinitriti, scazand biodisponibilitatea NO. Cum in experimentele noastre anterioare am aratat ca aspirina duce la cresterea NO in conditii de glucoza crescuta, am testat efectul aspirinei asupra ROS in conditii de glucoza crescuta.
Rezultatele experimentelor arata ca incubarea celulelor crescute in 33mM glucoza cu 1mM aspirina timp de 48 ore reduce speciile reactive de oxigen induse de concentratia crescuta de glucoza.
Dupa cum se observa in figura 5, concentratia crescuta de glucoza creste semnificativ ROS (p=0.0005), iar aspirina aduce la nivel bazal speciile reactive de oxigen produse de glucoza (p=0.0007).
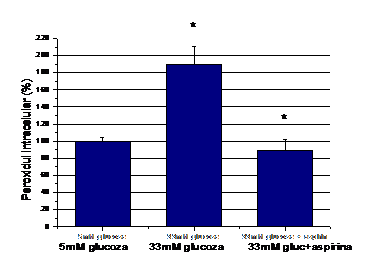
Figura 5.
Nivelul speciilor reactive de oxigen intracelulare
Efectul aspirinei asupra anionilor superoxizi (O2-)i eliberati in mediul de cultura al celulelor expuse la glucoza crescuta
De asemenea am masurat cantitatea de anioni superoxizi (O2-) eliberata de CE aflate in diferite conditii experimentale, prin masurarea reducerii fericitocromului c de catre O2-. Dupa cum se observa in figura 6 cantitatea de O2- este crescuta in celulele aflate in 33mm glucoza fata de control (5mM glucoza, p=0.004). Tratarea cu aspirina a celulelor aflate in conditii de glucoza crescuta inhiba producerea de O2- indusa de glucoza (p=0.008).
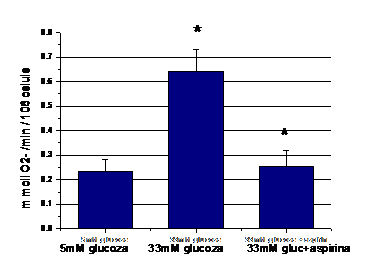
Figura 6. Efectul aspirinei asupra
nivelului O2- eliberati in mediul de cultura
3) Concentratia mare de glucoza activeaza NADPH oxidaza. Efectul aspirinei asupra activarii NADPH oxidazei
Pentru a investigata mecanismule de actiune ale aspirinei de a reduce producerea de O2- in CE expuse la 33mM glucoza, am testat efectul aspirinei asupra activitatii NADPH oxidazei. Enzima NADPH oxidaza a fost aleasa pentru ca studiile anterioare au desemnat-o ca fiind sursa principala de specii reactive de oxigen in vasculatura. Rezultatele obtinute arata ca nivelul crescut de glucoza declanseaza activarea NADPH oxidazei, iar aspirina reduce aceasta activare a NADPH oxidazei, aproape de valorile din probele control (figura 7).
Figura 7. Aspirina scade activitatea NADPH oxidazei indusa de glucoza
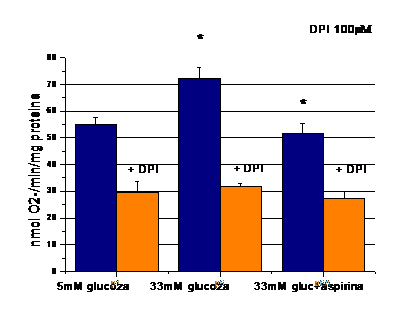
De asemenea, pentru a demonstra ca aspirina actioneaza reduce O2- produsi de NADPH oxidaza si nu de alte surse de anioni superoxizi, am testat efectul aspirinei asupra produceri de O2- in aceleasi probe dar care au fost incubate cu inhibitor specific de NADPH oxidaza, difeniliodonium (figura 7, benzile portocalii). Dupa cum se observa, in cazul in care anionii superoxizi produsi de NADPH oxidaza sunt inhibati, aspirina nu mai are efect asupra O2- produsi de alte surse (existenti in celule, dar produsi de alte surse celulare). Astfel, efectele aspirinei observate asupra ROS se datoreaza actiunii acesteia asupra NADPH oxidazei.
4) Efectul aspirinei asupra activarii PKC indusa de concentratia mare de glucoza
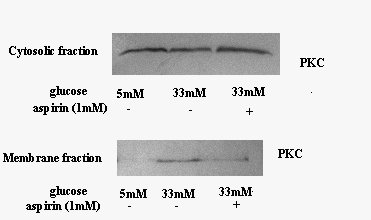
Se stie din literatura ca protein kinaza C (PKC) este activata in diabet, si ca PKC activata duce la producerea de anioni superoxizi prin activarea NADPH oxidazei. Pentru a demonstra ca aspirina scade activarea NADPH oxidazei actionand asupra PKC, am urmarit efectul aspirinei asupra acestei proteine, in conditii de glucoza crescuta prin Western Blot. Rezultatele arata ca in conditii normale, PKC este in majoritate localizata in fractia citosolica (figura 8). Concentratia mare de glucoza induce translocarea PKC din citosol in membrana ceea ce este echivalent cu activarea PKC. Tratarea celulelor expuse la glucoza crescuta cu aspirina reduce translocarea PKC in fractia membranara, demonstrand ca aspirina inhiba activarea PKC.
Figura 8. Aspirina inhiba activarea PKC indusa de 33mM glucoza
d. DISCUTII SI CONCLUZII
Nivelele crescute de glucoza sunt asociate cu un nivel crescut al stresului oxidativ, inlustrat in producere crescuta a speciilor reactive de oxigen. Disfuctiile celulei in conditii hiperglicemice sunt datorate in principal nivelelor crescute ale speciilor reactive de oxigen. Pentru a afla mecanismele implicate in efectele favorabile ale aspirinei (de a imbunatati homeostazia calciului si a creste productia de NO) am urmarit efectul acesteia asupra speciilor reactive de oxigen. Expunerea cronica a celulelor endoteliale la glucoza crescuta induce cresterea dramatica a nivelului speciilor reactive de oxigen, determinate cu ajutorul fluoroforului DCFH-DA. Nivelul anionii supeoxizi de oxigen eliberati in mediul de cultura de CE activate de glucoza este mult crescut, comparativ cu cel din celulele control. Aspirina reduce stresul oxidativ indus de glucoza la nivelul celulelor control (crescute in conditii normale de glucoza).
Aceste rezultate sunt sustinute de experimente anterioare care demonstreaza ca efectele benefice ale aspirinei in prevenirea dezvoltarii hipertensiunii la sobolanii hraniti cu glucoza se datoreaza proprietatilor antioxidante ale acesteia, reducand productia de anioni superoxizi [El Midaoui et al., 2002].
Acumularea de anioni superoxizi este unul dintre candidatii importanti pentru patogeneza vasculara si/sau lezarile endoteliale in conditii hiperglicemice. Recent, s-a aratat ca speciile reactive de oxigen derivate de la NADPH oxidaza sunt sursa majora de ROS in vasculatura si ca CE si CMN produc ROS prin activarea NADPH oxidazei [Bayraktutan et al., 1998; Mohazzab et al., 1999; Rajagopalan et al., 1996]. De asemenea, este bine cunoscut ca activarea NADPH oxidazei se datoreaza cel putin partial activarii PKC [Hempel et al., 1997], care este considerat eveniment cheie in generarea ROS [Inoguchi et al., 2000]. Astfel, incubarea in vitro a tesutului aortic de la animalele diabetice cu un inhibitor specific al PKC duce la scaderea semnificativa a productiei de superoxid [Hink et al., 2001]. Toate aceste date ne-a determinat sa urmarim efectul aspirinei asupra activarii NADPH oxidazei si a PKC pentru a determina mecanismul prin care aspirina isi exercita proprietatile antioxidante. Rezultatele obtinute arata ca aspirina inhiba activitatea NADPH oxidazei in CE activate de glucoza. De asemenea, aspirina inhiba activarea PKC indusa de glucoza, reducand procesul de translocare al PKC din citoplasma in nucleu. Astfel, putem presupune ca in celulele endoteliale umane activate de concentratia crescuta de glucoza, aspirina restabileste homeostazia calciului prin inhibarea activarii NADPH oxidazei dependenta de PKC, reducand astfel producerea de anioni superoxizi de oxigen.
Speram ca noile proprietati ale aspirinei, de antioxidant, combinate cu cele deja cunoscute (antiinflamator, antiagregant) sa faca din aspirina un medicament important pentru protectia celulelor endoteliale de efectele negative ale nivelelor crescute de glucoza care sunt asociate cu diabetul.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1307
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved