| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
IMUNITATEA DOBANDITA (INDUCTIBILA)
Imunitatea dobandita (inductibila) este starea de rezistenta a organismului fata de factorii 'non-self' antigenici, realizata prin structuri si functii de neoformatie cu un inalt grad de specificitate pentru antigen.
Principalele caracteristici ale imunitatii dobandite sunt urmatoarele:
are un caracter individual, sfera ei de cuprindere referindu-se numai la unii indivizi dintr-o specie;
prima treapta filogenetica pe care apare este reprezentata de vertebrate;
momentul aparitiei este dependent de patrunderea in organism a Atg. si poate fi oricand in cursul dezvoltarii ontogenetice a individului matur imunologic;
are un caracter netransmisibil ereditar numai in unele cazuri se poate transmite vertical de la mama la fat;
efectorii imunitatii dobandite sunt constituiti din structuri si functii de neoformatie de natura umorala (Atc. sau imunoglobuline) si celulara (limfocitele sensibilizate specific si alte tipuri de celule);
actiunea efectorilor este specifica si diferentiata in functie de Atg;
in determinismul imunitatii dobandite sunt implicate mecanisme cu memorie.
Imunitatea dobandita se poate clasifica dupa mai multe criterii:
1. Prezenta antigenului si participarea organismului la elaborarea efectorilor imuni, criteriu dupa care imunitatea dobandita poate fi impartita in:
a) imunitate activa, in care antigenul patrunde in organism pe cai naturale, de regula printr-o infectie naturala, sau pe cai artificiale, in mod voit prin vaccinare; imunitatea activa se instaleaza in 6 - 10 zile, timp necesar producerii efectorilor imuni si este de lunga durata, in functie de capacitatea imunogena a materialului imunizant, poate dura de la cateva saptamani, luni, ani si chiar toata viata.
b) imunitatea pasiva, in care antigenul nu vine in contact cu organismul, care nu-si produce singur efectorii imuni, primindu-i insa prin
transfer de la un organism donator cu o stare de imunitate activa. Prin analogie cu imunitatea activa, imunitatea pasiva poate fi dobandita pe cai naturale, prin transferul efectorilor, de cele mai multe ori a celor umorali de la mama la fat, transplacentar la speciile cu placenta hemocoriala si colostral, la speciile cu alte tipuri de placenta nepermeabila sau pe cai artificiale prin serumizare sau transfer de celule. Imunitatea pasiva se instaleaza chiar in momentul transferului efectorilor imuni si este de scurta durata de la cateva zile la doua - trei saptamani.
2. Tipul predominant de efectori imuni, criteriu dupa care pot fi deosebite urmatoarele tipuri de imunitate:
a) imunitatea de tip umoral in care responsabilitatea starii de rezistenta revine anticorpilor (imunoglobulinelor), care predomina si au capacitatea de a anihila efectul nociv al antigenului;
b) imunitatea de tip celular in care starea de rezistenta se datoreste diferitilor efectori celulari.
Sunt relativ putine antigenele care determina imunitate de tip exclusiv umoral sau celular, in majoritatea cazurilor, cele doua categorii de efectori imuni coexista si actioneaza cooperant in realizarea starii de imunitate.
3. Persistenta antigenului viu ca factor conditionant al starii de imunitate. Dupa acest criteriu, imunitatea se poate clasifica in:
a) imunitate sterila in care starea de imunitate nu este dependenta de persistenta antigenului viu in organsim;
b) imunitatea nesterila, numita si premunitie sau stare premunitiva, care implica necesitatea prezentei antigenului viu in organsim.
Mecanismele prin care organismul recunoaste antigenul, organele in care se produc efectorii imuni si in care acestia pot reactiona cu antigenul, sunt tratate pe larg in capitolele urmatoare.
IMUNITATEA MATERNO - FETALA
Maturarea sistemului imunitar la fetus si nou-nascut este un fenomen complex. El se desfasoara intr-un context particular si trebuie sa tina seama de numeroase aspecte, uneori contradictorii, si de interactiuni variate:
constructia unui sistem imunitar eficace, care sa permita organismului in mod progresiv o defensiva autohtona, contra agresiunilor ulterioare;
schimburile imunologice intre mama si progenitura sa;
la mamifere toleranta fetusului de catre mama in cursul gestatiei;
achizitionarea tolerantei tanarului organism in ceea ce priveste antigenele proprii.
Achizitionarea competentei imunitare se face progresiv pentru toate speciile de animale.
Ca o regula generala, la mamifere, speciile care au o scurta perioada de gestatie prezinta la nastere un sistem imunitar mai matur fata de cel al speciilor cu durata lunga de gestatie.
Celulele suse ale sistemului hematopoietic apar la nivelul sacului vitelin in momentul debutului gestatiei. Ele produc celule din toate liniile hematopoietice si in particular limfocite; aceastea migreaza dupa aceea din sacul vitelin catre ficatul fetal, apoi catre maduva osoasa, putin inainte de nastere. Din maduva osoasa isi asuma rolul pe toata durata vietii animalului.
Timusul este primul organ limfoid primar care se diferentiaza, acesta este responsabil de maturarea limfocitelor T la toate speciile. La pasari bursa lui Fabricius este in legatura cu diferentierea limfocitelor B; ea nu exista ca organ anatomic comparabil la mamifere.
Primele raspunsuri imune umorale sumt limitate la clasele de IgM, apoi intervine rapid IgM si IgG si in final IgM, IgG, IgA. IgD sunt ca si la adulti atasate suprafetei limfocitului. Nu exista practic niciodata anticorpi circulanti din clasa IgD.
Fetusul este in principiu steril, el neavand plasmocite. Prezenta acestor celule la fetus sau la nou-nascut sunt intotdeauna rezultatul unei infectii fetale.
Studiile practicate pe soarece au demonstrat ca in absenta tuturor influentelor exterioare, limfocitele B capabile sa raspunda la diversele stimulari antigenice apar de un ordin dat si imuabil: achizitionarea repertoriului exprimat este prin urmare sub control genetic.
Ca o regula generala, reactiile de imunitate, mediate celular, apar relativ devreme in cursul gestatiei. Animalul nou-nascut sau nou-eclozionat, care provine dintr-un mediu protejat, este brusc confruntat cu mediul exterior si cu o mare varietate de microorgansime (sau de virusuri) potential patogene. Nou-nascutii sunt totusi capabili din punct de vedere imunologic sa raspunda la agresiuni, raspunsul lor fiind de tip primar, caracterizat printr-o lunga perioada de latenta si printr-o scazuta sinteza de anticorpi. Nou-nascutul este asadar protejat numai de o haina imunitara pasiva, de origine maternala, pentru a nu fi expus la infectie.
Protectia maternala, exclusiv umorala, se poate exercita la nivel general (sistemic) sau local. Dupa nastere, la mamifere protectia locala la nivelul tubului digestiv este conferita prin secretii colostrale si lactate.
Din contra, imunitatea pasiva sistemica poate fi deja, achizitionata inainte de nastere (la primate), in mare parte dupa nastere (la carnivore si rozatoare) sau integral dupa nastere (la rumegatoare, suine si equine) in functie de tipul de placentatie.
Aceste modalitati de transmitere a imunitatii maternale furnizeaza animalului tanar un nivel de anticorpi circulanti echivalent celui de la mama sa.
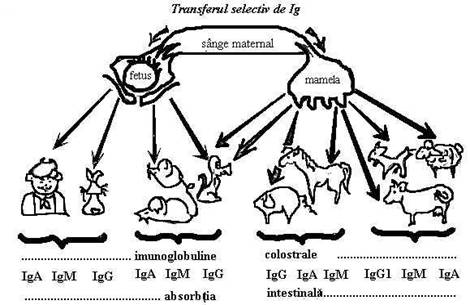
Imunitatea maternala, face parte din imunitatea dobandita pasiv, intrucat anticorpii gata formati trec de la mama la fat, fie prin filtrul placentar (imunitate congenitala, diaplacentara, transplacentara), fie prin lapte (imunitate trofogena, sau colostrala lactogena), fie prin vitelus (imunitate vitelina la pasari).
Transmiterea anticorpilor in timpul vietii intrauterine, este legata de tipul de placentatie al speciei respective. La speciile cu placentatie de tipul sindesmocorial (cal, bou, oaie, capra, magar, camila) trecerea substantelor protectoare de la mama la fat nu este posibila. Placentatia de tip hemocorial, prezenta la om, maimuta si rozatoare, usureaza trecerea anticorpilor; placentatia endoteliala (carnasierele, cerbul) ocupa un loc intermediar intre cele doua tipuri, lasand sa treaca numai o parte de anticorpi. Astfel, in functie de placentatie, de transferul de anticorpi, de la mama la fat, animalele pot fi clasificate astfel:
a) specii la care transferul de anticorpi este complet la nastere (iepure si cobai);
b) specii la care transferul de anticorpi se face atat in timpul vietii uterine, cat si dupa fatare (sobolani, soareci, carnasiere );
c) specii la care transferul de anticorpi se face exclusiv dupa fatare prin colostru si in unele cazuri prin lapte (rumegatoare, porcine, cabaline, etc.) (fig.9 si fig.10).
Laptele maternal constituie, asadar o sursa de IgG care se leaga de un receptor FcRn al celulelor epiteliale de la nivelul intestinului. Acest receptor exista sub forma de monomeri si de dimeri. Prin procesul denumit transcitoza, moleculele de IgG sunt transportate prin transferul epiteliului intestinal si intra in circulatie. Complexele rezultate din legatura dintre receptori si Ig sunt localizate in depresiunile membranei plasmatice, fiind situate preferential la baza microvililor. Receptorul FcRn se leaga de regiunea Fc formata de al doilea si al treilea domeniu constant al celor doua lanturi de imunoglobulina G. Acest receptor poseda o proprietate unica, fiind in masura sa se lege pe IgG la un pH acid (6,0), asa cum este cel existent la nivelul lumenului intestinal, si sa puna in libertate imunoglobulina la pH de 7,4, prezent in sange. Aceasta proprietate asigura transportul unidirectional al Ig din lumenul intestinal catre sange.
Studiile au demonstrat ca receptorul FcRn prezinta o stransa analogie de secventa cu moleculele de CMH din clasa I. Sinteza acestui receptor este tranzitorie si nu este prezenta decat in perioada de achizitie a imunitatii pasive, care la sobolan merge pana la trei saptamani.
La artiodactile, in special, colostrul este in mod evident mai bogat decat sangele in imunoglobula G (suinele) sau IgG1 (rumegatoarele). Totalitatea acestor Ig deriva din sangele maternal. Originea aproape exclusiv serica a IgG1 din colostru la rumegatoare este atestata prin observarea unei importante caderi a concentratiei acestei subclase de Ig in sangele maternal.
Originea aproape exclusiv serica a IgG din colostru la rumegatoare este atestata prin observarea unei importante caderi a concentratiei acestei subclase de IgG in sangele maternal, plecand de la a doua saptamana ce precede fatarea la oaie (de la 15 mg/ml la 6 mg/ml).
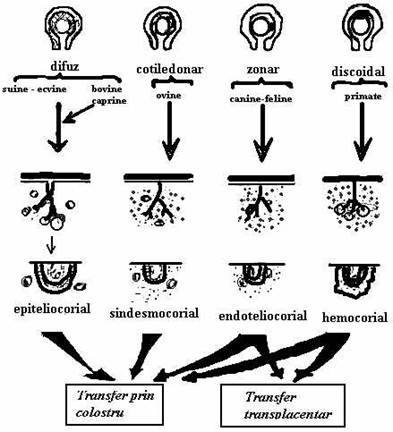
Fig.10 Diferite tipuri de placenta la
mamifere
BOHL (1975) a observat ca la scroafa care alapteaza nu se elaboreaza IgA specifice virusului gastroenteritei transmisibile a purcelului decat daca stimulii se exercita la nivelul intestinal.
El demonstreaza apoi ca la rozatoare majoritatea plasmocitelor producatoare de IgA, recunoscute in mamela, deriva din intestin. Aceste rezultate aduc proba experimentala a existentei unei legaturi imunologice intre intestin si mamela si permite explicarea faptului ca IgA din secretiile lactate sunt in speta specifice agentilor patogeni recunoscuti in lumenul intestinal.
Variatii specifice sunt observate in resorbtia intestinala a Ig. La primate, nu exista aproape nici un fel de pasaj, in timp ce intestinul purcelului nou-nascut absoarbe nespecific diferitele clase de Ig, cu exceptia IgA secretorie, care ramane fixata pe epiteliul intestinal. La vitelul nou-nascut nu exista aparent nici o selectie, toate Ig ingerate inclusiv IgA secretorie traverseaza epiteliul intestinal. Acestea nu subzista de obicei mai mult de doua zile in circulatie dupa care ele sunt eliminate printr-un proces de transudatie inversa la nivelul epiteliilor secretorii.
La rozatoare absorbtia se face gratie receptorilor pentru IgG din regiunea proximala a intestinului, unde activitatea lizozomala a celulelor este mai putin dezvoltata. Absorbtia IgG la aceste specii se continua pana la 18 - 21 de zile. La rumegatoare, la cal si la porc, absorbtia Ig este maximala in primele ore de la nastere apoi decade rapid, probabil din cauza activitatii lizozomale a celulelor epiteliale. Dupa nastere, aceasta activitate creste progresiv, antrenand digestia enzimatica a Ig., absorbtia incetand dupa 24 ore la bovine si cabaline si 36 ore la porc.
Nivelul anticorpilor maternali diminua apoi gradual cu o viteza dependenta de timpul de injumatatire al anticorpilor la specia considerata si cu viteza de crestere a acesteia.
Pe langa anticorpii specifici, principalii factori antimicrobieni nespecifici din colostru, sunt: lizozimul, lactoferina si lactoperoxidaze (BRUNO - REITER, 1978).
Colostrul vacilor contine 200 - 500 mg de lactoferina la 100 ml. Lactoferina are proprietatea de a lega Fe. Unele bacterii si in special E. Coli, au nevoie de Fe ca factor de crestere. Prin fixarea Fe pe lactoferina colostrala, germenii sunt lipsiti de acest factor si ca atare cresterea lor este inhibata (KOLB, 1981).
Progresiv secretia mamara evolueaza de la colostru catre lapte. La speciile domestice, aceasta evolutie este cu atat mai rapida cu cat rasa a fost selectionata pentru productia de lapte.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2818
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved