| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Metabolism glucidic
In cadrul metabolismului intermediar, procesele biochimice din metabolismul glucidic ocupa un loc central.
Glucoza reprezinta cel mai important "combustibil" al organismului, chiar daca prin oxidarea biologica a acesteia nu se elibereaza cea mai mare cantitate de energie. Pentru diferite organisme obtinerea glucozei necesare desfasurarii activitatii se realizeaza in mod diferit: organismele autotrofe isi constituie glucoza prin fotosinteza, in timp ce pentru organismele heterotrofe glucoza are origine exogena (luata din alimente mai ales sub forma de dizaharide si polizaharide) sau se sintetizeaza din compusi neglucidici (aminoacizi sau glicerol) prin procesul de gluconeogeneza.
Glucoza sintetizata endogen asigura doar partial necesarul zilnic, de aceea trebuie introdusa prin alimentatie. Se apreciaza ca organismul uman adult necesita zilnic 200-250 g glucoza pentru desfasurarea unui regim de activitate normal.
Glucidele introduse prin alimentatie (amidon, lactoza, zaharoza) nu se pot utiliza ca atare si, de aceea, ele trebuie sa fie metabolizate pana la glucoza, in principal, care este transportata apoi de vena porta la ficat unde poate fi depozitata sub forma de glicogen, catabolizata pentru obtinerea energiei necesare, redistribuita altor tesuturi sau convertita in compusi endogeni in functie de necesitatile organismului.
Surse exogene de glucoza sunt: amidonul (preluat din cartofi, orez, paine, etc ce constituie aproximativ 50% din glucidele ingerate), zaharoza (aproximativ 25%, preluata mai ales din zahar), lactoza (cca.10%, preluata din lapte), glucoza si fructoza in cantitate redusa (5%, preluate din fructe dulci, miere), celuloza (2%, luata din legume si fructe).
De-a lungul tractului gastrointestinal o parte din glucide (amidon, zaharoza, lactoza) sufera o serie de transformari inainte de a fi absorbite, altele (glucoza, fructoza) se absorb direct, in timp ce celuloza se elimina nehidrolizata, rolul acesteia in procesul de digestie fiind de a contribui la formarea bolului alimentar si de a stimula peristaltismul intestinal.
Hidroliza amidonului incepe la nivelul cavitatii orale sub actiunea amilazei salivare (ptialina) si continua in intestin sub actiunea amilazei pancreatice. Amilazele salivara si pancreatica sunt -amilaze care scindeaza legaturile -1,4-glicozidice interne actionand ca endoamilaze. Acestea hidrolizeaza componentele amidonului, amiloza si amilopectina, dar si glicogenul, eliberand maltoza. Deoarece amilazele nu pot hidroliza legaturile 1,6-glicozidice, produsii de hidroliza sunt dextrine (oligozaharide).
Hidroliza dizaharidelor in intestin este cataliazata de dizaharidaze specifice sintetizate de marginea in perie a mucoasei intestinale.
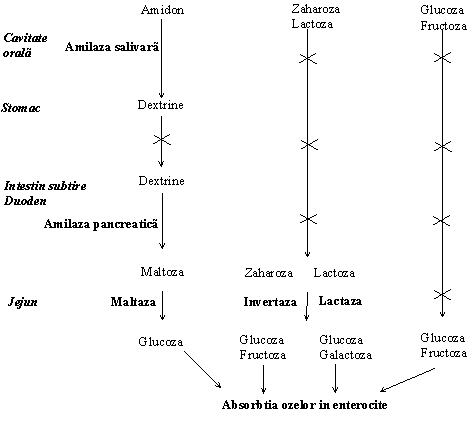
Figura Reprezentarea etapelor din digestia glucidelor in tractul gastrointestinal
Absorbtia ozelor rezultate dupa hidroliza enzimatica a di- si polizaharidelor din lumenul intestinal in enterocite se face prin doua mecanisme:
a) transport activ (realizat cu consum de energie) - pentru hexoze;
b) difuzie facilitata, fara consum de energie, pentru pentozele rezultate din pentozani, produsi de policondensare a ozelor continuti in coaja unor fructe (cirese, visine, prune).
Transportul glucozei din lumenul intestinal in enterocite se face prin cotransport simport cu ionii Na+ (pompa de sodium si glucoza), dependent de gradientul de Na+ stabilit de Na+, K+- ATP - aza din membrana plasmatica (pompa de Na+ si K+).
Transportorul simport de Na+ si glucoza de la polul apical al celulei intestinale preia din intestin glucoza odata cu patrunderea a doi ioni Na+ in celula, glucoza putandu-se acumula pana la o concentratie de 30000 ori mai mare decat in lumenul intestinal. Pe masura ce glucoza este pompata in enterocite, ea este eliminata simultan la polul bazal prin difuzie facilitata de transportorii GluT2 aflati in membrana enterocitelor.
Ionii de Na+ captati in enterocite sunt eliberati la polul bazal de Na+, K+- ATP - aza din membrana ce scoate din enterocit 3 ioni Na+ si introduce in celula 2 ioni K+.
Din enterocite, ozele sunt transportate de vena porta la ficat care le metabolizeaza specific. In tesuturi, glucoza poate fi folosita in functie de necesitatile celulelor: a) catabolizata pentru obtinerea energiei atunci cand organismul este in activitate; b) depozitata sub forma de glicogen in ficat si muschi sau c) convertita in compusi endogeni, in conditiile unui exces de glucoza.
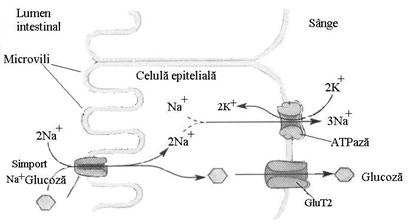
Figura. Transportul simport al glucozei in epiteliul intestinal
(dupa Nelson si Cox , 2000)
i Corelatii clinice Digestia diglucidelor poate fi afectata in cazul unor enzimopatii determinate de deficitul in dizaharidaze, ceea ce determina intoleranta la dizaharide (lactoza, zaharoza). In aceste conditii, dizaharidele se acumuleaza in intestin, determina cresterea presiunii osmotice in lumenul intestinal, ceea ce are ca rezultat pierderile digestive de apa, diaree, dureri abdominale (deoarece flora bacteriana sintetizeaza acid lactic). Simptomele se pot ameliora prin evitarea consumului de astfel de glucide (de ex., consumul de lapte prelucrat pentru indepartarea lactozei).
Glucoza din mediul extracelular este neutra si poate patrunde in celule prin difuzie. In acest proces un rol important revine transportorilor GluT, permeaze specifice cu afinitate variabila pentru glucoza si distribuite diferit in tesuturi, ce realizeaza un transport uniport al glucozei. Exista mai multi transportori GluT care se diferentiaza prin afinitatea pentru substrat, glucoza (GluT1 in eritrocit cu afinitate mare, GluT2 in ficat si intestin cu afinitate mai mica, GluT4 in muschi, tesut adipos, care este stimulat de insulina, hormonul hipoglicemiant).
Pentru a fi mentinuta in celule, glucoza este fosforilata si transformata in glucozo-6-fosfat, compus incarcat negativ care nu mai poate strabate membrana in sens invers. Reactia este ireversibila, reprezentand o etapa importanta in reglarea metabolismului glucozei.
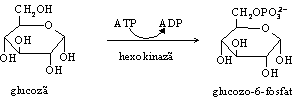
Hexokinaza (HK) este activa in toate celulele si are afinitate mare pentru glucoza (Km=0,01mM). Prezinta mai multe izoenzime si poate fosforila si alte oze: fructoza, manoza. Nivelul crescut al glucozo-6-fosfatului (G-6-P) inhiba activitatea hexokinazei.
In ficat, reactia este catalizata si de glucokinaza (GK), izoenzima a HK ce are afinitate mica pentru glucoza (Km=10mM) si care fosforileaza numai glucoza. Enzima este activa numai cand organismul are consum crescut de glucide (postprandial) si formeaza G-6-P pe care ficatul il depoziteaza sub forma de glicogen. Nu este inhibata de G-6-P si este indusa de insulina.
Glucozo-6-fosfatul este un intermediar comun al mai multor cai metabolice si ocupa un rol central in metabolismul glucozei (Figura ).
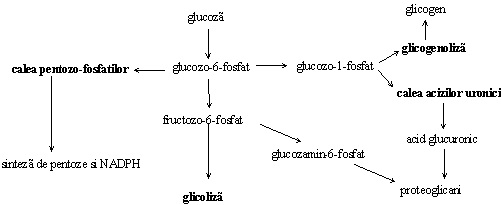
Figura. Pozitia glucozo-6-fosfatului in directionarea proceselor din metabolismul glucidic
Glicoliza este procesul prin care glucoza este degradata pana la piruvat, in conditii aerobe, sau pana la lactat, in conditii anaerobe.
Este prima cale metabolica a carei secventa a fost elucidata in prima parte a secolului 20 de catre Warburg, Meyerhoff si Embden, de unde si numele de calea Embden - Meyerhoff.
Din punct de vedere intracelular, enzimele ce intervin in acest proces sunt localizate in citoplasma tuturor celulelor.
Degradarea glucozei pana la piruvat, respectiv lactat se face prin 10, respectiv 11 etape impartite, in functie de intermediarii pe care-i contin, astfel:
faza1 - faza esterilor hexozofosforici, etapele 1-5;
faza 2 - faza esterilor triozofosforici - etapele 6-11.
Secveta glicolitica este urmatoarea:
Fosforilarea glucozei, prezentata anterior, furnizeaza glucozei energia pentru a fi metabolizata ulterior, prin transferul grupei fosfat din ATP ( G'0 = -16,7 kJ/mol).
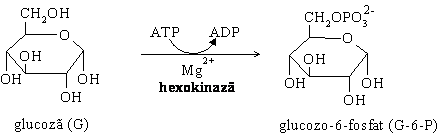
2. Izomerizarea glucozo-6-fosfatului la fructozo -6- fosfat (F-6-P) este o reactie reversibila catalizata de glucozo-6-fosfat izomeraza. Echilibrul reactiei este deplasat spre stanga, astfel ca viteza de formare a fructozo-6-fosfatului este franata ( G'0 =1,7 kJ/mol).
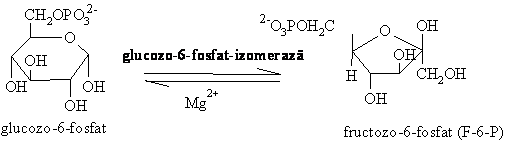
70% 30%
3. Fosforilarea fructozo-6-fosfatului este o reactie importanta in reglarea glicolizei, catalizata de o kinaza ce transfera grupa fosfat din ATP, fosfofructokinaza - 1 (FFK-1). Reactia este ireversibila ( G'0 -14 kJ/mol).
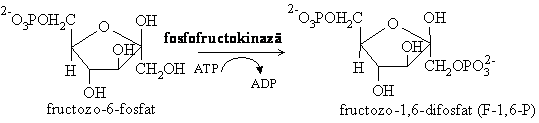
FFK-1 este o enzima allosterica, tetramer, ce se gaseste sub doua forme interconvertibile: forma relaxata (R) activa si forma tensionata (T) inactiva. Enzima prezinta doua izoenzime: FFK tip M ce se gaseste predominant in muschi, creier, ficat si FFK tip L care predomina in ficat, rinichi, tesut adipos. Aceasta enzima este punctul major de reglare a glicolizei. Efectorul allosteric pozitiv al acestei enzime este fructozo-2,6-difosfatul (format sub actiunea FFK-2 in ficat sub controlul glucagonului) ce mareste fluxul de glucoza spre glicoliza, iar efectori allosterici negativi sunt ATP-ul si acidul citric.
Scindarea fructozo-1,6-difosfatului se face sub actiunea fructozo -1,6- bisfosfat aldolazei si reprezinta reactia inversa unei condensari aldolice.

88% 12%
Deoarece aceasta reactie are ΔG>0 ( G = 23,8 kJ/mol) inseamna ca reactia se produce spontan de la dreapta la stanga. Aldolaza are mai multe izoenzime: aldolaza A (prezenta in toate celulele), aldolaza B (in ficat, rinichi) si aldolaza C (in creier, inima).
5. Conversia triozofosfatilor. Cei doi triozofosfati sunt intr-un echilibru catalizat de o triozofosfat izomeraza. Dihidroxiacetonfosfatul (DHAP) este un rezervor metabolic din care se formeaza treptat gliceraldehid-3-fosfat (G-3-P) (ΔG > 0, ΔG = 7,5 kJ/mol).

In reactiile ce se desfasoara in prima faza a glicolizei se consuma 2 moli ATP.
Faza a doua include reactiile prin care energia legaturilor C-C din glucoza este convertita in energia din ATP.
Oxidarea fosforilanta a gliceraldehid-3-fosfatului. Aceasta reactie presupune oxidarea grupei aldehidice de la C1 la grupa carboxil si fixarea unui rest fosfat sub forma de anhidrida. Anhidrida formata are o energie libera foarte mare la hidroliza ( G - 49,3 kJ/mol).

Enzima este un tetramer ce are in centrul activ o grupa -SH dintr-un rest de cisteina. G-3-P se leaga covalent la grupa -SH a enzimei oxidandu-se apoi pe seama NAD+ aflat, de asemenea, in centrul activ al enzimei. Este o etapa foarte importanta deoarece odata cu oxidarea G-3-P are loc formarea unei grupe fosfat macroergice.
Hidroliza 1,3-difosfogliceratului cu transferul grupei fosfat pe ADP, in prezenta fosfoglicerat kinazei, rezultand 3-fosfoglicerat (3-PG) ( G = -18,5 kJ/mol).
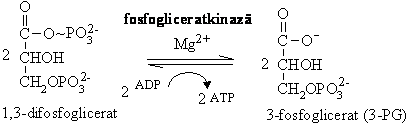
Pentru ca energia din ATP provine din oxidarea unui substrat, gliceraldehid-3-fosfatul, reactiile 6 si 7 constituie un proces de cuplare energetica cu 1,3-DPG drept intermediar comun, care transforma o reactie imposibila termodinamic intr-un proces posibil. Formarea ATP prin transferul grupei fosfat de la un substrat se numeste fosforilare la nivel de substrat, fiind un mecanism diferit de cel al fosforilarii oxidative cuplate cu respiratia celulara.
In eritrocit calea glicolitica este deviata la acest nivel de reactia:
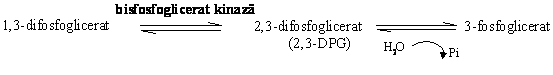
Desi in alte celule 2,3-DPG este in urme, in eritrocit concentratia sa este 5 mM, fiind un important modulator allosteric al oxigenarii hemoglobinei, legarea sa la aceasta scazand afinitatea hemoglobinei pentru oxigen.
Izomerizarea 3-fosfogliceratului la 2-fosfoglicerat (2-PG) in prezenta fosfoglicerat mutazei, enzima dependenta de prezenta ionilor Mg2+ ( G = 4,4 kJ/mol).

Deshidatarea 2-fosfogliceratului. Enolaza catalizeaza o reactie reversibila prin care rezulta al doilea intermediar macroergic al glicolizei, fosfoenolpiruvatul (PEP) ( G'0 = 7,5 kJ/mol).

Fosfoenolpiruvatul (PEP) este unul dintre compusii cu cel mai inalt nivel de energie.
Hidroliza fosfoenolpiruvatului cu transferul grupei fosfat pe ADP se desfasoara sub actiunea piruvat kinazei, enzima ce necesita prezenta ionilor K+ si Mg2+. Si aceasta este o reactie de fosforilare la nivel de substrat realizata pe seama energiei inmagazinate ( G'0 = -31,4 kJ/mol). Aceasta reactie este un punct important de reglare a glicolizei, enzima fiind inhibata de ATP si activata de fructozo-1,6-difosfat.
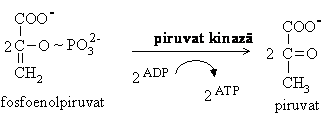
Piruvat kinaza are doua izoenzime: tip M care se gaseste in muschi si tip L ce actioneaza predominant in ficat.
Piruvatul rezultat din aceasta etapa este folosit in mod diferit in functie de disponibilitatea in oxigen a tesutului. In conditii de aerobioza, aceasta este ultima etapa glicolitica, piruvatul rezultat patrunde in mitocondrii si este convertit in acetilCoA ce va fi degradata complet pana la CO2 si H2O prin respiratie celulara, procese prezentate ulterior.
Metabolizarea piruvatului in conditii anaerobe se desfasoara diferit la diverse organisme. La drojdii metabolizarea se continua prin fermentatie, iar la alte microorganisme si la organismele superioare prin transformare in lactat.
In celulele fara mitocondrii (hematia) sau in conditii de deficit de oxigen (cornee, muschi in contractie), piruvatul este metabolizat in continuare intr-o alta etapa din glicoliza.
Reducerea piruvatului sub actiunea lactat dehidrogenazei este reactia prin care se reoxideaza NADH format sub actiunea gliceraldehid-3-fosfat dehidrogenazei. Celulele care nu au mitocondrii nu il pot reoxida prin lantul respirator, iar in conditiile unei activitati musculare intense se produce mai mult NADH decat poate fi reoxidat. Astfel se regenereaza NAD+ necesar pentru desfasurarea continua a glicolizei.
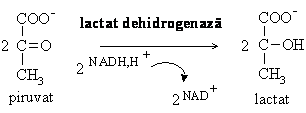
Lactatul acumulat in muschi este responsabil de crampele si febra musculara, fiind preluat de ficat si utilizat pentru gluconeogeneza.
Metabolizarea anaeroba a piruvatului prin fermentatie
In aceste procese, piruvatul se oxideaza cu formare de ATP prin reactii in care moleculele organice functioneaza si ca donori si ca acceptori de electroni.
Fermentatia alcoolica este procesul biochimic ce sta la baza obtinerii berii si vinului:

![]()
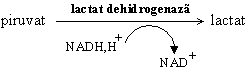
Lactatul produs prin fermentatia lactica de unele microorganisme este responsabil de gustul amar al laptelui acru si de gustul si mirosul sucului de varza murata:
|
Reactia |
ATP consumat |
ATP sintetizat |
|
1. glucoza àglucozo-6-fosfat | ||
|
3. Fructozo-6-fosfatà fructozo-1,6-difosfat | ||
|
7. 1,3-difosfogliceratà 3-fosfoglicerat | ||
|
10. Fosfoenolpiruvat à Piruvat | ||
|
Bilant |
In cazul in care degradarea incepe de la glicogen se vor forma 3 moli de ATP pentru ca glucozo-6-fosfatul rezulta direct din glicogen prin fosforoliza, deci fara consum de ATP.
Din cele 11 etape ale glicolizei, 3 sunt ireversibile si sunt catalizate de enzime ce au rol in controlul sensului pe care il ia transformarea intermediarilor glicolitici in functie de necesitatile organismului:
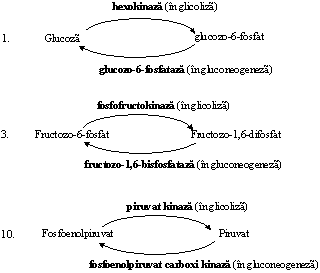
Glucozo-6-fosfatul, ATP, acetilCoA, citratul sunt inhibitori ai unor enzime din glicoliza, iar AMP, fructozo-2,6-bisfosfatul si fructozo-1,6-bisfosfatul sunt activatori.
Corelatii clinice
a. Procese tumorale. Captarea glucozei si glicoliza se desfasoara mult mai rapid in tesutul tumoral decat in cele necanceroase, deoarece initial acestora le lipseste o retea intensa de capilare, fiind obligate sa functioneze in conditii de hipoxie. In plus acestea au si un numar redus de mitondrii pentru a realiza o degradare completa a glucozei. Acestea produc in cantitate crescuta anumite enzime glicolitice, intre care o izoenzima a hexokinazei care nu este inhibata de glucozo-6-fosfat. Sinteza excesiva a enzimelor glicolitice este controlata de proteinele codificate de oncogene sau genele supresoare tumorale, de exemplu p53.
b. Glucokinaza este o enzima a carei actiune este stimulata de insulina. In diabet zaharat, actiunea enzimei este deficitara datorita reglarii ineficiente.
Si alte oze pot sa participe la glicoliza prin transformarea in intermediari glicolitici.
Fructoza obtinuta din hidroliza zaharozei poate alimenta glicoliza in mod diferit in tesuturi.

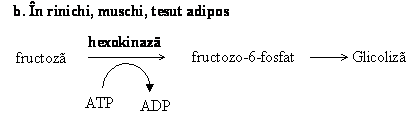
Manoza, componenta importanta a heteropoliglucidelor, poate alimenta glicoliza in acelasi mod ca si fructoza.
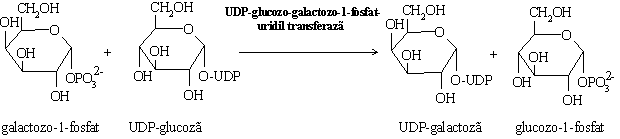
 Galactoza are o modalitate particulara de a se transforma in glucoza si a
intra in glicoliza.
Galactoza are o modalitate particulara de a se transforma in glucoza si a
intra in glicoliza.
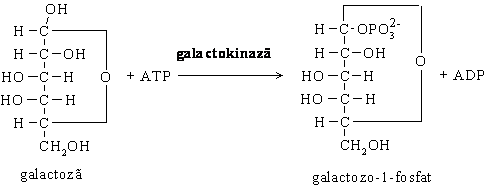
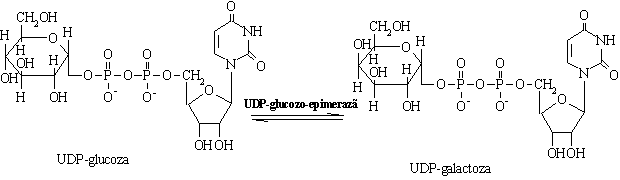
a) Deficitul de fructokinaza este caracterizat de metabolizarea ineficienta a fructozei cu eliminarea acesteia in urina, boala numita fructozurie esentiala.
b) Calea poliol este o cale metabolica de transformare a ozelor in care intervin doua enzime: aldozo reductaza, enzima care transforma glucoza in sorbitol si sorbitol dehidrogenaza, enzima ce intervine in transformarea sorbitolului in fructoza.
In conditii normale, transformarea glucozei pe aceasta cale este redusa (sub 5%), metabolizarea intensificandu-se in conditii de hiperglicemie, de exemplu in diabet zaharat. In cazul intensificarii procesului, produsii caii poliol se acumuleaza la nivelul unor structuri precum retina, nervi, rinichi si determina alterari structurale si functionale.
Limitarea transformarii pe aceasta cale se poate realiza prin folosirea unor medicamente ce functioneaza ca inhibitori ai aldozo reductazei.
c) Galactozemia esentiala este o enzimopatie determinata de deficitul de galactozo-1-fosfat-uridil transferaza sau de galactokinaza cand se acumuleaza galactoza, acumulare insotita de cataracta si afectiuni neurologice.
Degradarea glucozei prin glicoliza, ciclul Krebs si lantul respirator este un proces care furnizeaza o cantitate mare de energie, variabila in functie de sistemele transportoare ale echivalentilor de reducere (suveici malat-aspartat si glicerol-3-fosfat) ce functioneaza in tesut.
Bilantul total al oxidarii glucozei
|
Suveica glicerol-3 fosfat |
Suveica malat |
|
|
Glicoliza Fosforilarea glucozei Fosforilarea fructozo-6-P Defosforilarea 1,3-DPG Defosforilarea PEP Total ATP glicoliza Oxidarea GA-3P duce la 2NADH,H+ |
-1 -1 -1 -1 2 2 2 2 2 2 |
|
|
Decarboxilarea acidului piruvic duce la 2NADH,H+ |
||
|
Ciclul Krebs |
||
|
2 GTP à 2 ATP |
2 |
2 |
|
Oxidarile a 2moli izocitrat, cetoglutarat, malatà NADH,H+ | ||
|
Oxidarea a 2 moli succinat à 2 FADH2 | ||
|
Lantul respirator si fosforilarea oxidativa | ||
|
2NADH,H+ din oxidarea G-3-P |
2 x 1,5 = 3 |
2 x 2,5 = 5 |
|
2NADH,H+ din decarboxilarea acidului piruvic |
2 x 2,5 = 5 |
2 x 2,5 = 5 |
|
6NADH,H+ din ciclul Krebs |
6 x 2,5 = 15 |
6 x 2,5 = 15 |
|
2FADH2 |
2 x 1,5 = 3 |
2 x 1,5 = 3 |
|
Total | ||
Reactia finala:
Glucoza + 6 O2 + 32ADP + 32Pi à 6CO2 + 32 ATP +38 H2O
Componenta exergonica Glucoza + 6 O2 à 6CO2 + 6 H2O G0' = - 2937 kJ/mol
Componenta endergonica 32ADP + 32Pi à 32 ATP +32 H2O G0' = 1600 kJ/mol
= 100 x 1600/2937 = 54%
In conditii aerobe eficienta procesului este mult mai mare. Astfel se explica efectul Pasteur, prin aerobioza rezulta 32 moli ATP iar prin anaerobioza 2 moli, deci in conditii de anaerobioza cantitatea de glucoza consumata este de 16 ori mai mare.
Procese metabolice in faza de absorbtie (postprandial)
Postprandial, organismul are un exces de substraturi energogene si trebuie sa desfasoare reactii prin care inmagazineaza energia pe care acestea o inglobeaza.
Glicogenogeneza este procesul biochimic prin care se realizeaza sinteza glicogenului, poliglucid de rezerva de origine animala. Procesul de glicogenogeneza este un proces anabolic ce se realizeaza atunci cand exista surplus de glucoza si contribuie alaturi de glicogenoliza la mentinerea glicemiei.
In 1957, Leloir si colaboratorii au demonstrat ca biosinteza glicogenului se desfasoara in ficat si mai putin in muschi, dar pe o cale complet diferita de cea a degradarii glicogenului, proces ce se realizeaza, de asemenea, in organele mentionate.
Sinteza glicogenului are loc in fibrele musculare albe numai in perioadele de repaus cand glucoza este in exces. In timpul contractiei musculare, glicogenul este degradat pentru a se obtine glucoza si, apoi, ATP.
Pentru sinteza glicogenului glucoza trebuie sa fie activata, sub forma de UDP-glucoza.
![]()

Reactia de activare devine ireversibila deoarece, in vivo, se formeaza fosfat prin degradarea pirofosfatului sub actiunea pirofosfatazei:
![]()
Sinteza glicogenului se realizeaza prin atasarea resturilor glucozil de pe UDP-glucoza pe o molecula de glicogen "amorsa" (glicogen (n-1)). Formarea glicogenului primer o realizeaza glicogenina (o proteina cu M 37,2 kDa) care functioneaza autocatalitic.
![]()
In final, UTP-ul necesar activarii se reface prin reactia:
![]()
Sistemele enzimatice specifice glicogenogenezei sunt:
a) Glicogen sintaza este enzima ce catalizeaza transferul unor grupe glucozil activate pe o molecula de glicogen "amorsa" (primer) formand legaturi 1,4-glicozidice putand adauga maxim 10 resturi glucozil. Este o enzima care se prezinta sub doua forme interconvertibile prin fosforilare-defosforilare, glicogen sintaza D, fosforilata (inactiva) si glicogen sintaza I, nefosforilata (activa);
b) Enzima de ramificare (amilo α1,4 -α1,6-transglucozidaza) enzima ce scindeaza un bloc de 6-7 resturi glucozil de pe un lant in crestere si le transfera pe un alt lant realizand o legatura 1,6-glicozidica. Noua ramificatie trebuie sa se gaseasca la o distanta de cel putin 4 resturi glucozil fata de ramificatia adiacenta.
Ecuatia glicogenogenezei este:
Glicogen(n-1) + Glucoza + 2ATP à Glicogen(n) + 2ADP + 2Pi
Este o cale particulara de metabolizare a glucozo-6-fosfatului care ocoleste o serie de etape glicolitice. Se mai numeste suntul pentozofosfatilor sau calea fosfogluconatului.
Functii biologice:
Este o cale metabolica prin care din glucozo-6-fosfat rezulta derivati ai ozelor cu trei, patru, cinci, sase, sapte atomi de carbon. Este singura cale prin care rezulta riboza si deoxiriboza necesare in sinteza acizilor nucleici.
Procesul metabolic presupune initial desfasurarea a doua etape de oxidare catalizate de enzime NADP+- dependente. Una dintre enzime este glucozo -6- fosfat dehidrogenaza care catalizeaza prima reactie a caii metabolice, aceea de oxidare a glucozo-6-fosfatului odata cu reducerea coenzimei:
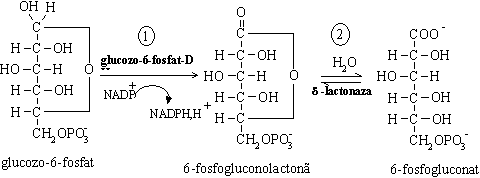
De aceea, este principala cale prin care se formeaza NADPH,H+ necesar biosintezelor reductive (de acizi grasi, colesterol si alti derivati sterolici), hidroxilarii compusilor straini organismului prin sistemul oxidazelor cu functie mixta, refacerii glutationului redus (GSH);
Determina formarea de triozofosfati importanti pentru metabolismul lipidic.
Ocoleste etapele consumatoare de energie din glicoliza.
Localizare intracelulara si tisulara Reactiile caii pentozofosfatilor se desfasoara in citoplasma deoarece aici se desfasoara procesele la care participa NADPH,H+.
Procesul se desfasoara cu intensitate maxima in tesuturile in care lipogeneza si sinteza hormonilor steroizi sunt accelerate: ficat, tesut adipos, glande mamare, corticosuprarenala precum si in eritrocit. Enzimele acestei cai lipsesc in miocard, muschi scheletici, creier, rinichi.
Calea pentozofosfatilor se desfasoara in doua etape majore:
a. conversia hexozelor la pentoze;
b. conversia pentozelor la hexoze.
Secventa caii pentozofosfatilor
I. Conversia hexozelor la pentoze
Oxidarea glucozo-6-fosfatului in prezenta glucozo-6-fosfatdehidrogenazei
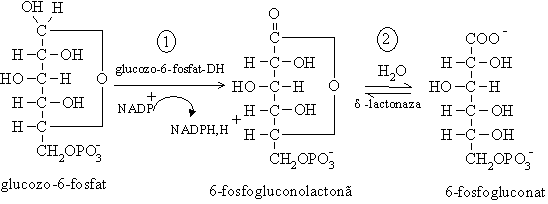
Glucozo-6-fosfat dehidrogenaza este inhibata de NADPH,H+ si de esterii acizilor grasi si coenzima A.
Decarboxilarea oxidativa a 6-fosfogluconatului in prezenta 6-fosfogluconat dehidrogenazei:
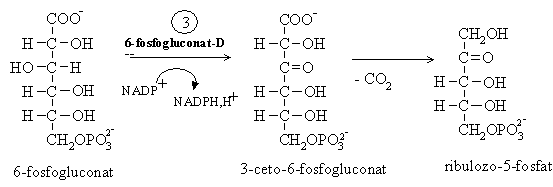
Enzima este inhibata de fructozo-1,6-bisfosfat care impiedica transformarea ulterioara a pentozelor in hexoze.
Izomerizarea ribulozo-5-fosfatului

Ecuatia generala a primei etape:
Glucozo-6-fosfat + 2NADP+ + H2O à ribulozo-5-fosfat + 2NADPH,H+ + CO2
II. Conversia pentozelor la hexoze include reactii de izomerizare, transcetolizare, transaldolizare:
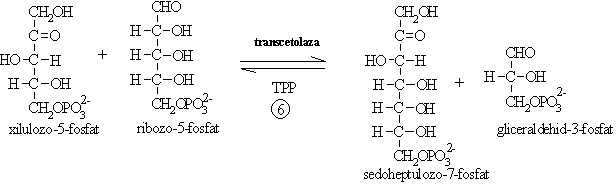
Transcetolaza transfera o grupare cu doi atomi de carbon ce contine si o grupare cetonica de pe o cetoza pe o aldoza.
Conversia sedoheptulozo-7-fosfatului
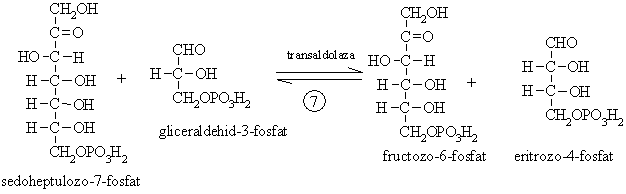 Conversia
eritrozo-4-fosfatului
Conversia
eritrozo-4-fosfatului

Etapele 4,5,6,7,8 sunt reversibile pentru ca atunci cand celula are nevoie de mai mult NADPH,H+ si nu de ribozo-5-fosfat sa se poata desfasura procesul in sens invers pana la intermediarii glicolitici.
Utilizarea glucozo-6-fosfatului fie prin glicoliza fie pe calea pentozofosfatilor depinde de necesitatile celulei pentru ATP, NADPH si ribozo-5-fosfat.
a. Daca celula are nevoie si de NADPH,H+ si de ribozo-5-fosfat se desfasoara predominant primele 4 reactii.
b. Daca celula necesita mai mult ribozo-5-fosfat decat NADPH, H+ sunt extrase din glicoliza fructozo-6-fosfatul si gliceraldehid-3-fosfatul pentru a ocoli etapele din calea pentozofosfatilor unde se sintetizeaza NADPH,H+. Se produc doar reactiile 5, 6, 7, 8.
c. Daca este necesar mai mult NADPH,H+, ribozo-5-fosfatul este folosit pentru a produce intermediari glicolitici pentru ca glucozo-6-fosfatul sa fie folosit in producerea de NADPH,H+.
Reactia generala pentru aceste situatii ar putea fi:
![]()
Corelatii clinice. NADPH,H+ sintetizat in calea pentozofosfatilor este folosit si la reducerea glutationului oxidat:

Aceste reactii prezinta importanta pentru eritrocit, deoarece asigura glutationul redus necesar descompunerii H2O2 care in cantitate mare ar determina oxidarea hemoglobinei la methemoglobina si stres oxidativ. H2O2 provine din actiunea radiatiilor ionizante asupra unor compusi endogeni, din degradarea xenobioticelor (substante straine organismului), din unele procese biochimice (ex., degradarea bazelor purinice etc.)
Deficitul de glucozo-fosfat dehidrogenaza determina anemia hemolitica. S-a observat ca unii subiecti sunt susceptibili la a manifesta anemie hemolitica acuta la administrarea unor medicamente ca antimalarice, sulfamide, antipiretice, simptomele aparand in 48-96 ore, datorita deficitului de activitate a acestei enzime.
Incidenta acestei deficiente este crescuta in zonele geografice unde poate sa apara malaria.

Calea acizilor uronici reprezinta o cale de metabolizare a glucozo-6-fosfatului cu formare de acid glucuronic si xiluloza. Importanta biologica a acestei cai consta in:
a) Genereaza glucuronatul necesar pentru: i) procese de detoxifiere hepatica ce presupun conjugarea fenolilor, hormonilor, bilirubinei, a unor medicamente pentru marirea solubilitatii in apa si reducerea toxicitatii; ii) sinteza proteoglicanilor.
b) La plantele superioare si la mamifere, cu exceptia omului, primatelor, cobaiului, este calea de sinteza a acidului ascorbic (vitamina C).
Secventa cai acizilor uronici:
1. ![]()
2. Oxidarea glucozei

UDP-glucuronatul poate transfera radicalul glucuronil pe diversi acceptori in sinteza proteoglicanilor sau formarea glucuronoconjugatilor cu rol in marirea solubilitatii in apa a unor compusi endogeni sau exogeni.
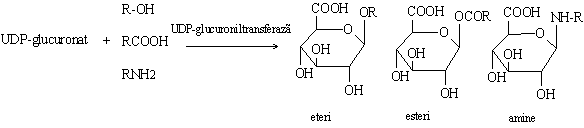
Glucuronatul liber se poate transforma pe doua cai:
a. transformarea glucuronatului in xiluloza;
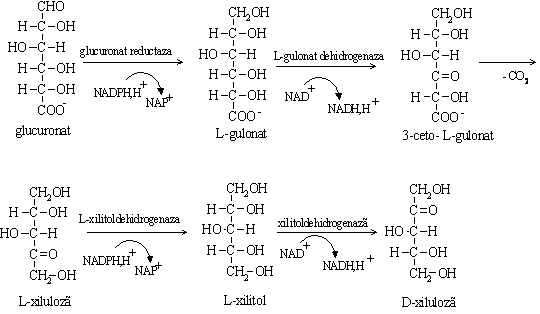
i Corelatii clinice. Lipsa xilitoldehidrogenazei (NADP+) dependenta determina eliminarea prin urina a L-xilulozei, caracteristica pentozuriei esentiale.
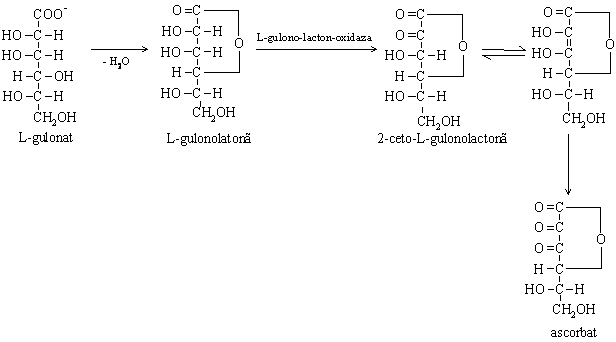
b. transformarea glucuronatului in acid ascorbic
Reprezinta o cale de metabolizare intalnita la plante si mamifere cu exceptia primatelor, cobaiului si omului. Este localizata in microzomii hepatici.
Enzima L-gulonolactonoxidaza lipseste la mamiferele mentionate care sunt dependente de aportul de vitamina C.
Procese metabolice in faza de repaus alimentar (interprandial)
Interprandial, organismul trebuie sa consume rezervele energetice dobandite din aportul in exces de glucoza din faza de absorbtie sau sa aiba capacitatea sa sintetizeze "de novo" glucoza necesara din compusi neglucidici.
Alimentarea glicolizei interprandial se face folosind ca materie prima glicogenul ce se degradeaza prin procul de glicogenoliza.
Localizare tisulara. Procesul se desfasoara in ficat si in muschiul in contractie.
Degradarea glicogenului presupune scindarea legaturilor 1,4- si 1,6-glicozidice prin actiunea conjugata a doua sisteme enzimatice distincte:
a) Sistemul fosforilazic care cuprinde enzimele ce intervin in activarea fosforilazei, enzima ce scindeaza legaturile 1,4-glicozidice din glicogen prin fosforoliza reactie ce implica transferul unui rest fosfat pe un rest glucozil, cu formare de glucozo-1-fosfat;
b) Enzima de deramificare - are capacitatea de a transfera o unitate triglucidica de pe un lant pe altul formand legaturi 1,6-glicozidice si de a scinda restul de la zona de ramificatie.
Fosforilaza a actioneaza asupra capatului terminal nereducator al lantului poliglucidic si scindeaza legatura 1,4-glicozidica cu formare de glucozo-1 fosfat.
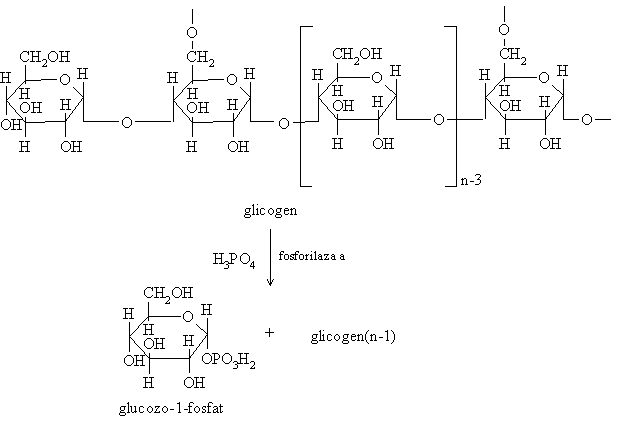
Fosforilaza nu poate sa desfaca legaturile 1,6-glicozidice actiunea sa oprindu-se la 4 resturi glucozil fata de o ramificatie 1,6. La acest nivel actioneaza cel de-al doilea sistem enzimatic enzima de deramificare care are proprietatea de a transfera o unitate triglucidica de pe un lant pe altul.
In zona de ramificatie ramane un singur rest glucozil care va fi scindat de aceeasi enzima ce are si actiune 1,6-glucozidazica.
Se continua apoi activitatea fosforilazei pana in apropierea unei noi ramificatii.
Glucozo-1-fosfatul rezultat este izomerizat la glucozo-6-fosfat ce poate fi utilizat specific in functie de tesut (degradat prin glicoliza).
Intre actiunea fosforilazei la nivel muscular si hepatic exista diferente notabile care sunt dependente de concentratia glucozei sanguine si de actiunea unor hormoni:
adrenalina - ce determina activarea fosforilazei musculare;
glucagonul - care stimuleaza activarea fosforilazei hepatice.
In ficat, glucozo-6-fosfatul rezultat prin izomerizarea glucozo-1-fosfatului este defosforilat sub actiunea glucozo-6-fosfatazei rezultand glucoza care intra in circulatia sanguina si restabileste nivelul glicemiei, fiind apoi distribuita tesuturilor.
In muschi, glucozo-6-fosfatul intra direct in glicoliza pentru obtinerea energiei proprii muschiului. Din punct de vedere energetic, glicogenoliza este un proces mai eficient decat glicoliza, bilantul net fiind de 3 moli de ATP.
Ecuatia glicogenolizei este:
Glicogen(n) + 3ADP + Pi à Glicogen(n-1) + 3 ATP
Catabolismul glicogenului nu este avantajos din punct de vedere energetic. Totusi organismul realizeaza acest proces, deoarece glicogenul muscular este mai rapid metabolizat, grasimile neconstituind sursa de energie in lipsa oxigenului si neputand contribui la mentinerea glicemiei necesare unor tesuturi glucodependente (hematie, creier).
Reglarea glicogenolizei si glicogenogenezei.
Cele doua procese se afla sub controlul adrenalinei, glucagonului si insulinei, astfel incat ele sa nu se produca simultan.
Fosforilaza a reprezinta punctul principal de control al glicogenolizei, iar glicogen sintaza al glicogenogenezei.
Insulina stimuleaza glicogenogeneza, adrenalina si glucagonul determina o succesiune de reactii ce stimuleaza glicogenoliza si inhiba glicogenogeneza.
Excitarea nervoasa a activitatii musculare este mediata de modificarea concentratiei ionilor Ca2+. Impulsul nervos determina depolarizarea membranei, eliberarea ionilor Ca2+din reticulul sarcoplasmic si contractia musculara. In plus, ionii Ca2+ pot sa semnalizeze activarea glicogen fosforilazei si inactivarea glicogen sintazei, astfel ca se va degrada mai mult glicogen pentru a furniza ATP-ul necesar contractiei musculare.
Corelatii clinice. Deficienta ereditara a unor sisteme enzimatice implicate in metabolismul glicogenului determina aparitia unor boli ereditare denumite glicogenoze, caracterizate prin tezaurizarea in diverse tesuturi unor cantitati mari de glicogen cu structura normala sau anormala in unele tesuturi (ficat, rinichi, intestin, muschi, creier, etc), fiind insotite de hipoglicemie, acidoza lactica. Sunt noua tipuri de glicogenoze (I-IX). Sunt noua tipuri de glicogenoze notate I-IX, de exemplu:
Maladia Von Gierke (tip I) - deficit de glucozo-6-fosfataza;
Maladia Pompe (tip II) - deficit de glicozidaza lizozomala.
Maladia Forbes (tip III) - deficit de enzima de deramificare.
Maladia Andersen (tip IV) - deficit de enzima de ramificare.
Reprezinta procesul de biosinteza a glucozei din compusi neglucidici (sinteza de novo a glucozei). Este o modalitate de obtinere a glucozei cand aportul exogen este intrerupt, iar rezervele de glicogen epuizate.
Procesul este intensificat in inanitie, regim alimentar bogat in lipide si proteine si sarac in glucide, efort prelungit, diabet zaharat si partial, in perioade interprandiale peste 7-8 ore.
Localizare tisulara si intracelulara. Procesul se desfasoara in ficat, rinichi, intestin subtire pentru ca numai aceste tesuturi contin enzima glucozo-6-fosfataza, necesara pentru formarea glucozei. Enzimele sunt localizate si in mitocondrie si in citoplasma.
Principalii compusi glucoformatori sunt: piruvatul, lactatul, aminoacizii glucoformatori, glicerolul.
Se poate spune ca gluconeogeneza decurge oarecum prin parcurgerea in sens invers a etapelor glicolizei, dar cu sinteza de glucoza si consum de ATP. Se ocolesc etapele ireversibile din glicoliza (cele catalizate de piruvatkinaza, fosfofructokinaza si hexokinaza).
Gluconeogeneza din piruvat presupune desfasurarea urmatoarelor etape:
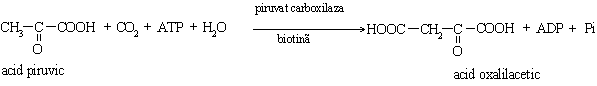
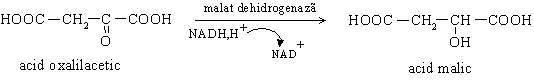
1. Etapa intramitocondriala in care se realizeaza carboxilarea piruvatului cu formare de oxalilacetat. Este o reactie catalizata de piruvat carboxilaza biotin dependenta. Oxalilacetatul nu poate strabate membrana mitocondrialai, de aceea se transforma in L-malat, sub actiunea malat dehidrogenazei mitocondriale. Malatul iese din mitocondrie cu ajutorul unui transportor membranar si este reconvertit la oxalilacetat de catre malat dehidrogenaza citoplasmatica.
2. Etapa citoplasmatica incepe cu o reactie specifica, conversia oxalilacetatului la fosfoenolpiruvat, in prezenta fosfoenolpiruvat carboxikinazei:

Fosfoenolpiruvatul initiaza reactiile de transformare inverse celor din glicoliza pana la fructozo-1,6-difosfat (vezi figura) ce va fo transformat prin etapa catalizata de fructozo-1,6-bisfosfataza:
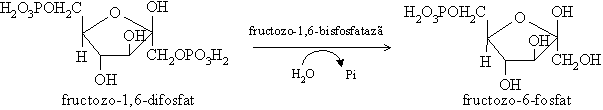
Fructozo-6-fosfatul se transforma in glucozo-6-fosfat invers glicolizei si apoi acesta se transforma in etapa catalizata de glucozo-6-fosfataza. Aceasta are loc in reticulul endoplasmic. Se formeaza vezicule membranare ce difuzeaza spre membrana plasmatica, fuzioneaza cu aceasta si elibereaza glucoza in sange.

Ecuatia globala:
2Piruvat + 4ATP + 2GTP + 2NADH,H+ + 6H2O à Glucoza + 2NAD+ + 4ADP + 6Pi + 2GDP
Pentru sinteza unui mol de glucoza din piruvat se consuma 6 legaturi macroergice.
Gluconeogeneza din lactat (ciclul Cori) nu are reactii specifice. Lactatul format in muschi, este transportat la ficat si transformat in piruvat sub actiunea lactat dehidrogenazei, iar piruvatul in glucoza prin reactiile mentionate anterior.
Gluconeogeneza din aminoacizi presupune transformarea acestora in intermediari glucidici diferiti.
Alanina, serina, cisteina, treonina si glicina se transforma in piruvat;
Acidul glutamic, acidul aspartic, glutamina, asparagina, triptofanul, prolina, arginina, fenilalanina si tirozina se transforma in oxalilacetat;
Izoleucina, metionina si valina se transforma in succinat.
Gluconeogeneza din glicerol. Glicerolul rezultat din lipoliza din tesutul adipos si ficat este fosforilat si transformat apoi in dihidroxiacetonfosfat, intermediar glicolitic. Gluconeogeneza din glicerol reprezinta 10% din capacitatea ficatului.
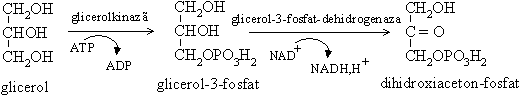
Propionatul rezultat din acizii grasi cu numar impar de atomi de carbon se transforma in oxalilacetat si apoi in glucoza.

Secventa gluconeogenezei inversa glicolizei
Reglarea gluconeogenezei se realizeaza prin:
a) modularea activitatii enzimelor cheie de diferiti efectori;
b) reglare hormonala.
Intensitatea gluconeogenezei este invers proportionala cu a glicolizei.
Efectorii pozitivi ai glicolizei sunt efectori negativi ai gluconeogenezei.
|
Glicoliza |
Gluconeogeneza |
||
|
Efectori pozitiv |
Efectori negativi |
Efectori pozitiv |
Efectori negativi |
|
Fructozo-1,6-bisfosfat |
AcetilCoA |
AcetilCoA |
Fructozo-1,6-bisfosfat |
|
AMP |
AMP |
||
Insulina actioneaza direct la nivelul ficatului inhiband sinteza unor enzime cheie ce actioneaza in gluconeogeneza, stimuland deci glicoliza;
Glucagonul stimuleaza lipoliza in tesutul adipos, dar poate sa si induca enzimele gluconeogenetice determinand activarea unor factori de transcriptie;
Hormonii glucocorticosteroizi secretati de corticosuprarenala stimuleaza proteoliza extrahepatica si induc sinteza enzimelor gluconeogenetice specifice.
Gluconeogeneza este dependenta de existenta substraturilor pentru piruvat carboxilaza si PEP carboxikinaza.
Compusii care limiteaza concentratia substraturilor respective, au efect inhibitor asupra gluconeogenezei.
De exemplu, consumul cronic de etanol la persoanele subnutrite se asociaza cu hipoglicemie deoarece metabolizarea etanolului reduce cantitatea de piruvat, respectiv oxalilacetat necesare gluconeogenezei.
Metabolizarea etanolului se face pe mai multe cai, una dintre acestea fiind oxidarea la acetaldehida sub actiunea alcool dehidrogenazei.
CH3CH2OH + NAD+ = CH3CHO + NADH, H+
Piruvat + NADH, H+ Lactat + NAD+ sau Oxaloacetat + NADH, H+ Malat + NAD+
Reactiile globale sunt:
Etanol + Piruvat Acetaldehida + Lactat, respectiv Oxaloacetat + Etanol Malat + Etanal
Prin aceasta se argumenteaza efectul inhibitor al etanolului asupra gluconeogenezei.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 12619
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved