| CATEGORII DOCUMENTE |
| Alimentatie nutritie | Asistenta sociala | Cosmetica frumusete | Logopedie | Retete culinare | Sport |
Modularea moleculelor de adeziune celulara la suprafata endoteliului activat
I.1. Expresia ICAM-1, VCAM-1 si E-selectina indusa de TNF-a si glucoza crescuta in celulele endoteliale. Adeziunea monocitelor la endoteliul activat. Efectul medicamentelor antiinflamatoare: aspirina, clexanul
a. INTRODUCERE SI OBIECTIVE
Proprietatile adezive ale endoteliului - monostratul de celule al sistemului cardiovascular care intra in contact cu sangele - sunt centrale pentru patologia acestuia. In conditii normale, suprafata luminala a celulelor endoteliale prezinta proprietati relativ neadezive si nontrombogenice pentru constituentii celulari si macromoleculari ai sangelui. Diferitele interactii dintre celulele endoteliale si constituentii sangelui sunt modificate in anumite boli care includ inflamatia, ateroscleroza, angiogeneza patologica si leziunile vasculare. Cele mai multe boli inflamatorii sunt caracterizate de o acumulare anormala de celule circulante (neutrofile, monocite, limfocite T, celule plasmatice, plachete). Atasarea leucocitelor, si migrarea lor ulterioara in vasculatura si in tesut necesita actiunile comune a mai multor molecule de adeziune celulara si a chemokinelor. Adeziunea celulara este una din caile prin care celulele comunica unele cu altele si este un proces complex care implica un numar mare de molecule diferite. Nu toate moleculele de adeziune sunt exprimate constitutiv la suprafata celulara, expresia lor fiind indusa de diferiti activatori, ca citokine, interleukine, stres oxidativ si concentratii crescute de glucoza. Activarea este indusa direct de agonist sau ca o consecinta a unei reactii de semnalizare initiata de legarea unei alte molecule de adeziune la ligandul ei specific. Astfel, adeziunea celulara si reglarea ei este un proces complex. Cele mai importante familii de molecule de adeziune sunt: integrinele, imunoglobulinele, selectinele si caderinele.
Se stia ca mecanismul de aciune al medicamentelor antiinflamatorii nonsteroidale este inhibarea sintezei prostaglandinelor prin blocarea ciclooxigenazei COX-2, insa evidentele experimentale au aratat ca blocarea COX - 2 nu este singurul si poate nici cel mai important mecanism de actiune a acestor medicamente. Aspirina, agent nonsteroidian antiplachetar, inhiba ireversibil si covalent COX inhiband biosintexa tromboxanului plachetar. In ultimii ani, aspirinei i s-au atribuit proprietati antioxidante si abilitatea de a modula transcriptia unor gene imunologice importante. Pentru ca nu existau date in literatura relativ la efectul unor doze terapeutice de aspirina asupra activarii endoteliului, am testat efectul aspirinei asupra moleculelor de adeziune celulara induse de TNF-a si glucoza crescuta.
Heparinele de greutate moleculara mica (LMWHs) reprezinta un grup heterogen de substante folosite ca medicamente anticoagulante si antitrombotice, fiind eficiente in tromboza venoasa si arteriala. Enoxaparina este o heparina de masa moleculara mica (~4300D) obtinuta prin taierea esterului benzil al heparinei cu tratament alcalin. Trialele clinice [Tyrell et al., 1999; Lever et al., 2000] au demonstrat ca in sindromul coronarian acut acest medicament este mai eficient decat alte heparine. S-a aratat ca LMWHs inhiba inflamatia acuta in vivo [Smalbegovic et al., 2001], insa se cunostea foarte putin despre posibilele mecanisme implicate. Astfel, ne-am propus sa evaluam activitatea potential anti-inflamatoare a enoxaparinei prin explorarea efectelor directe ale acesteia asupra moleculelor de adeziune celulara ICAM-1, E-selectina si P-selectina.
S-a aratat ca adeziunea crescuta a monocitelor la celulele endoteliale poate juca un rol important in ateroscleroza accelerata asociata diabetului. Adeziunea monocitelor izolate de la pacientii cu diabet de tip II la CE in cultura este mai mare decat adeziunea monocitelor izolate de la pacientii normali [Carantoni et al., 1997; Kunt et al., 1999]. In plus, s-a demonstrat ca adeziunea monocitelor la CE activate de concentrtii crescute de glucoza (mimand conditiile diabetului) este mai mare decat la CE expuse la concentratie normala de glucoza [Kim et al., 1994; Morigi et al., 1998; Manduteanu et al., 1999]. Printre posibilele mecanisme implicate in acest proces, s-a sugerat ca moleculele de adeziune celulara supra-exprimate de concentratia de glucoza crescuta pot juca un rol semnificativ [Morigi et al., 1998; Manduteanu et al., 1999]. Astfel, in studiile noastre am urmarit efectul aspirinei si clexanului nu numai asupra expresiei moleculelor de adeziune celulara, dar si asupra rolului functional al acestora, adeziunea monocitelor la endoteliul activat de TNF-a sau glucoza crescuta.
b. MATERIALE SI METODE
Celule
CEV- celulele endoteliale valvulare au fost izolate de la valve atrioventriculare bovine si cultivate (conform metodei descrise Manduteanu et al., 1998) si au fost crescute in DMEM continand 20% ser fetal de vitel.
EA 926 este o linie de celule endoteliale vasculare umane permanente obtinuta prin hibridizarea HUVEC cu A549 (celule ce provin de la o carcinoma de plaman, umane). Sunt crescute in DMEM continand 15% ser fetal de vitel.
Celulele au fost subcultivate in placi de cultura cu 96 si 24 de godeuri.
U937 - o linie celulara monocitica (oferita prin amabilitatea Profesorului S.C. Silvestein, Columbia University, New York, USA) crescute in suspensie in mediu de cultura RPMI 1640 continand 5% ser fetal de vitel.
ELISA
Celulele crescute in conditii normale, sau activate si activate in prezenta medicamentelor antiinflamatoare au fost spalate de doua ori cu tampon de spalare (PBS continand 1% albumina din ser bovin) si fixate cu 1% paraformaldehida timp de 20 de minute la temperatura camerei. Fixatorul a fost indepartat cu tampon de spalare, iar celulele au fost incubate cu anticorpi monoclonali anti ICAM-1, VECAM-1, E si P-selectina (5mg/ml) timp de 1h. Dupa o spalare, celulele au fost incubate cu al doilea anticorp IgG de soarece, conjugat cu enzima Horseradish peroxidase (1:1000) timp de 1h la temperatura camerei. A urmat adaugarea subtratului (2mg/ml o-phenylenediamine, 0.03 H2O2 in tampon fosfat 0.1M) pentru reactia peroxidazei. Dupa 30 de minute, reactia a fost stopata prin adaugarea de acid sulfuric 2M, iar absorbtia a fost citita la un cititor ELISA (Multiscan) cu filtru interferential la 492 nm..
Microscopie de fluorescenta
Pentru aceasta tehnica s-a folosit acelasi protocol ca si pentru ELISA cu deosebirea ca celulele au fost crescute pe lamele de cultura iar cel ce-al de-al doilea anticorp folosit a fost cuplat cu fluoroforul FITC care are lunginea de unda de excitatie (lex) la 495nm si emisia, lem, la 519nm. Vizualizarea rezultatelor s-a facut la microscopul de fluorescenta (Nikon Microphot-SA).
Determinarea expresiei genice a moleculelor de adeziune celulara
S-a urmarit sinteza ARNm specific genelor ce codifica molecule de adeziune (ICAM-1, VCAM-1) din celulele endoteliale normale, activate sau activate si tratate cu aspirina.
Mai intai s-a izolat ARN total utilizand kit special de microizolare (Glass Max RNA microisolation - spin cartridge system) bazat pe extractia ARN cu guanidinizotiocianat, mercaptoetanol si precipitare cu alcool etilic. Concentratia de ARN total extras s-a apreciat prin masurarea extinctiei la 260nm. ARN izolat a fost supus tehnicii RT-PCR. S-a utilizat kitul de reverstranscriere si amplificare intr-o singura etapa (one tube RT-PCR system ce contine revers transcriptaza AMV si Taq-ADN polimeraza). In reactia de amplificare s-au folosit urmatorii primeri specifici: pentru ICAM-1 sens :5'-GTCCCCCTCAAAAGTCATCC-3'si antisens: 5'-AACCCCATTCAGCGTCACCT-3', pentru VCAM-1 sens : 5'-ACCACAGGCTGTGAGTCC-3' si antisens: 5'-TGTGTCTCCTGTCTCCGC-3'. Reactia de RT-PCR a fost controlata prin utilizarea de primeri specifici pentru gliceraldehid-fosfatdehidrogenaza (GAPDH), sens: 5'-ACCACAGTCCATGCCATCAC-3'si antisens : 5'-TCCACCACCCTGTTGCTGTA-3'
Produsii rezultati au fost supusi elecroforezei in gel de agaroza 1.5%, iar rezultatele analizate cu un system analizor de geluri (Image Master VDS-Pharmacia Biotech). Nivelul de ARNm MCP-1 a fost normalizat relativ la ARNm GAPDH, iar rezultatele obtinute de la trei experimente independente au fost analizate statistic.
Adeziune celulara
Pentru evaluarea adeziunii monocitelor la CE activate de TNFa si glucoza crescuta si urmarirea efectului aspirinei si clexanului asupra acesteia, CE activate au fost spalate cu mediu de cultura incalzit (37 C) si apoi incubate cu monocite (500 000/ml) suspendate in RPMI cu 1% ser fetal de vitel, timp de 30 minute la 37 C. Au urmat trei spalari cu acelasi mediu de cultura pentru a indeparta celulele neaderate, dupa care monostratul a fost fixat cu 2% paraformaldehida in 0.1 M tampon fosfat, 20 minute. Monostratul fixat a fost spalat de 3 ori cu PBS si examinat in contrast de faza la microscop. Numarul monocitelor aderate a fost cuantificat prin numarare directa a monocitelor in cinci campuri microscopice diferite (pentru fiecare conditie experimentala au existat probe duplicat sau triplicat). Datele morfometrice obtinute au fost exprimate ca numar de monocite aderate pe camp microscopic. Procesarea statistica s-a facut cu Anova din programul Origin.
c. REZULTATE
A. Efectele aspirinei asupra celulelor endoteliale activate de glucoza crescuta si TNFa
1. Celule activate cu concentratie crescuta de glucoza
Celulele endoteliale ombilicale umane (EA 926) au fost crescute in conditii normale de glucoza (5mM, acestea fiind celulele control) sau in conditii de glucoza crescuta (33mM). Pentru a urmari efectul aspirinei asupra expresiei moleculelor de adeziune, la confluenta, EA 926 crescute in 33mM glucoza au fost impartite in doua loturi experimentale: i) celule incubate inca 2 zile in coditii de glucoza crescuta (GC) si ii) celule incubate in 33mM glucoza dar tratate cu aspirina (1mM) timp de 2 zile (GC+A).
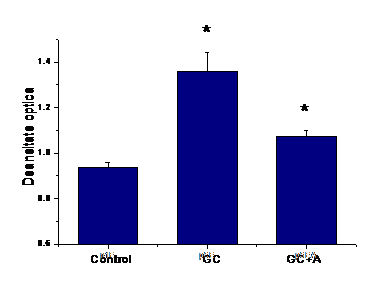
A.1) Efectul aspirinei asupra expresiei de VCAM-1 la suprafata celulelor EA 926 aflate in conditii de glucoza crescuta
Concentratia crescuta de glucoza induce expresia VCAM-1 la suprafata celulelor endoteliale. Tratamentul cu aspirina a redus semnificativ (p=0.04) expresia de VCAM-1 (figura I.1).
Figura I.1. Expresia VCAM-1 la suprafata celulelor endoteliale EA 926 activate de glucoza crescuta.
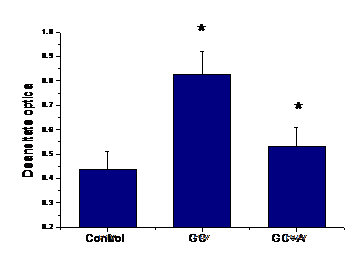
A.2) Efectul aspirinei asupra expresiei de ICAM-1 la suprafata celulelor EA 926 aflate in conditii de glucoza crescuta
Incubarea celulelor endoteliale aflate in 33mM glucoza cu aspirina a redus semnificativ expresia de ICAM-1 (p=0.03, figura I.2)
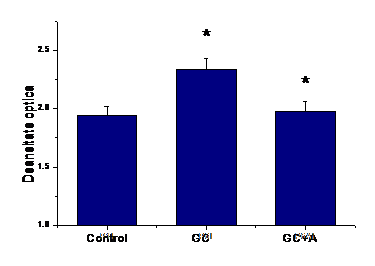
Figura I.2. Expresia VCAM-1 la
suprafata celulelor endoteliale EA 926 activate de glucoza crescuta.
A.3) Efectul aspirinei asupra expresiei de E-selectina la suprafata celulelor EA 926 aflate in conditii de glucoza crescuta
Si in cazul moleculei de adeziune E-selectina, concentratia mare de glucoza induce cresterea expresiei acestei molecule comparativ cu celule crescute in conditii normale de glucoza. Tratarea cu aspirina reduce semnificativ (p=0.03) expresia E-selectinei (figura I.3).
Figura I.3. Epresia E-selectinei
A.4) Efectul aspirinei asupra expresiei genice de ICAM-1 si VCAM-1 in celulele endoteliale umane activate cu glucoza crescuta
Pentru a urmari daca cresterea expresiei proteice de la suprafata celulelor endoteliale activate de glucoza crescuta este datorata unor modificari la nivel de expresie genica, am urmarit efectul glucozei crescute asupra expresiei genice a ICAM-1 si VCAM-1. Rezultatele obtinute prin tehnica RT-PCR arata ca expresia genica a ICAM-1 este indusa de concentratia crescuta de glucoza (33mM) si scazuta de aspirina (A1=1mM, A2=0.5mM) (figura I.4 A). Astfel, cresterea expresiei proteice a ICAM-1 la suprafata membranara a celulelor activate de glucoza se datoreaza unei cresteri a expresiei genei acestei molecule.
Figura I.4. Expresia genica a ICAM-1 (A) si VCAM-1 (B)
![]()

In cazul moleculei de adeziune VCAM-1, expresia genica este slab exprimata in celulele control si crescuta semnificativ in celulele activate de glucoza. Aspirina scade expresia genica a VCAM-1 (figura I.4 B). Astfel, aspirina reduce expresia ICAM-1 si VCAM-1 de la suprafata celulara, actionand la nivel de expresie genica a celor doua molecule de adeziune.
A.5) Adeziunea monocitelor la endoteliul activat cu concentratie crescuta de glucoza
Pentru a testa rolul functional al expresiei moleculelor de adeziune induse de glucoza crescuta, am efectuat experimente de adeziune a monocitelor (U937) la celulele endoteliale activate de glucoza in absenta sau prezenta de aspirina.
Figura I.5. Adeziunea momocitelor la celulele endoteliale
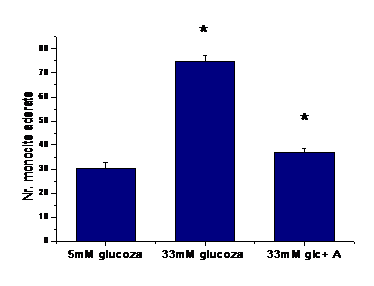
Rezultatele obtinute arata ca numarul monocitelor care adera la celulele endoteliale activate cu 33mM glucoza este crescut semnificativ fata de numarul monocitelor aderate la CE control. De asemenea, concentratia de 1mM aspirina reduce semnificativ adeziunea monocitelor la CE activate de glucoza (figura I.5).
2. Celule activate cu TNFa
Anterior s-a demonstrat ca TNF-a, o citokina puternic pro-inflamatoare, induce supra-expresia moleculelor de adeziune celulara. Elementele noi aduse de studiul nostru sunt efectele aspirinei asupra moleculelor de adeziune induse de TNF-a. Astfel, in dorinta de urmari daca efectul anti-inflamator al aspiriniei obtinut in conditii de activare cu glucoza crescuta se regaseste si in alt sistem de activare al celulelor endoteliale, am urmarit efectul acesteia in CE activate cu TNF-a. In experimentele nostre, celulele endoteliale valvulare (CEV) bovine au fost crescute in DMEM cu 20% ser fetal de vitel iar celulele endoteliale ombilicale umane (EA 926) au fost crescute in DMEM cu 10% ser. La confluenta, CEV au fost activate cu TNFa (50ng/ml) timp de 6 ore sau 4 ore in prezenta sau absenta de aspirina. In cazul celulelor EA 926, concentratia de TNFa necesara pentru inducerea moleculelor de adeziune la suprafata celulelor a fost de 1ng/ml.
A.6) Efectul aspirinei asupra expresiei de VCAM-1 pe CEV stimulate cu TNF-a
Dupa perioada de incubare cu TNF-a timp de 4ore sau 6 ore nu s-a produs detasarea celulelor de pe vasul de cultura, nu au aparut modificari morfologice ale celulelor endoteliale si nu a fost evidentiata o mortalitate crescuta. In concluzie, citokina (TNF-a) nu a fost toxica pentru celule.
In urma stimularii CEV cu 50ng/ml TNF-a timp de 6 ore expresia de VCAM-1 a fost crescuta (figura I.6), dar redusa semnificativ de ambele concentratii de aspirina (0.5mM si 1mM).
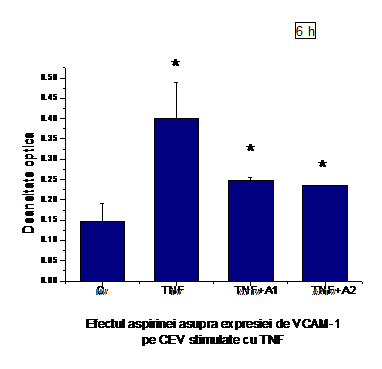
Figura I.6. Expresia VCAM-1 in CEV activate cu TNFα si tratate cu aspirina
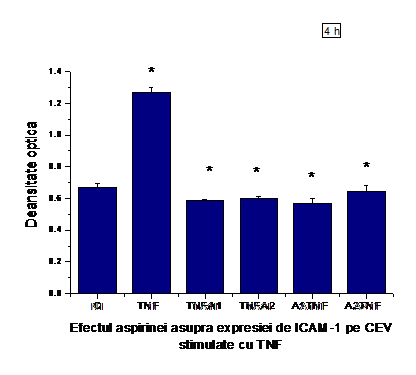
A.7) Efectul aspirinei asupra expresiei de ICAM-1 pe CEV stimulate cu TNF-a
In figura I.7 se observa ca expresia moleculei de adeziune ICAM-1 a fost redusa cu aproximativ 50% de ambele concentratii de aspirina (A1=1M, A2=0.5mM) pe CEV stimulate timp de 6 ore cu TNF-a (50ng/ml). In cazul probelor A1TI si A2TI s-a facut o preincubare de 30 minute cu aspirina, urmata de activarea cu TNF-a
Figura I.7. Expresia ICAM-1 la suprafata CEV activate cu TNFα
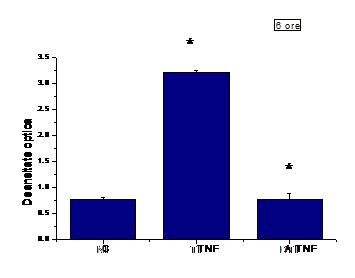
A.8) Efectul aspirineii asupra expresiei de ICAM-1 in celulele EA 926 stimulate cu TNF-a
De asemenea, expresia de ICAM-1 a fost readusa la valoarea bazala de catre aspirina (figura I.8)si in EA 926. Aspirina a fost folosita la concentratia de 1mM pe EA 926 stimulate timp de 6 ore cu TNF-a. S-a facut o preincubare de 30 minute cu aspirina, urmata de activarea cu TNF-a
Figura I.8. Expresia ICAM-1 la suprafata celulelor endoteliale EA 926 activate cu TNFα
Rezultatele obtinute prin tehnica ELISA au fost confirmate de rezultatele de imunofluorescenta.
A.9) Expresia de ICAM-1 la suprafata celulelor EA 926 activate cu TNF-a (1ng/ml) observata la microscopul de fluorescenta.
S-a urmarit expresia de ICAM-1 la suprafata celulelor activate si activate in prezenta de aspirina. Dupa cum putem observa celulele control prezinta foarte slab expresie de ICAM-1 la suprafata. In cazul celulelor activate cu TNFa (1ng/ml) expresia de ICAM-1 este crescuta semnificativ. Aspirina reduce expresia de ICAM-1 indusa de TNFa
Figura I.9. Imunofluorescenta moleculei de adeziune ICAM-1 la suprafata EA 926
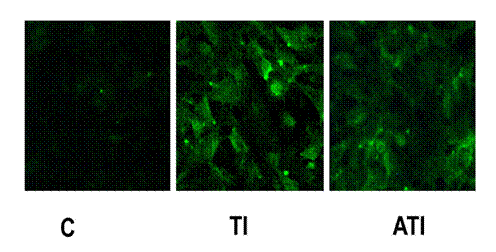
Imagine obtinuta la microscopul de fluorescenta a celulelor EA 926 care au ce-l de-al doilea anticorp cuplat cu FITC, cu lungimile de unda de excitatie, lex=495nm si emisie, lem=519nm.
B. Efectele clexanului asupra celulelor endoteliale activate de glucoza crescuta
B.1) Efectele enoxaparinei asupra moleculelor de adeziune celulara induse de concentratia crescuta de glucoza
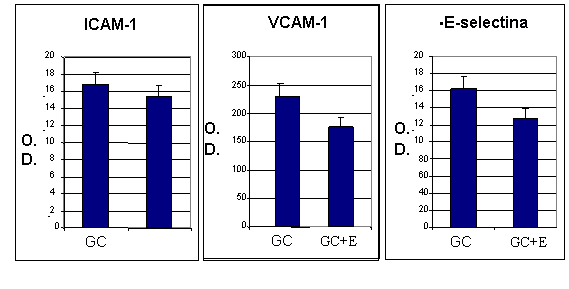
Pentru a elucida efectul enoxaparinei asupra expresiei moleculelor de adeziune celulara, celulele endoteliale (CEV sau EA 926) au fost crescute in conditii normale de glucoza (Control) sau in conditii de glucoza crescuta (GC). La confluenta, celulele crescute in GC au fost expuse in continuare timp de 48 ore la GC in prezenta sau absenta de enoxaparina (16mg/ml). Testele ELISA au aratat ca in urma expunerii CEV sau EA 926 la glucoza crescuta, acestea prezinta o expresie crescuta a moleculelor de adeziune celulara: ICAM-1, VCAM-1 si E-selectina. Dupa cum se observa in figura I.10, in cazul CEV, considerand controlul 100%, cresterea a fost de 168 8% pentru ICAM-1, ~229 10% pentru VCAM-1, si ~162 10% pentru E-Selectina. De asemenea, EA 926 cultivate in conditii crescute de glucoza prezinta cresteri de: 174 8% pentru ICAM-1, 138 10% pentru VCAM-1 si 186 8% pentru E-selectina (figura I.11). Rezultatele arata ca activarea cu glucoza produce cresteri similare ale expresiei de ICAM-1 si E-selectina in ambele tipuri de celule endoteliale, comparativ cu controlul, in timp ce expresia VCAM-1 este mai mare in celulele endoteliale valvulare decat in cele ombilicale. Enoxaparina reduce semnificativ expresia moleculelor de adeziune in ambele tipuri de celule endoteliale. Astfel, expresia ICAM-1 a fost redusa cu 10%, a E-selectinei cu 21% si a VCAM-1 cu 23% in VEC (figura I. 10) iar in cazul EA 926, ICAM-1 cu11%, E-Selectina cu 10% si VCAM-1 cu 10%, (figura I.11)
Aceste date arata ca enoxaparina are efecte similare asupra expresiei de suprafata a ICAM-1 in ambele tipuri celulare, avand un efect mai mare asupra expresiei de E-selectina si VCAM-1 in CEV.
Figura I.10 Efectul enoxaparinei (E) asupra celulelor de adeziune celulara in CEV
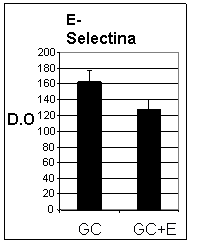
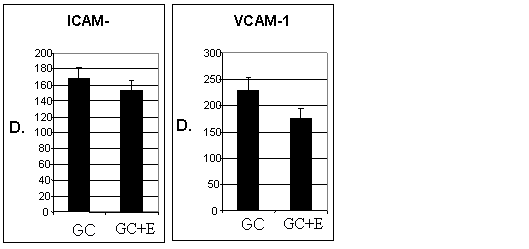
Figura I. 11 Efectul enoxaparinei (E) asupra moleculelor de adeziune in EA 926
B.2) Efectul enoxaparinei asupra adeziunii monocitelor la CE activate de glucoza
Studiile anterioare au artat ca interactia crescuta dintre monocite si celulele endoteliale poate juca un rol important in accelerarea procesului de ateroscleroza asociat cu diabetul. S-a aratat ca adeziunea monocitelor izolate de la pacientii cu diabet de tip 2 la CE in cultura este mai mare decat a monocitelor izolate de la subiecti normali [Carantoni et al., 1997; Kunt et al., 1999]. Astfel, in experimentele noastre am studiat efectul enoxaparinei asupra adeziunii monocitelor la CE activate de glucoza. In acest scop, CE aflate in diferite conditii experimentale au fost incubate cu monocite.
Figura I. 12. Efectul enoxaparinei asupra numarului de monocite aderate la CEV
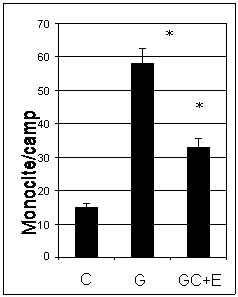
Evaluarea morfometrica a monocitelor arata ca numarul de monocite aderate la CEV crescute in mediu cu concentratie mare de glucoza este de patru ori mai mare decat al celor aderate la celulele control. Tratarea celulelor cu enoxaparina duce la diminuarea semnificativa a adeziunii monocitelor (~ 45%) (figura I.12)
In cazul adeziunii monocitelor la EA 926, numarul monocitelor aderate la celulele crescute in 33mM glucoza a fost de 3 ori mai mare comparativ cu cele crescute in 5mM glucoza (Fig I.13).
Figura I.13 Efectul enoxaparinei asupra numarului de monocite aderate la EA 926
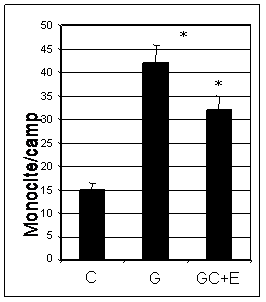
Dat fiind ca testele de adeziune au fost facute in conditii similare pentru ambele tipuri de celule endoteliale (CEV SI EA 926), putem presupune ca in urma activarii cu glucoza celulele originare din valve de inima (CEV) au dezivitate mai mare decat CE ombilicale. De asemenea, noi am gasit ca expresia VCAM-1 este mai mare in CEV decat in EA 926 ceea ce demonstreaza pe de-o parte rolul major pe care il are VCAM-1 in procesul de adeziune al monocitelor la endoteliul activat, si pe de alta parte sustine ideea ca CEV sunt mai adezive decat CE ombilicale (EA 926). De asemenea, rezultatele arata ca tratamentul cu enoxaparina (16mg/ml) timp de 48 ore reduce adeziunea monocitelor la ambele tipuri de celule endoteliale (43% in cazul CEV, figura I.12 si 23% in cazul EA 926, figura I.13).
d. DISCUTII SI CONCLUZII
Studiile anterioare au aratat ca interactiile crescute dintre celulele endoteliale cu monocitele joaca un rol important in ateroscleroza accelerata asociata diabetului. Grupul nostru (dar si altii) a demonstrat ca adeziunea monocitelor este mai mare la celulele endoteliale crescute in concetntratii mari de glucoza, decat la celulele endoteliale crescute in conditii normale de glucoza [Kim et al., 1994; Morigi et al., 1998; Manduteanu et al., 1999]. Printre posibilele mecanisme implicate, un rol important il poate avea expresia crescuta a moleculelor de adeziune celulara.
Rezultatele noastre demonstreaza ca TNFα si nivelele crescute de glucoza asociate diabetului induc expresia moleculelor de adeziune celulara care au un rol crucial in atragerea leucocitelor in situsurile lezate ale endoteliuli, in procesul inflamator. Astfel, ca urmare a inducerii acestor molecule, procesul de adeziune al monocitelor la endoteliul activat este amplificat.
Aspirina si enoxaparina sunt doua medicamente a caror eficienta in bolile cardiovasculare a fost dovedita, datorita efectului lor antiplachetar, respectiv anti-coagulant si antitrombotic. Studii recente au aratat ca aceste medicamente au proprietati noi, printre care anti-inflamatoare. In experimentele noastre aspirina si enoxaparina sunt implicate in reducerea expresiei moleculelor de adeziune celulara de la suprafata celulelor endoteliale crescute in conditii patologice de glucoza. Dat fiind ca aspirina reduce expresia moleculelor de adeziune atat in conditii de activare a CE cu glucoza crescuta dar si cu TNFa, putem presupune ca aspirina intervine in calea de sinteza a acestor molecule, indiferent de agonistul care le stimuleaza expresia. Deosebit de important este faptul ca aceste rezultate au fost obtinute cu concentratii terapeutice de aspirina si enoxaparina care nu sunt toxice subiectilor umani. De asemenea, concluzia care se desprinde din acest capitol este ca exista cai suplimentare de actiune ale unor medicamente cunoscute (in cazul nostru aspirina si enoxaparina), care sunt raspunzatoare de efectul antiinflamator al acestora.
Rezultatele obtinute ajuta la elucidarea bazelor moleculare responsabile pentru efectele medicamentelor antiinflamatorii asupra receptorilor de adeziune, pot ajuta la intelegerea mecanismelor implicate in obtinerea acestor efecte si pot astfel servi ca punct de plecare pentru dezvoltarea unor noi agenti antiinflamatori, mai specifici, folositori in terapia medicamentoasa a bolilor inflamatoare ale sistemului cardiovascular.
I.2. Mecanisme de actiune ale aspirinei si enoxaparinei implicate in reducerea expresiei moleculelor de adeziune
In prima parte a acestui capitol am aratat efecte mai putin cunoscute ale unor medicamente clasice ca aspirina si enoxaparina, si anume de a reduce adeziunea monocitelor la endoteliul activat, prin reducerea expresiei moleculelor de adeziune. In continuare ne propunem sa evidentiem mecanismele de actiune prin care aspirina si enoxaparina intervin in procesul inflamator, reducand expresia moleculelor de adeziune.
In urma diverselor studii care au pus in evidenta mecanisme noi de actiune ale medicamentelor antiinflamatorii nonsteroidale, diferite de calea clasica acceptata, de inhibare a sintezei de prostaglandine, se discuta de efectele medicamentelor antiinflamatorii asupra caii de adeziune a leucocitelor. Aceste studii pot fi de mare ajutor in descoperirea unor noi agenti antiinflamatori care sa blocheze selectiv moleculele de adeziune celulara.
Studiile au debutat prin identificarea tintelor de semnalizare intracelulara, incluzand factorii de transcriptie care controleaza genele inflamatoarei. Exista trei familii de factori de transcriptie importanti care par sa fie reglatori critici ai genelor moleculelor asociate cu bolile inflamatorii (ca citokine, factori de crestere, molecule de adeziune) si anume: activator protein 1 (AP-1)/ATF2, nuclear factor of activated T cells (NFAT) si nuclear factor B (NF-kB). Activitatea acestor factori de transcriptie este reglata direct sau indirect de caile protein-kinazelor activate de factori mitogeni (MAPK).
O caracteristica a MAPK este ca activarea lor completa necesita fosforilarea atat a unei treonine cat si a unei tirozine, care, in proteina sunt separate de un singur aminoacid. Astfel se asigura mentinerea in stare intacta a MAP-kinazelor atat cat ele nu sunt activate specific de o MAPKK.
NF-kB prezinta un interes deosebit in comunitatea medicala datorita rolului pe care il joaca in expresia multor gene ca: TNF-a, IL-1, IL-6, molecule de adeziune, care sunt importante in raspunsul inflamator [Satunton et al., 1990; Smith et al., 1989]. El exista in citoplasma intr-o forma inactiva, asociat cu o proteina inhibitoare IkB. Diversi stimuli extracelulari incluzand TNF-a, IL-1 initiaza o cascada de semnalizare care duce la activarea a doua kinase IkB, IKK-1si IKK-2, care fosforileaza IkB specific la reziduurile serinice amino-terminale [Ozes et al., 1999; Zhang et al., 1999]. Aceasta fosforilare are ca efect translocarea NF-kB in nucleu unde se leaga la gena tinta si initiaza transcriptia diferitelor gene proinflamatoare.
Studiile care incearca sa descifreze mecanisme noi de actiune ale medicamentelor nonsteroidiene au pus in evidenta ca sodiul salicilat si aspirina exercita unele din efectele lor asupra celulelor Jurkat T umane si liniei celulare pre-B din soareci, prin inhibarea factorului de transcriptie nuclear NF-kB, implicat in transcriptia genelor unor citokine pro-inflamatorii si molecule de adeziune endoteliala [Cianferoni et al., 2001]. Pe de alta parte, enoxaparina este acceptata ca un medicament anticoagulant si antitrombotic si mai putin ca avand rol in inflamatia acuta. Totusi, exista indicii ca heparinele de masa moleculara joasa inhiba inflamatia acuta in vivo [Nelson et al., 1993], dar se cunoaste foarte putin despre posibilele mecanisme implicate. Intr-un studiu recent, [Thourani et al., 2000] s-a aratat ca heparina inhiba translocarea NF-kB indusa de TNF-a, si s-a sugerat ca acest efect este datorat legarii medicamentului la secventa de localizare nucleara a NF-kB. Astfel, putem presupune ca enoxaparina poate avea un efect similar asupra translocarii (activarii) NF-kB.
In ultimii ani au aparut dovezi care arata ca expunerea celulelor endoteliale la glucoza crescuta induce cresterea activitatii NF-kB si ca inhibitori specifici ai factorului de transcriptie NF-kB reduc semnificativ adeziunea monocitelor indusa de glucoza; aceste date sugereaza ca glucoza crescuta induce cresterea moleculelor de adeziune prin activarea NF-kB [Morigi et al., 1998]. De asemenea s-a aratat ca concentratiile crescute de glucoza induc disfunctia celulei endoteliale prin generarea speciilor reactive de oxigen [Kashiwagi et al.,1996) Morigi et al., 1998], care, la randul lor induc activarea NF-kB [Du et al., 1999; Kashiwagi et al., 1996]. Astfel, in incercarea noastra de a gasi mecanismele de actiune ale aspirinei si enoxaparinei asupra inducerii moleculelor de adeziune in conditii de glucoza crescuta, am urmarit efectul acestor medicamente asupra speciilor reactive de oxigen si asupra activarii factorului de transcriptie NF-kB, in conditii de glucoza crescuta.
b. MATERIALE SI METODE
Celule
Au fost folosite linia de celule endoteliale (CE) umane EA 926 descrisa in prima parte a acestui capitol (I.1).
Western Blot
Pentru a studia efectul aspirinei si enoxaparinei asupra factorului de transcriptie NF-kB, CE au fost crescute in DMEM continand : (i) concentratie normala de glucoza, 5mM (control), (ii) concentratie crescuta de glucoza, 33mM (GC) si (iii) GC urmata de tratarea timp de 48 ore cu aspirina (1mM) si enoxaparina (16mg/ml). Dupa incubare, celulele au fost lizate in tampon de liza ce contine : 10mM Hepes pH 7.6, 15mM KCL, 2mM MgCl2, 0.1 mM dithiothreitol-DTT, 1% Triton X 100, 0.5 mM PMSF si 5 mg/ml inhibitor1 (benzamidina, pepstatina, leupeptina). Fractiile nucleara si citosolica au fost separate prin centrifugare (650g, 10 min), iar concentratia de proteina din fiecare fractie determinate prin Amido. Proteina din fractia nucleara (80mg) a fost separate in gel de poliacrilamida 12% si transferata pe membrana de nitroceluloza. A urmat blocare timp de 2 ore in tampon cu 5 % lapte degresat si incubarea peste noapte (4 C) cu primul anticorp (2mg/ml anti-subunitatea p65 a NF-kB). Dupa spalari a urmat incubarea timp de o ora cu al doilea anticorp (diluat 1:10 000, la temperature camerei). Detectia benzilor rezultate s-a facut ce kitul ECL (expunere 5 minute) si apoi prin expunerea membranei la film de raze-X. Filmele au fost analizate cu un system vidoe (Image Master from Pharmacia) iar densitatea optica a benzilor a fost determinate cu softul Total Lab 1.11 de la Pharmacia.
Determinarea speciilor reactive de oxygen (ROS)
Acumularea intracelulara de ROS a fost evaluate prin fluorimetrie, cu ajutorul 2',7'-diclorofluoresceinei diacetata printr-o metoda descrisa anterior de [Hempel et al., 1999; Zang et al., 2000]. Colorantul (nefluorescent) (DCFH-DA) este permeabil in celula endodeliala; in interiorul celulei colorantul este hidrolizat la 2',7'-dichlorofluoresceina (DCF), si in urma interactiei cu ROS devine fluorescent. Astfel, celulele aflate in conditii experimentale diferite (control, GC, GC tratate cu aspirina sau enoxaparina) au fost tripsinizate, spalate si incarcate cu 10 mmol/L DCFH-DA in HBSS timp de 45 minutes la 37 C. Dupa doua spalari in tampon HBSS, fluorescenta a fost masurata la spectofotometru Shimadzu RF 5001 PC la lungimea de unda de emisie de 535 nm si excitatie de 505nm. Nivelul ROS a fost exprimat ca unitati de fluorescenta per miligram proteina SE.
Analiza statistica
Procesarea statistica s-a facut cu programul Anova din Origin si testul Student's. Rezultatele au fost considerate semnificative daca valoarea P a fost mai mica decat 0.05.
c. REZULTATE
1) Efectul medicamentelor asupra speciilor reactive de oxigen induse de concentratia crescuta de glucoza
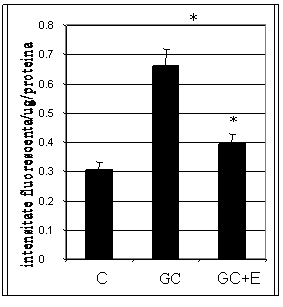
Pentru determinarea mecanismului de actiune al aspirinei si enoxaparinei asupra moleculelor de adeziune induse de hiperglicemie, mai intai am urmarit efectul acestora asupra speciilor reactive de oxigen (ROS) induse de glucoza crescuta. Rezultatele obtinute arata ca CE crescute in conditii de glucoza crescuta prezinta acumulare semnificativa de ROS comparativ ce celulele control (Figura I.14 si I.15). Tratarea celulelor crescute in 33mM glucoza cu aspirina si enoxaparina (48 ore) reduce nivelul ROS la nivelul celulelor control (figurile I.14, respectiv figura I.15).
Figura I.14. Efectul aspirinei asupra speciilor reactive de oxigen

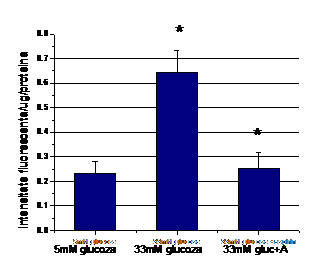
Figura I.15. Efectul enoxaparinei asupra speciilor reactive de oxigen
2) Efectul medicamentelor asupra activarii NF-kB indusa de concentratia crescuta de glucoza
Asa cum am amintit mai sus, NF-kB este mentinut in forma inactiva in citoplasma celulara de protein kinaza IkB. Activarea acestui factor de transcriptie datorita fosforilarii kinazei IkB are ca urmare translocarea subunitatilor NF-kB in nucleu si legarea la genele tinta pe care le regleaza. Astfel, in studiile noastre, pentru a vedea daca NF-kB este implicat in expresia crescuta a moleculelor de adeziune, am urmarit expresia acestuia in fractia nucleara a celulelor aflate in diferite conditii experimentale. Rezultatele de Western Blot arata ca expresia subunitatii p65 in fractia nucleara este crescuta semnificativ de concentratia crescuta de glucoza si ca atat aspirina (figura I.16) cat si enoxaparina (figura I.17) reduc semnificativ aceasta expresie.
Figura I.16. Aspirina reduce epresia nucleara a subunitatii p65 a NF-kB

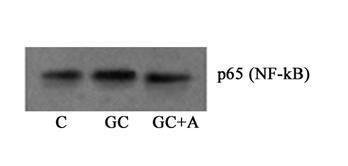
Figura I.17. Enoxaparina reduce epresia nucleara a subunitatii p65 a NF-kB

p65 (NF-kB)

C GC GC+E
d. DISCUTII SI CONCLUZII
Nivelele crescute de glucoza duc la cresterea speciilor reactive de oxigen care, in ultima vreme s-au dovedit a fi molecule semnalizatoare cu un rol deosebit de important in activarea diferitelor cai de semnalizare si factorilor de transcriptie si in expresia moleculelor pro-inflamtoare. NF-kB este un factor de transcriptie sensibil redox care regleaza o multime de gene inflamatoare. Exista studii care au aratat ca expunerea celulelor endoteliale la o concentratie crescuta de glucoza induce cresterea activitatii NF-kB, si ca inhibitori specifici ai acestuia reduc adeziunea crescuta a monocitelor la aceste celule. Si in experimentele noastre, NF-kB este activat de glucoza crescuta, sugerand ca expresia crescuta a moleculelor de adeziune are loc prin intermediul acestui factor de transcriptie. Mai mult, speciile reactive de oxigen supra-exprimate de nivelul crescut de glucoza activeaza acest factor de transcriptie, care in plus duce la expresia crescuta a moleculelor de adeziune celulara ICAM-1, VCAM-1 si E-selectina.
Aspirina si enoxaparina reduc nivelele crescute ale speciilor reactive de oxigen, ceea ce are ca efect scaderea activarii factorului de transcriptie NF-kB a carui implicare in expresia moleculelor de adeziune este larg studiata si bine stabilita. Un alt posibil mecanism prin care aspirina scade expresia nucleara a NF-kB este prin prevenirea degradarii inhibitorului NF-kB, anume IkB, si astfel imposibila translocare, deci activare a acestui factor de transcriptie [Kopp & Ghosh, 1994]. In cazul enoxaparinei, mecanismul prin care aceasta inhiba activarea NF-kB este mai putin clar. Din experimentele noastre putem presupune ca reducerea nivelului speciilor reactive de oxigen este un posibil mecanism prin care enoxaparina afecteaza activarea NF-kB. Astfel, cele doua medicamente au efecte benefice asupra celulelor endoteliale activate de concentratie mare de glucoza reducand expresia moleculelor de adeziune celulara -ICAM-1, VCAM-1 si E-selectina- printr-un mecanism ce implica scaderea expresiei nucleare a factorului de transcriptie NF-kB.
Descifrarea mecanismelor de actiune ale medicamentele antiinflamatorii, medicamente care pot interfera cu adeziunea celulelor circulante la celulele endoteliale prin inhibarea unuia sau mai multor pasi de aderare, poate reprezenta un pas important in terapia impotriva efectelor distrugatoare ale inflamatiei cronice necontrolate.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1795
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2024 . All rights reserved