| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Bacteriofagii
Bacteriofagii sunt virusuri care infecteaza celulele bacteriene (eubacterii). Cel mai adesea, rezultatul interactiunii este liza celulei gazda. Fenomenul lizei transmisibile a bacteriilor a fost descoperit de Twort (l9l5) si l-a atribuit unei enzime litice sau unui virus. D'Herelle (l9l7) a descoperit independent fenomenul lizei bacteriene, a demonstrat natura particulata a agentului inductor si l-a denumit bacteriofag ("mancator de bacterii") sau prescurtat, fag. Denumirile fagilor deriva de la grupul de organisme gazda: actinofagi (infecteaza actinomicetele), cianofagi (pentru cianobacterii), micofagi (infecteaza fungii microscopici).

Fig. 95. Principalele tipuri morfologice de bacteriofagi.
Se disting trei tipuri morfologice de fagi (fig. 95):
fagii in forma de cireasa cu coada: au cap si coada de lungimi diferite, flexibila sau rigida);
fagii filamentosi
fagii sferici, fara coada.
Structura moleculara este mai bine cunoscuta pentru fagii din seria T, in special din grupul T-par (T2, T4, T6) si fagul l, (infectiosi pentru E. coli), fagul φ X l74.
Anatomia moleculara a fagilor din seria T-par
Studiul fagilor la nivel molecular a fost initiat in anii '40 de catre M. Delbruck. Particula fagica matura din seria T-par, are gr. mol. de 2,2 x l08 D si este alcatuita din ADN (40%)si proteine (60%).
Simetria fagului T4 este binara: capsomerele regiunii capului sunt asezate dupa o simetrie icozaedrica, iar la nivelul cozii, dupa o simetrie helicala.
Capul fagului T4, cu o lungime de l00 nm si latimea de 65 nm, pe imaginile electrono-optice ale preparatelor umbrite, are forma poliedrica*, iar in sectiune longitudinala, are un contur hexagonal (fig. 96).
*Forma geometrica exacta a capului nu este cunoscuta, deoarece capsomerele nu se observa in structura sa intacta, ci numai dupa dezintegrarea partiala. Pe baza formei umbrelor virionilor in preparatele umbrite, capul este descris ca o prisma hexagonala bipiramidala. Cele doua piramide sunt decalate una fata de alta cu un unghi de 30o. Din cauza decalajului, ar rezulta o configuratie geometrica, denumita antiprisma hexagonala bipiramidala.
In interiorul capului se gaseste genomul fagic, o molecula de ADN dublu catenara, lineara, cu o lungime de circa 50 m si aproximativ 200 de gene, "impachetata" foarte strans, asociata cu alte tipuri de molecule:
o proteina interna (300 - l000 de molecule), cu gr. mol. mica;
un polipeptid acid (l000 - 3000 de molecule);
doua tipuri de poliamine (spermidina si putrescina).
La varful uneia dintre piramide se gaseste un mic "dop proteic", care face legatura intre capsida (cu axe de simetrie de tip 5 si 2) si gat, un tub cilindric, cu simetrie helicala. Dopul proteic ar avea rolul de "adaptor de smetrie".
Gulerul are forma unei placi hexagonale, cu un orificiu in zona centrala, strabatut de cilindrul axial al cozii, situat in continuarea gatului.
Cilindrul axial al cozii are un diametru de 7,5 nm si un lumen canalicular de 2 nm. Prin acest canal, ADN fagic este transferat in celula bacteriana.
In jurul cilindrului axial este asamblata teaca contractila a cozii. Cilindrul axial este mai lung decat teaca cozii si proemina in raport cu ea, avand rolul de a perfora peretele celulei bacteriene. Teaca cozii este alcatuita din l44 de capsomere, dispuse pe 24 de randuri, dupa o simetrie helicala. Prin contractie, teaca se scurteaza, datorita reasezarii spatiale a capsomerelor si elibereaza cilindrul axial pe o lungime de circa 50 nm, permitandu-i sa perforeze peretele celulei bacteriene.
La extremitatea cozii se gaseste placa bazala, cu aspectul uni disc hexagonal, cu diametrul de 40 nm, formata dintr-un pivot central si 6 subunitati asezate dupa o simetrie radiala de tip 6. Orificiul central al pivotului este strabatut de cilindrul axial al cozii. Placa este prevazuta cu 6 carlige, denumite crosetele cozii, cate unul la fiecare varf al hexagonului. Rolul crosetelor este de a fixa ferm particula fagica pe suprafata celulei bacteriene.
Fibrele cozii, in numar de 6, sunt structuri filamentoase proteice, lungi de l30 nm si par a fi alcatuite din 4 subunitati identice. Cu una dintre extremitati, fiecare fibra este fixata stabil la unul dintre varfurile placii hexagonale, iar cu extremitatea distala se leaga lax de subunitatile gulerului, formand astfel invelisul extern al cozii, in jurul tecii contractile. Inainte de fixarea pe celula, fibrele se desprind din insertia lor distala pe guler si raman legate numai de placa bazala. Fibrele cozii ar avea rolul unor structuri de ancorare a fagului de peretele bacterian, in cursul interactiunii primare.
Genomul fagic. Fagii cei mai mici - fagi ARN "masculi" - (Q-beta, MS2 si f2) au genomul format dintr-o molecula de ARN monocatenar, de circa 3,5 x l03 baze, cu topologie lineara. Adeseori, molecula de ARN formeaza structuri secundare dublu catenare in ''ac de par''(hairpin), prin intermediul puntilor de H care reunesc 60-80% din totalul bazelor.
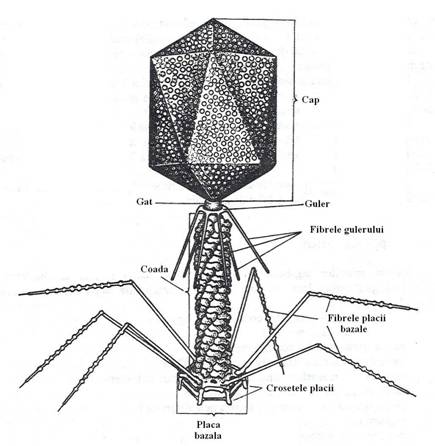
Fig. 96. Anatomia bacteriofgului T4.
Fagii filamentosi (Ml3, f1, fd) si cei cu simetrie icozaedrica, fara coada ( X l74 etc.), au genomul format dintr-o molecula de ADN monocatenara, circulara.
Fagii din seria T-par (T2, T4, T6), T-impar (T1, T3, T5, T7), fagul , P2 au genomul format dintr-o molecula de ADN dublu catenara, lineara.
Exista deosebiri ale secventelor terminale ale moleculei de ADN. Astfel, la fagii , P2 (cu genom ADN dublu catenar, linear), ambele extremitati se termina cu secvente monocatenare, complementare unele fata de altele, formand capete aderente, coezive sau "lipicioase". Prin legarea celor doua extremitati, pe baza complementaritatii, genomul fagului, liber in celula gazda, se circularizeaza.
Fagii din seria T-impar au ca genom, o molecula de ADN dublu catenara lineara, iar la extremitati, au secvente nucleotidice repetate invers, de la 250 la l0000 de baze.
Genomul fagului T4 contine circa 200 de gene. Peste 50 dintre ele au rol in procesul asamblarii, iar celelalte codifica diferite proteine cu rol in ciclul de multiplicare. O mica parte dintre genele de multiplicare sunt esentiale, iar restul sunt neesentiale, deoarece au corespondent printre genele cromosomului bacterian. Fenomenele mutationale ale genelor neesentiale nu blocheaza desfasurarea ciclului de multiplicare, dar randamentul infectiei este net inferior (se asambleaza un numar mai mic de fagi progeni).
Informatia genetica a fagilor, ca si a celulelor bacteriene are caracter continuu, adica, de regula lipsesc secventele necodificatoare. Cateva gene ale fagului T4 contin cate un intron de circa l000 pb (de exemplu, gena timidilat-kinazei).
Genomul fagic se deosebeste de al virusurilor infectioase pentru celulele animale, prin continutul in baze neobisnuite: 5-hidroxi-metil-citozina, in locul citozinei la fagii din seria T-par sau 5-hidroxi-metil-uracil si 5-4,5-dihidroxi-pentil-uracil.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 2560
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved