| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Condriomul celular
Este format din totalitatea condriozomilor, elemente caracteristice celulei eucariote si absente la procariote.
Condriomul a fost descoperit de BENDA la 1900 dar progrese insemnate in studiul sau au fost inregistrate numai in urma observatiilor efectuate in microscopia cu contrast de faza, ME si dupa ultracentrifugarea si izolarea lui.
De notat faptul ca aceste organite celulare prezinta acelasi aspect atat in celula animala cat si
cea vegetala.
Morfologie. In microscopia optica se deosebesc trei aspecte principale: unele au aspect globular oval si se numesc mitocondrii, altele au aspect alungit, in forma de virgula si se numesc coridrioconte iar altele in forma de granule insirate numite condriomite (Fig. 45).
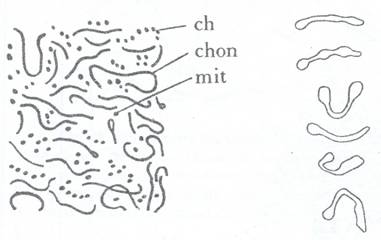
Fig. 45: Forme de condriom: ch- condriomite, choncondrioconte, mit- mitocondrii (stg) si modificarea in vivo a condriocontelor sub actiunea curentilor citoplasmatici.
Dimensiunile lor sunt cuprinse intre 0,3-1,5 μm cele globuloase si 2-30 μm cele alungite. Ele au refringenta foarte scazuta astfel ca in microscopia optica pot fi observate numai in contrast de faza sau prin folosirea unui colorant vital verdele de Janus, care se fixeaza pe flavoproteine si datorita enzimelor oxidative, colorantul nu este redus la leucoderivati, ramanand deci colorat numai la nivelul condriomului. In general, condriomul este sensibil la fixatori si la actiune a apei, aceasta din urma, in mediu hipotonic, produce cavulatie prin care condriozomii se veziculeaza si se leaga in filamente lungi. Ei sunt sensibili si fata de curentii citoplasmatici, sub actiunea carora se curbeaza sau se deformeaza si se ramifica. Ei pot sa fuzioneze si sa se divida.
Ultrastructura lor a fost descifrata numai dupa
1952, prin folosirea ME. In sectiuni ultrafine a fost observata o
ultrastructura foarte
Membrana externa are o grosime de 75 A si suprafata lipsita de ribozomi si fara relatii cu RE. Ea prezinta o structura tipica, tripartita. Membrana interna se repliaza spre interior, trirnitand in matrice prelungiri sub forma de creste aplatizate (tipul 'crista') uneori ramificate sau sinuoase, iar alteori prelungirile au forma de tubuli (tipul 'tubulus'). In matrice au fost puse in evidenta granule dense de 300-400 A, fibrile de ADN, ribozomi (Fig. 47). Prin tehnica 'coloratiei negative' au fost observate pe creste, unitatile structurale pe care FERNANDEZ- MORAN (1964) le-a denumit ,.,particule elementare' ale mitocondriilor sau oxizomi, la nivelul lor avand loc transferul de electroni din lantul oxidativ din mitocondrii. Acestia sunt alcatuiti dintr-un cap sferic (80 A), un corp cilindric (50 A) si o parte bazala (110 A) (Fig. 47).
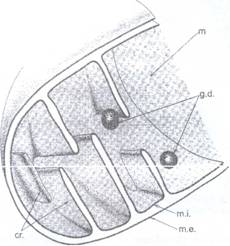
Fig. 46 : Schema ultrastructurii unei
mitocondrii.
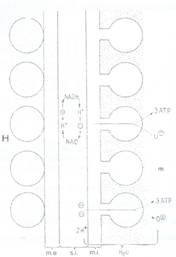
Fig. 47: Schema structurii oxizomilor atasati de membrana interna a mitocondriei. me- membrana externa; H- hialoplasma; si- spatiu intermembranar; mi- membrana interna; m- matrice mitocondriala.

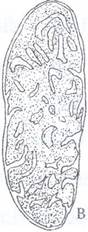
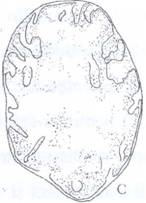
Fig. 48: Evolutia ultrastructurii mitocondriilor in cursul procesului de diferentiere celulara la crizanteme. A- celule meristematice cu creste reduse si matrice putin densa. B- celule la inceputul diferentierii cu numar mare de creste si matrice densa. C- celule in varsta cu creste putine, tubuli si matrice cu densitate foarte redusa.
Celulele meristematice prezinta mitocondrii sarace in creste si bogate in matrice in timp ce in celulele diferentiate creste procesul de lamelizare in defavoarea matricei, iar in celulele in varsta, crestele se rarefiaza si apar tuburi (Fig. 49).
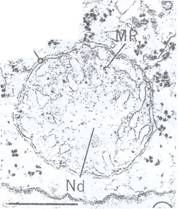
Fig. 49: Mitocondrie din tesutul liberian al racinii de fasole vazuta in microscopia electronica. E- membrana dubla; MR ribozomi mitocondriali; Nd- regiunea nucleoidului cu continut de ADN.
Compozitia chimica. La inceput s-au pus in evidenta in mitocondrii lipide si proteine corespunzatoare structurilor membranare foarte dezvoltate. Dupa izolarea mitocondriilor prin centrifugare s-a aprofundat si studiul lor chimic. A urmat apoi tehnica de fractionare a mitocondriilor care a condus la izolarea membranelor externe, interne, a crestelor si a matricei. In urma cercetarilor chimice efectuate s-a confirmat continuul ridicat de proteine (60-70%) si lipide (25-30%) din substanta uscata, membrana externa fiind de 2-3 ori mai bogata in fosfolipide. Dintre acestea in membrana externa sunt prezente fosfatidilinositolul si colesterolul, iar in membrana interna, cardiolipina (difosfatidilglicerol). Mitocondriile contin o proportie ridicata de ioni de K+ si Mg2+, iar dintre nucleotide se remarca in primul rand cele ale adeninei (ATP, NAD). Mitocondriile sunt bogate si in enzime oxidoreducatoare si se cunoaste cu precizie si localizarea lor pe membrana externa, interna, spatiul intermembranar si in matrice. Astfel, enzime 'marker' sunt considerate a fi monoaminoxidaza (MAO) pentru membrana externa, enzime ale metabolismului ATP in spatiul intermembranar, citocom-C-oxidaza pentru membrana interna si enzimele ciclului Krebs in matrice. In matricea mitocondriala este prezent si ADN, acesta fiind asemanator cu cel bacterian si diferit de cel nuclear prin GM, raportul de baze si lipsa histonelor, avand o forma circulara si poate transcrie cca 25 tipuri diferite de ARNm. Duplicarea ADN-ului mitocondrial se face independent de cel nuclear. Toate acestea pledeaza in favoarea originii simbiotice a mitocondriilor, rezultat al patrunderii in celula eucariota a unui procariot aerob.
Mitocondriile se pot inmulti prin diviziune sau se pot forma 'de novo' pornind de la ultracondriom (Fig. 50).
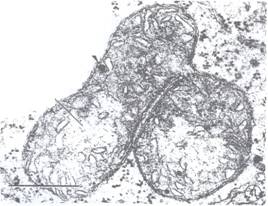
Fig. 50: Imagine electronomicroscopica ce surprinde un proces de diviziune a mitocondriilor.
Formarea crestelor pe membrana mitocondriala interna conduce la cresterea suprafatei acesteia de cca 5 ori in comparatie cu membrana externa si implicit a capacitatii de reactie biochimica a acesteia.
ADN-ul mitocondrial este capabil sa exercite functiile de transcriptie si duplicatie. In mitocondrii au fost identificate toate cele 3 tipuri de ARN necesare pentru o sinteza autonoma de proteine, asemanate insa cu cea din bacterii.
ADN-ul mitocondrial nu poate asigura insa numai cca 30% din proteinele mitocondriale, cea mai mare parte a acestora fiind codificate de ADN-ul nuclear.
Intensitatea procesului respirator din mitocondrii este adaptat la cerintele celulare, printr-un mecanism de autoreglare foarte simplu, in functie de cantitatea de ATP si ADP prezente in celula. Intr-un tesut cu cheltuieli energetice importante se formeaza suficient ADP, (prin hidroliza ATP-ADP + P + energie) ceea ce va stimula activitatea respiratorie a mitocondriilor. Dimpotriva intr-un tesut bogat in ATP va fi o concentratie scazuta de ADP care va inhiba respiratia.
Rolul biologic al condriomului
In celulele eucariote energia necesara activitatilor vitale celulare se obtine prin degradarea
aeroba si completa a glucidelor, lipidelor, pana la stadiul de caz si H2O, degradare care este cuplata cu sinteza ATP. Prima etapa a acestor degradari are loc in citoplasma prin glicoliza anaeroba, etapa
finala are loc in mitocondrii, in care energia eliberata prin oxidarea substantelor, glucidice si lipidice in primul rand, este transformata in energie chimica sub forma legaturilor fosfodiesterice ale ATP. Astfel energia necesara unor procese metabolice ce se fac cu consum de energie, provine din hidroliza unei legaturi macroergice din ATP. Fata de glicoliza anaeroba din citoplasma, prin care glucoza este degradata numai pana la acid piruvic si prin care se formeaza doar doua molecule de ATP, prin oxidarea piruvatului de catre oxigenul molecular pana la CO2 si H2O randamentul este mult superior producand 34 molecule de ATP. Mentionam ca in regnul vegetal se produce ATP si la nivelul altor organite si anume la nivelul cloroplastelor, insa aici sursa de energie nu este glucoza ci
lumina solara utilizata eficient in procesul de fotosinteza.
S-a constat ca in mitocondrii si in cloroplaste se utilizeaza acelasi mecanism, chimiosmoza,
pentru a produce ATP din ADP si fosfat anorganic (Pi). Aceasta sursa de energie provine din gradientul de concentratie al protonilor si din potentialul electric membranar. Daca in fotosinteza, in cloroplaste, acest gradient este asigurat de energia absorbita de pigmentii clorofilieni din lumina, in
mitocondrii este utilizata energia rezultata din oxidarea glucidelor si lipidelor pentru pomparea ionilor de H+ (protonilor) prin membrana mitocondriala interna care genereaza gradientul de protoni. Aceasta forta, stocata in gradientul de protom, va fi folosita pentru sinteza ATP din ADP si Pi.
In celulele eucariote, catabolismul glucozei incepe in hialoplasma in care aceasta este transformata in doua molecule de acid piruvic cu formarea a doua molecule de ATP si reducerea a doua molecule de NAD+ la NADH + H+. In continuare, in mitocondrii, piruvatul este oxidat la CO2.
La inceput, in membrana mitocondriala externa, piruvatul este transformat in scetil-CoA (acetil coenzima A) si CO2. Aceasta substanta, acetatul activat de CoA, se formeaza si in timpul oxidarii, in mitocondrii, a acizilor grasi rezultati din lipide sau a aminoacizilor din proteine.
Rolul diferitelor componente In procesul respirator este prezentat mai jos.
Membrana mitocondriala externa este neteda si expune mitocondria in contact cu hialoplasma. Ea contine numeroase proteine canal (numite si porine) fiind foarte permeabila pentru ioni si molecule mici (cu GM<10.000). De aici rezulta ca doar membrana interna reprezinta singura bariera de permeabilitate a mitocondriei spre hialoplasma pe de o parte si spre matricea mitocondriala pe de alta parte. Aceasta membrana mitocondriala interna are o compozitie speciala avand cea mai mare proportie de proteine (76%) dintre toate membranele celulare. Ea contine si o fosfolipida foarte hidrofoba numita cardiolipina care este difosfatidil glicerol, avand rolul de a reduce permeabilitatea membranei la protoni si HO-. Membrana interna si matricea mitocondriala reprezinta sediul reactiilor de oxidare a acetatului (acetil CoA) si a acizilor grasi la CO2 si H20 cuplata cu sinteza ATP din ADP si Pi.
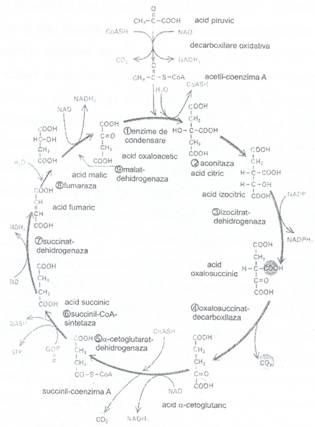
Fig. 51: Ciclul lui Krebs
Aceste procese cuprind mai multe etape si pot fi subdivizate in 3 grupuri de reactii:
1. Oxidarea acetatului si acizilor grasi la C02, cuplata cu reducerea transportorilor de electroni NAD+ si FADH2, care are loc la nivelul matricei mitocondriale.
2. Transferul electronilor de pe NADH + H+ si FAD2 la O2, care are loc la nivelul membranei interne si care este cuplat cu generarea unui gradient electrochimic de protoni de o parte si de alta a acestei membrane.
3. Utilizarea energiei din gradientul transmembranar de protoni pentru sinteza ATP de catre ATP sintetaza, la nivelul membranei mitocondriale interne.
Stadiul final al reactiilor de oxidare a glucidelor si lipidelor este reprezentat de ciclul Krebs (Fig. 51) al acizilor tricarboxilici sau ciclul citric, care consta dintr-un complex de 9 reactii enzimatice. In ciclui citric sunt oxidati 2/3 din totalitatea compusilor cu C ai celulei, produsii sai majori fiind CO2 si NADH. C02 este eliminat iar NADH + H+ transfera electronii de inalta energie pe care-i poarta in lantul respirator, fiind in final utilizati pentru a reduce O2 la H20 (Fig 51).
Ciclul Krebs incepe cu Acetil-CoA (C2) provenit din piruvat sau din acizii grasi, care este transferat pe molecule de acid oxalilacetic (C4), dand nastere astfel la acidul citric (C6). In continuare, printr-o serie de 7 reactii enzimatice, acidul acetic activat (acetilul) va fi complet degradat la 2C02, odata cu regenerarea acidului oxalilacetic si ciclul se reia.
Rezultatul final al acestor reactii este producerea a doua molecule de CO2, 3 molecule de NADH + H+, si una de FADH2 si a ATP, iar cea mai importanta contributie a ciclului Krebs la metabolism este furnizarea electronilor de inalta energie preluati tranzitoriu de NADH si F ADH2, ce vor fi apoi transferati in lantul respirator de pe membrana interna, in final ei fiind transferati la O2, ultimul acceptor de electroni din lant, formandu-se H20.
Oxigenul molecular (O2) este utilizat pentru a reoxida coenzimele reduse (NAD+, FAD). Energia care se elibereaza prin oxidarea unei singure molecule de NADH sau F ADH2 de catre O2 este suficienta pentru a sustine sinteza mai multor molecule de ATP. Ca urmare oxidarea NADH si sinteza ATP cuplate cu transferul de electroni au loc in mai multe etape (trepte) prin intermediul unor proteine care formeaza lantul transportator de electroni, iar energia va fi eliberata succesiv in cantitati mici. Deplasarea electronilor de la NADH+ H+ si FADH la O2 este catalizata de o serie de proteine purtatoare de electroni organizate in trei complexe macromoleculare pe membrana interna mitocondriala: NADH - dehidrogeneza, complexul b-C1 si citocrom-oxidaza.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5873
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved