| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Hormonii suprarenalieni
Glandele suprarenale (adrenale) sunt alcatuite din doua regiuni distincte, r.iu- au structuri, functii si origini embriologice diferite. Zona corticala (90%) arc o structura epiteliala, iar medulara (10%) este de origine nervoasa.
1. Hormonii medulosuprarenalieni
Zona medulara este alcatuita din celule cromafine (feocromocite).
Celulele cromafine au origini embriologice comune cu celulele ganglionare ale
sistemului nervos simpatic. Celulele cromafine se gasesc si in plamani, intestin,
ficat (celulele cromafine ectopice). Hormoni elaborati de medulosuprarenala se numesc catecolamine. Medulara impreuna cu sistemul nervos simpatic si parasimpatic alcatuiesc sistemul neuroendocrin - simpatoadrenal - ce are rol in adaptarea organismului la diverse solicitari.
Numele de catecolamine provine de la elementul de structura comun si anume nucleul catecol (ortodifenol) si functia aminica.
Catecolaminele sunt: dopamina, noradrenalina (norepinefrina) si adrenalina (epinefrina).
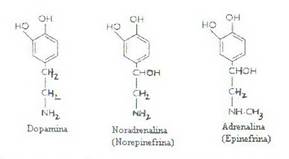
Dopamina se gaseste in anumite regiuni ale sistemului nervos central, avand rol de neurotransmitator la nivelul terminatiilor nervoase dopaminergice.
Dopamina este absenta in sange.
Noradrenalina reprezinta aproximativ 20% din secretia totala a medulosuprarenalei. indeplineste rolul de neurotransmitator la nivelul terminatiilor nervoase postganglionare simpatice, iar sangele cuprinde o fractiune de noradrenalina eliberata pe aceasta cale si care nu a fost recaptata la nivelul sinapselor.
Noradrenalina din celulele ectopice este sintetizata "in situ' sau captata din sange.
Adrenalina reprezinta aproximativ 80% din secretia totala medulosuprarenaliana. Nu se sintetizeaza in celulele ectopice, dar poate fi captata din plasma.
Biosinteza catecolaminelor
Precursorul catecolaminelor este tirozina. Transformarea tirozinei in adrenalina cuprinde 4 reactii:
- transformarea tirozinei in DOPA (3,4, dihidroxi fenilalanina), sub actiunea unei monoxigenaze (tirozin hidroxilaza)
decarboxilarea DOPA la dopamina intr-o reactie piridoxal-fosfat-dependenta
- o noua hidroxilare la nivelul catenei laterale sub actiunea hidroxilazei dependenta de ascorbat si Cu2 + rezultand noradrenalina
formarea
adrenalinei, prin metilarea la azot a noradrenalinei sub actiunea unei metil transferaze
specifice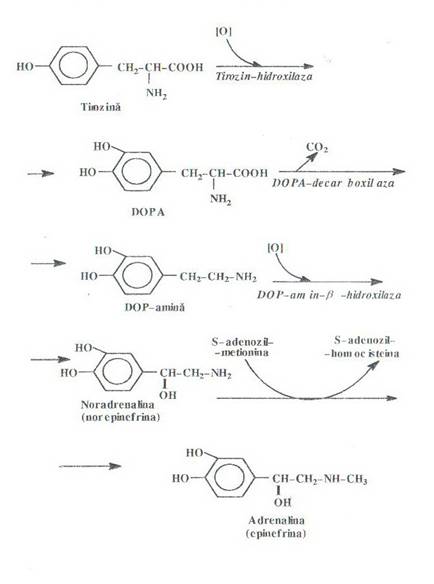
Biosinteza catecolaminelor se realizeaza prin utilizarea in calitate de precursor a tirozinei. La rindul ei, tiiozina poate pioveni din rezerva de aminoacizi a organismului, sau se poate forma prin hidroxilarea enzimatica a fenilalaninei. Sub actiunea tirozin-hidroxilazei
hepatice, tirozina este oxidata intr-o reactie ireversibila cu formμare de dihidroxi- fenilalanina (DOPA).
O parte importanta din cantitatea de DOPA astfel formata serveste la biosinteza pigmentilor melaninici . Restul se decarboxileaza cu formare de DOP-amina care prin hidroxilare la catena laterala formeaza adrenalina. La acest nivel intervine o enzima specifica numita N- metil-transferaza -care metileaza gruparea aminica a noradrenalinei cu formare de adrenalina.
Procesul de biosinteza a catecolaminelor prezinta un mecanism de reglare la nivel molecular prin faptul ca adrenalina si noradrenalina functioneaza ca inhibitori alosterici ai tirozin-hidroxilazei realizind astfel o inhibitie de tip feed back. in conditii fiziologic normale, organismul unui adult secreta y.ilnic 6 10 mg de catecolamine, iar concentratiile plasmaticc normale se situeaza in jurul valorii de 0,5 ug/1 pentru adrenalina si respectiv 3-5 μg/1 in cazul noradrenalinei.
Degradarea catecolaminelor se face foarte rapid in cazul hormonilor exogeni administrati intravenos (in cazul adrenalinei exogene de exemplu, timpul de injumatatire este de doar 10 -- 20 secunde).
Catecolaminele endogene se degradeaza mai lent, fiind apoi eliminate pe cale renala sub forma libera sau sub forma sulfo- si respectiv glucurono-conjugata. Catabolismul catecolaminelor se realizeaza in doua etape:
Etapa de 3-oxometilare are loc sub actiunea catecol-O -metil-transferazei (COMT) care transforma adrenalina in 3-oximetil-adrenalina sau metadrenalina, iar noradrenalina in normetadrenalina; COMT este o enzima cu masa moleculara de 24 kDa prezenta in citosolul hepatocitelor si celulelor renale si poseda o activitate catalitica foarte inalta.
Etapa de dezaminare oxidativa are loc sub actiunea monoaminoxidazei (MAO) specifice. Aceasta etapa se realizeaza mult mai lent, iar produsul de degradare (acidul
3-metoxi-4-hidroxi-mandelic sau acidul vanil-mandelic)se elimina prin urina.
Aceste etape de degradare sint distincte, metilarea putindu-se realiza atit inaintea cit si dupa dezaminarea oxidativa. De aceea, inhibitorii specifici ai MAO nu pot bloca degradarea catecolaminelor, in schimb ei inhiba catabolismul serotoninei cerebrale.
Reactiile ce au loc sunt:
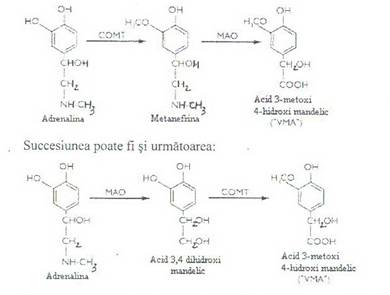
MAO opereaza transformari ale gruparilor aminice care prin oxidare la grupa imino trec in aldehide si apoi in acid.
Noradrenalina prin aceeasi succesiune de transformari formeaza acelasi produs final (acidul vanilmandelic)
Catabolismul noradrenalinei se realizeaza astfel:
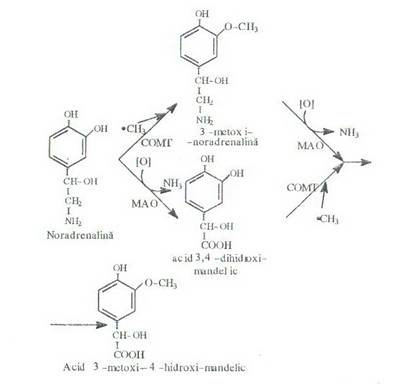
Acidul vanilmandelic se elimina prin urina si dozarea lui da indicatii asupra metabolismului catecolaminelor. La bolnavii cu feocromocitom (tumora a medulosuprarenalei) creste foarte mult nivelul urinar atat cel al metanefrinelor cat si al acidului vanilmandelic.
Catabolismul adrenialinei se realizeaza in mod asemanator. Si in aceasta situatie se poate realiza mai intii dezaminarea oxidativa si apoi metilarea produsului intermediar obtinut sau, se realizeaza mai intii metilarea adrenalinei dupa care are loc metilarea:
Catecolaminele din circulatia sanguina si cele eliberate la nivelul sinapselor din sistemul nervos central pot fi recaptate de catre celule pentru a fi stocate sau degradate, In primul caz este vorba de neuronii adrenergici din SNC si sistemul nervos vegetativ cind se intrerupe stimularea stimularea adrenergica datorita absentei unei activitati enzimatice extracelulare similare cu acetilcolin-esteraza. In cel de-al doilea caz,se realizeaza transportul catecolaminelor in in tesuturile periferice cxtra-neurale (muschi netezi, inima etc.) si degradarealor rapida
Rolul biologic al catecolaminelor
Principalele efecte pot fi impartite in doua categorii in functie de natura receptorilor acestor hormoni.
Efecte de tip α.Adrenalina are efectele cele mai puternice ,ea actionand sinergic cu noradrenalina si fenilefrina ,iar fentolamina are actiune antagonista. Principalele efecte de tip α constau in actiunea vasoconstrictoare cu cresterea presiunii arteriale ,actiunea asupra contractiei uterului si vezicii urinare ,stimularea eliberarii de acetilcolina la nivelul muschilor scheletici etc.
Aceste efecte sunt datorate existentei a doua tipuri de receptori specifici :receptori α 1
Care sunt Ca-dependente si intotdeauna post sinaptici,specifici pentru muschii netezi respectiv receptori α 2 ce pot fi post-si pre-sinaptici,specifici sistemului nervos central.In acest din urma caz este implicata si actiunea inhibitoare a AMPC.
Efecte de tip β.Efectele de tip β sunt diverse ,ele manifestandu-se la nivelul mai multor organe si tesuturi.Astfel,la nivelul inimii ,catecolaminele manifesta un efect inotrop pozitiv cu crestsrea frecventei contractiilor ,un efect cronotrop pozotiv.Se mai observa si o crestere a excitabilitatii si conductibilitatii cardiace. Asupra vaselor sanguine ,catecolaminele manifesta o actiune vasodilatatoare,iar la nivelul sistemului respirator o actiune bronhodilatatoare. La nivelul muscular ,acesti hormoni actioneaza in sensul relaxarii uterului ,vezicii urinare,si sfinctereelor inestinale ,iar in muschii striati este stimulata glicogenoliza.Catecolaminele manifesta si o actiune lipolitica la nivelul tesutului adipos.
Adenalina are si unele efecte metabolice si fiziologice specifice. La nivelul cardiac are loc o crestere a fortei sistolice si a debitului cardiac,iar efectele asupra vaselor sanguine sunt diferite,in functie de tesut sau organ.
La nivelul musculaturii netede ,adrenalina manifesta o actiune bronhodilatatoare puternica,o actiune de inhibare a tonusului si peristaltismului intestinal cu spaeme ale sfincterelor ,splenocontractie si crestere diurezei.
Cresterea brusca a secretiei de adrenalina are repercursiuni imediate asuora sistemului nervos central ,in special prin stimularea hipotalamusului cu cresterea secretiei de ACTH hipofizar,cand se instaleaza starea de stres.A cest efect a fost demonstrat prima data cand ,in urma injectarii de adrenalina,s-a instalat o stare de anxietate.Din punct de vadere metabolic,adrenalina stimuleaza hiperglicemia ca urmare a activarii glicogenolizei la nivel hepatic.
Reglarea activitatii medulosuprarenalelor
Medulosuprarenala este singura glanda endocrina supusa unui control preponderent nervos. Principalii centri nervosi adrenalino-secretori sint localizati la nivel bulbar, excitarea lor determinind o secretie abundenta de catecolamine.
Un exemplu al reglarii pe cale nervoasa a secretiei de catecolamine il reprezinta reglarea presiunii arteriale. Raspunsul medulosuprarenalei la modificarea presiunii arteriale nu este insa un raspuns direct. Aceste oscilatii actioneaza asupra unor receptori aortici specifici. Reactia umorala este tardiva si ea urmeaza raspunsului nervos.
Medulosuprarenala nu este esentiala pentru viata, dar fara secretia medulara organismul se adapteaza greu la diversi factori stresanti. Rolul catecolaminelor, prin actiunile de la nivelul sistemului circulator cat si asupra metabolismului energetic sunt implicate in adaptarea organismului la agresiuni interne sau externe, fizice sau psihice (frig, emotii, hipoglicemie, frica, etc.). Adrenalina in principal mediaza efectele metabolice ce au ca rezultat cresterea consumului de ATP, si de oxigen si eliberarea de caldura.
In ficat este stimulata glicogenoliza si gluconeogeneza, care asigura glucoza pentru mentinerea glicemiei si asigurarea combustibilului pentru creier. Inhibitia insulinei reduce consumul de glucoza in tesuturile periferice.
2.Hormonii corticosuprarenalieni
Portiunea corticala a suprarenalei este alcatuita din 3 zone distincte histologic:
- zona externa (zona glomerulara) producatoare de mineralocorticoizi
- zona mediana (zona fasciculata) producatoare de glucocorticoizi
- zona interna (zona reticulata) producatoare de hormoni androgeni. Structural, hormonii corticali sunt steroizi (corticosteroizi) ce contin in
molecula 21 atomi de carbon (C21) cei minerale- si gluco-corticoizi, iar hormonii androgeni contin 19 atomi de carbon (C 19).
Din categoria steroizilor ce exercita actiuni hormonale fac parte: cortisolul (glucocorticoid), aldosterona (mineralocorticoid),
dehidroepiandrosterona si androstendiona (androgeni).
in biosinteza steroizilor precursorul utilizat este colesterolul, ce poate fi procurat din urmatoarele surse:
-sinteza "de novo'
-captare din LDL (β-lipoproteine)
-din rezervele de acilcolesterol ale glandei.
Etapa initiala in biosinteza steroizilor, comuna pentru toti steroizi, este transformarea colesterolului in pregnenolona sub actiunea unei enzime ce contine citocromul P 450. Aceasta transformare are loc in mitocondrie. Prin hidroxilari succesive la C 22 si C 20 si sub actiunea unei enzime (desmolaza) si a citocromului P450, a oxigenului si a NADPH + H+ se scindeaza legatura C 22-C 20 eliberarea aldehidei izocaproice si formarea pregnenolonei (Fig. 2.15).
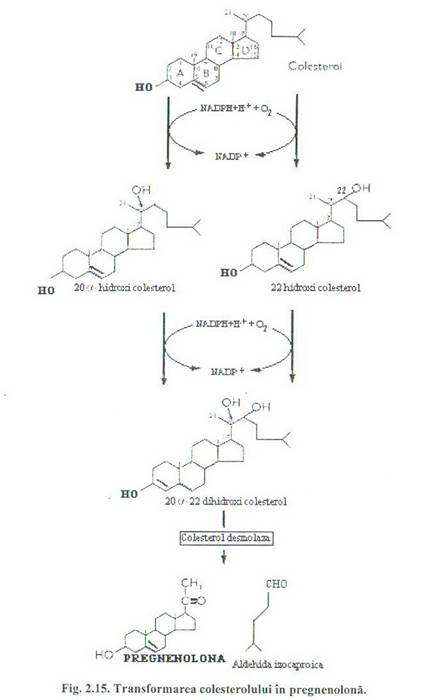
Aceasta etapa este reglata de ACTH in suprarenala si de catre LH in testicule si ovare.
Controlul secretiei prin ACTH implica:
- cresterea captarii de LDL. ca sursa de colesterol in zona fasciculata
- stimularea eliberarii colesterolului din esteri si inhibarea depunerii sale ca ester
- activarea conversiei colestrol →pregnelonona
Calea care duce la cortisol incepe fie prin hidroxilarea la C 17 (sub actiunea 17 α- hidroxilazei) si formarea 17 α-pregnelonona fie prin actiunea combinata a unei dehidrogenaze si a unei izomeraze (Δ5→ Δ 4 izomeraza) cu formarea de progesterona. 17α - hidroxilarea are loc numai in zona interna. Alte doua hidroxilari la C 21(sub actiunea 21-hidroxilazei) si la C11 (sub actiunea 11 β-hidroxilazei) se formeaza cortisol.
Calea care duce la aldosterona nu implica hidroxilarea la C 17Prin hidroxilare la C 21 si apoi la C 11se formeaza 11-dezoxicorticosterona ca actiune mineralocorticoida slaba si respectiv corticosterona cu actiune glucocorticoida. Sinteza de steroizi C 19 este cantitativ redusa si fara semnificatie, in acest sens pregnenolona prin hidroxilare la C 17urmata de oxidare si detasarea C 20si C 21 (sub forma unei molecule de acid acetic) se formeaza 17 -dehidroepiandrosterona sau 17 α- hidroxiprogesterona se transforma analog in androstendiona. Compusii C obtinuti in cortex nu au activitate hormonala ca atare decat dupa transformarea in testosterona si dchidrotestosterona in tesuturile tinta (fig. 2.16).
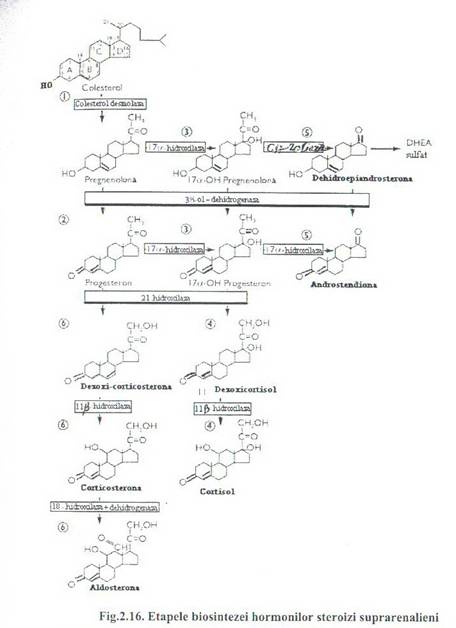
Cortisolul
Fractiunea libera este cea activa biologic. Cortisolul este vehiculat (70%) de catre CBG (cortisol binding globuline) sau transcortina si serubalbumina vehiculeaza 15%.
Timpul de injumatatire pentru cortisol este de 1.5-2 ore. Dupa unele remanieri (hidrogenarea dublei legaturi la C4 si reducerea gruparii cetonice de la Cs) are loc conjugarea cu acidul glucuronic si eliminarea prin urina a produsilor de conjugare.
Reglarea secretiei se face prin urmatoarele mecanisme:
prin feedback pe axul hipotalamo-hipofizar. Aceasta inseamna ca cresterea concentratiei de cortisol plasmatic inhiba secretia de CRH (hipotalamic) si de ACTH (adenohipofizar)
variatie diurna (ritm circadian), nivelul maxim fiind atins la trezirea din somn, iar nivelul minim seara factorii de stres mediaza pe cai nervoase eliberarea de CRH si ACTH care -stimuleaza secretia de cortisol. Evaluand efectele se poate concluziona ca raspunsul la stres, mediat de cortisol, este mai lent decat cel mediat de catecolamine, dar cu implicatii mai profunde in metabolism.
Mecanism de actiune
Cortisolul actioneaza la nivelul genomului celular prin intermediul receptorilor intracelulari prezenti in mai toate celulele.
Cortisolul stimuleaza sinteza unor proteine care au rol antiinflamator (ex. lipocortina) care inhiba procesul de sinteza a eicosanoizilor (cu actiune proinflamatoare). Deasemenea cortisolul represeaza sinteza unor proteine cheie care accentueaza procesele inflamatorii (ex. interleukina-1).
Actiuni biologice
in metabolismul intermediar exercita multiple actiuni anabolice si catabolice, in functie de natura tesutului, a starii organismului (nutritie sau foame) in functie de concentratia altor hormoni, etc.
Actiunea cea mai clara este stimularea gluconeogenezei hepatice si efluxul glucozei urmata de hiperglicemie. Actiunile sale metabolice sunt antagonice insulinei. Stimuleaza procesele catabolice la nivelul tesuturilor periferice (lipoliza trigliceridelor din tesutul adipos, proteoliza proteinelor din muschii scheletici, etc) cu eliberarea de substraturi pentru gluconeogeneza hepatica (glicerol, aminoacizi). La nivel hepatic stimuleaza gluconeogeneza prin aport crescut de substrat (aminoacizi, glicerol) precum si prin inductia enzimelor gluconeogenetice. Efectul hiperglicemiant al cortisolului este si rezultatul reducerii patrunderii de glucoza in tesuturile insulino-dependente (tesutul muscular si adipos) prin micsorarea afinitatii receptorilor pentru insulina in muschi si tesutul adipos.
Cortisolul exercita actiune imunosupresoare. Este antiinflamator mai puternic decat aspirina. Lipocortina stimulata de cortisol, inhiba fosfolipaza ai, intrerupand astfel cascada arahidonatului si productia de prosteaglandine si leucotriene (ce sunt mediatori ai raspunsului inflamator)
Aidosterona
Este hormonul corticosteroid (mineralocorticoid) ce participa la mentinerea homeostaziei hidrice si electrolitice.
Sinteza sa este controlata in principal de angiotensina si intr-o masura mai mica de concentratia plasmatica a potasiului (K+) si de ACTH. Circula in sange liber nu legat de serumalbumine care au afinitate mica dar capacitate mare de legare.
Reglarea secretiei
Se face prin urmatoarele mecanisme:
-sistemul renina-angiotensina
prin concentratia ionilor de potasiu (K+)
Sistemul renina-angiotensina are un rol important in reglarea volumului fluidului extracelular si a presiunii arteriale. Renina este o enzima proteolitica sintetizata si secretata de celulele juxtaglomerulare (renale). Aceste celule sunt inzestrate cu baroreceptori.
In plasma renina actioneaza asupra unei proteine plasmatice angiotensinogen (globulina sintetizata de ficat si secretata in sange). Angiotensinogenul in prezenta reninei pierde un decapeptid de la capatul N-terminal, numit angiotensina I. Angiotensina I, sub influenta unei enzime (enzima de conversie) este transformat hidrolitic in angiotensina II prin pierderea a 2 aminoacizi de la capatul C-terminal al angiotensinei I.
Angiotensina II este o substanta vasoactiva foarte puternica producand constrictia arteriolelor. in zona glomerulara, angiotensina II determina eliberarea aldosteronei. Cresterea concentratiei plasmatice a aldosteronei determina retentia de sodiu si secundar cresterea volumului si a presinuii sangelui. Angiotensina II actioneaza foarte scurt, fiind degradata rapid sub actiunea angiotensinazei (Fig.2.17).
Sinteza aldosteronei este promovata atat prin stimularea transformarii colesterol→pregnenolona cat si prin stimularea transformarii corticosteronei in aldosterona.
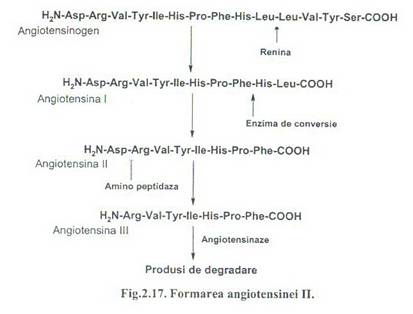
Enzima de conversie este eliberata de membrana plasmatica a celulelor endoteliului vascular si inhiba prin proteoliza peptide din clasa kininelor (ex. bradikinina), . antagonizand astfel actiunea vasodilatatoare a acestora. Bradikinina produce relaxarea muschilor netezi. Angiotensina II stimuleaza secretia de aldosterona si produce vasoconstrictie arteriala. Prin aceste actiuni mentine volumul si presiunea arteriala.
O alta cale de reglare a secretiei de aldosterona este prin concentratia ionilor de potasiu (K+) (Fig.2.18.)
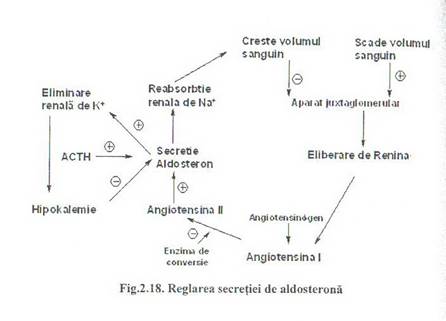
Reglarea secretiei de aldosterona prin concentratia ionilor de potasiu (K+) este un sistem independent de sitemul renina-angiotensina. O usoara crestere a Kalemiei stimuleaza secretia de aldosterona care prin actiune kaliurica va restabili valaorea kalemiei. Scaderea kalemiei inhiba secretia de aldosterona.
Actiuni biologice ale aldosteronei
Participa la mentinerea homeostaziei hidrice si electrolitice.
La nivel renal, determina reabsorbtia de Na+, antrenand prin aceasta retentia osmotica de apa si eliminarea renala a H+, K+, NH4+. La nivelul genomului celular, aldosterona induce sinteza unei proteine care este responsabila de transportul sodiului de-a lungul membranelor celulare.
La nivelul tubilor renali actiunea aldosteronei consta in stimularea transportului transcelular de sodiu din lumenul tubilor in spatiul interstitial, acest transport se face prin permearea pasiva a membranei apicale. Cand reabsorbtia sodiului nu se face concomitent cu reabsorbtia unei cantitati corespunzatoare de apa, atunci concentratia sodiului creste, iar cresterea presiunii osmotice va activa eliminarea hormonului antidiuretic. Acesta actioneaza la nivel renal asupra
pompei de apa, crescand reabsorbtia renala de apa. Aldosterona este inhibata de catre hormonul natriuretic atrial.
Aldosterona regleaza si transportul electrolitilor prin celulele epiteliale de la nivelul intestinului si al glandelor salivare si sudoripare.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5252
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2026 . All rights reserved