| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Motilitatea si chimiotaxia
Motilitatea bacteriana are doua particularitati: este foarte rapida si se face dupa o directie aleatorie. Motilitatea a fost studiata cu ajutorul unui microscop special (microscop de urmarire) la o mutanta monoflagelara de E. coli (fig 38).
Intr-un mediu neutru (fara atractanti si fara repelenti), celula bacteriana se deplaseaza dupa o traiectorie lineara, timp de aproximativ o secunda in care parcurge circa 30 mm (de 15 ori lungimea celulei) apoi se rostogolete timp de 0,l secunde si reia deplasarea lineara dupa o directie intamplatoare. S-a incercat sa se explice mecanismul alternantei deplasarii lineare si al rostogolirii. S-a reusit imobilizarea extremitatii libere a flagelului, cu anticorpi specifici, iar celula a ramas libera. S-a demonstrat astfel, ca motorul rotativ imprima flagelului doua tipuri de rotatii:
o micare de rotatie in sens orar
o micare de rotatie in sens antiorar.
La o celula peritriha, cand flagelii se rotesc in sens antiorar, ei formeaza un fascicul coerent, astfel incat imprima o micare de propulsie in linie dreapta. Cand sensul rotatiei flagelilor devine "orar", celula se dezechilibreaza si se rostogolete. Imediat dupa rostogolire, rotatia flagelilor devine "antiorara". In conditii normale de mediu (fara atractanti si fara repelenti), mobilitatea celulei peritrihe este intamplatoare, in directii imprevizibile si se face printr-o alternanta de delplasari in linie dreapta, intrerupte de rostogoliri.
Cercetarile de reologie au aratat ca datorita structurii flagelului, in mediul nutritiv lichid (bulion), deplasarea celulei bacteriene imtampina o rezistenta foarte mare. Cum se explica o viteza asa de mare, constanta in timp, intr-un mediu care opune o rezistenta foarte mare? Explicatia rezida in metabolismul foarte intens al celulei bacteriene si in natura particulara a sursei de energie utilizata. Pentru mobilitate, celula bacteriana nu utilizeaza ATP, ci energia electrochimica, adica fluxul de electroni care strabate regiunea bazala a motorului rotativ. Energia de rotatie a structurii bazale este energia protonica (chemiosmotica) generata de deplasarea protonilor prin membrana celulara, care furnizeaza forta inegalabila de deplasare a celulei bacteriene.
Motilitatea celulei bacteriene este influentata de prezenta in mediu, a substantelor chimiotactic pozitive (atractante) sau a substantelor chimiotactic negative (repelente) (fig. 39). Substantele atractante (aminoacizi, glucide, ioni de Ca si Mg, O2 pentru cele aerobe etc.) determina deplasarea celulei spre zonele in care concentratia lor este mai mare. Substantele repelente (alcoolii, acizii grai, O2 in forma ionica (O2--), OH--, metalele grele etc.) au efect chimiotactic negativ asupra celulelor bacteriene, determinand indepartarea lor din zonele cu concentratii mari.
Cand celula se deplaseaza intr-o directie favorabila (in directia unui atractant, sau se departeaza de un repelent), deplasarea este preponderent lineara si rostogolirile sunt mai putin frecvente, ceea ce semnifica o prelungire a perioadei de rotatie antiorara i o scadere a frecventei de rotatie orara a flagelilor.
Daca bacteria se deplaseaza intr-o directie nefavorabila sau intr-un mediu neutru crete frecventa rostogolirilor si scade corespunzator durata deplasarilor lineare. Rostogolirea are rolul de a corecta deplasarea celulei de la o directie greita. Reglarea frecventei rasturnarilor este esentiala pentru chimiotaxie.
Mobilitatea diferentiata se datoreaza prezentei, la suprafata celulei, a unor chemosenzori. Chemosenzorii sunt structuri moleculare specializate, alcatuite din chemoreceptor si proteinele de chimiotaxie, care au capacitatea de a accepta grupari metilice.
Chemoreceptorul inregistreaza prezenta atractantului sau repelentului in mediu si transmite informatiile chimice in celula. Chemoreceptorii sunt proteine de legare din structura membranei plasmatice si din spatiul periplasmic. Extremitatea NH2 a moleculei este extracelulara si are rolul de a lega semnalul chimic (atractant sau repelent). Unele molecule (aspartatul) se leaga direct de chemoreceptor, iar altele sunt purtate de proteinele de legare. Chemoreceptorii au o specificitate relativa fata de compusul chimic pe care il leaga. De exemplu, chemoreceptorul pentru galactoza, leaga glucoza si fucoza, iar cel pentru manoza leaga si glucoza.
Dupa legarea semnalului chimic, chemoreceptorul catalizeaza metilarea proteinelor de chimiotaxie, din categoria proteinelor transmembranare. Metilarea proteinei de chimiotaxie activeaza moleculele transductoare, care difuzeaza spre rotorul flagelar i comuta rotatia de la un sens la altul.
Gradul de metilare a proteinei de chimiotaxie este dependent de cantitatea de chemoefector din mediu.
In absenta factorilor atractanti sau repelenti, proteinele de chimiotaxie sunt demetilate de proteine citoplasmatice si producerea factorilor difuzibili este oprita.
Factorul difuzibil nu este cunoscut, dar probabil este o molecula mica.
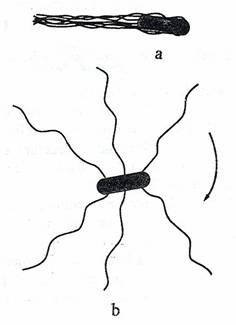
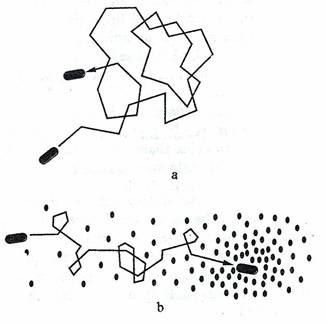
Fig. 38. a. Pozitia flagelilor la E. coli in timpul deplasarii lineare. Cand se rotesc in sens antiorar se strang intr-un manunchi si propulseaza celula. b. Cand flagelii se distribuie uniform, sensul de rotatie devine orar si celula se rostogoleste.
Fig 39. a. Intr-un mediu neutru, deplasarea lineara este intrerupta de rostogoliri, care schimba intamplator directia de deplasare. b. In prezenta unui factor chimic atractant, miscarile de rostogolire sunt partial suprimate. Deplasarea este orientata in directia atractantului.
Daca bacteria vine concomitent in contact cu un atractant si cu un repelent, raspunsul chimiotactic depinde de concentratia celor doi agenti si de afinitatea chemoreceptorilor care il leaga (fig 40). Daca concentratia repelentului este mai mare, bacteria se indeparteaza, si invers, daca atractantul are o concentratie superioara, celula se deplaseaza spre zona cu gradient maxim a acestuia.
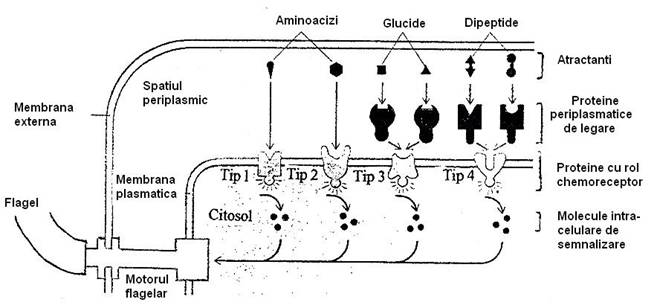
Fig. 40. Mecanismul transducerii semnalului chimiotactic la bacterii. Atractantii chimici se leaga de receptorii de tip 1 su 2 din membrana plasmatica sau de proteinele periplasmatice, care la randul lor se leaga de chemoreceptorii de tip 3 sau 4. Receptorii de chimiotaxie se activeaza si emit un semnal ce determina continuarea rotirii motorului flagelar in sens antiorar. Celula se deplaseaza preponderent in linie dreapta, iar rostogolirile sunt partial supresate (dupa Alberts si colab., 1994).
Chemosenzorul bacterian este primul care apare in lumea vie, dar este foarte sensibil si exact.
O proprietate definitorie a proteinelor cu rol transductor este adaptabilitatea la stimul, ce deriva dintr-un grad semnificativ de memorie a chemosenzorului, deoarece percepe modificarile de concentratie a substantelor chimice, atat in spatiu (de 20 de ori lungimea celulei), cat si in timp si dispune de un sistem de prelucrare a informatiei care moduleaza deplasarea celulei intr-un mediu cu atractanti si repelenti. Astfel, daca un atractant este adaugat in mediul de cretere, rostogolirea celulei este suprimata pentru cateva zecimi de secunda. Dupa un timp, frecventa rostogolirilor revine la normal, in prezenta continua a atractantului. Bacteriile raman in starea de adaptare la atractant, atata timp cat concentratia acestuia ramane constanta. Marirea concentratiei atractantului suprima din nou miscarile de rostogolire, iar indepartarea atractantului mareste frecventa rostogolirilor, pana cand adaptarea se produce din nou.
Rolul esential al chemoreceptiei este de a modula deplasarea celulei ca raspuns la schimbarile concentratiei unui component al mediului (si nu de a semmnaliza o concentratie constanta a acestuia), orientand deplasarea intr-o directie favorabila.
In suspensie, bacteriile flagelate se afla intr-o micare continua, dupa directii intamplatoare. Daca in mediu se creeaza gradienti ai factorilor fizici sau chimici, celulele se vor deplasa in acea zona a gradientului, care furnizeaza conditii optime. Deplasarea este rezultatul unui raspuns tactic (taxie). Cele mai cunoscute taxii sunt:
fototaxia asigura deplasarea bacteriilor fotosintetizante spre zonele cu luminozitate optima;
aerotaxia asigura deplasarea celulelor spre zonele cu concentratie optima a O2;
termotaxia. Temperaturile fiziologice sunt atractante, iar cele prea mari sau prea scazute sunt repelente.
Studiile de biomecanica si de reologie* considera ca mobilitatea celulei bacteriene este un caz particular.
Reologia este un domeniu al fizicii care studiaza forta de frecare si rezistenta pe care o intampina un corp solid in timpul deplasarii in functie de vascozitatea mediului lichid.
Viteza de deplasare este foarte mare, de 40-l00 de ori lungimea corpului pe secunda, fara echivalent in lumea vie. Datorita structurii flagelului, bacteriile intampina o rezistenta foarte mare in mediul lichid foarte vascos (bulion nutritiv). Explicatia deplasarii rapide consta in metabolismul foarte intens al celulei bacteriene si in natura particulara a sursei de E utilizata pentru miscarea de rotatie a flagelului. Pentru rotatia flagelului, celula bacteriana nu utilizeaza ATP, ci energia electrochimica, adica fluxul de protoni care strabate regiunea bazala a motorului rotativ. Energia protonica (chemiosmotica) furnizeaza forta pentru viteza de deplasare inegalabila a celulei bacteriene.
Semnificatia biologica a motilitatii. Motilitatea favorizeaza supravietuirea bacteriilor, oferindu-le posibilitatea sa se deplaseze spre zonele din mediu in care conditiile de existenta sunt mai favorabile. Bacteriile simbiotice fixatoare de N2 (Rhizobium) se deplaseaza spre radacinile plantelor leguminoase si initiaza procesul infectios. Desi deplasarile insumeaza 2 centimetri/zi, distanta este foarte semnificativa.
Pentru bacteriile patogene, mobilitatea (daca este prezenta) favorizeaza penetrarea stratului de mucus ce tapeteaza celulele epiteliale si faciliteaza aderenta bacteriilor, flagelul fiind astfel un factor de virulenta.
Alte tipuri de mobilitate. Mobilitatea flagelara este avantajoasa numai pentru microorganismele ce traiesc in mediul aquatic. Multe microorganisme traiesc in medii cu continut scazut de apa sau in medii care-si schimba gradul de umiditate: biofilme, sol etc. Unele bacterii flagelate produc un numar excesiv de flageli laterali, care le permite sa se deplaseze intr-un strat subtire de fluid pe o suprafata solida. Multe alte procariote se deplaseaza pe o suprafata solida fie prin alunecare, fie prin miscari bruste discontinui.
Mobilitatea prin alunecare se definete ca micarea unei celule neflagelate in directia axului celular lung, pe suprafata unor medii solidificate, sau in mediile lichide, la interfata aer-apa. Este o micare lenta, de tarare, care nu implica un singur mecanism. Mecanismele mobilitatii prin alunecare includ actiunea unor proteine fibrilare contractile, situate in straturile superficiale ale celulei; propagarea directionala a undelor de-a lungul suprafetei celulare; eliminarea orientata a mucilagiului; eliberarea controlata a surfactantilor la polii celulelor etc.
Celulele de Proteus (un zeu grec al marii, care ia multiple infatisari pentru a se face nevazut) pe suprafata mediului solidificat se deplaseaza prin roire (swarming). Roirea se produce pe o suprafata solida care nu permite mis carea flagelara obinuita in mediul lichid. Studiile de roire pe agar incep prin uscarea suprafetei agarului pentru a indeparta umiditatea. Astfel este impiedicata migrarea prin deplasarea in mediul lichid.
Procesul roirii cuprinde urmatoarele evenimente: a) creterea celulelor roitoare; b) migrarea lor pe suprafata agarului; c) diviziunea lor in celule scurte.
Comportamentul cunoscut sub denumirea de roire poate fi descris ca o miscare a celulelor alungite si flagelate pe suprafata mediului solid, in cicluri periodice de micare si consolidare. Celulele care roiesc sufera importante modificari morfologice. Roirea este determinata de aparitia celulelor filamentoase (de 20-80 um lungime), cu un numar mare de flageli (500-1000 flageli/celula), denumite celule roitoare. In timpul consolidarii, celulele roitoare se divid pentru a produce celule scurte (2-4 um lungime, cu 1-10 flageli) care cresc si se divid o perioada de timp, apoi se diferentiaza pentru a forma o alta generatie de celule roitoare. Celulele roitoare apar la periferia coloniei, pe suprafata libera, in grupe mici, care se deplaseaza pe o distanta scurta si se intorc din nou la colonie. Pe masura ce perioada de roire creste, celulele se deplaseaza in grupe mai mari, tot mai departe de colonie, inainte de a se reintoarce. Deplasarea in grupe mai mari este mai eficienta. La terminarea primei perioade de roire, micarea se oprete si celulele intra intr-o perioada de consolidare. Din celulele filamentoase, prin diviziune transversala in cateva puncte rezulta celule scurte normale. Celulele scurte rezultate prin diviziunea celulelor roitoare cresc si se divid si ciclul se repeta cu a II-a banda de roire, care incepe de la marginea primei benzi. Fenomenul se numete zonare si continua pana cand intreaga suprafata a placii este acoperita de cateva benzi concentrice de cretere densa si laxa. Miscarea poate fi rezultatul chimiotaxiei negative fata de catabolitii care se acumuleaza, sau al chimiotaxiei pozitive fata de substantele nutritive.
La spirochete, micarea este rezultatul contractiei i relaxarii unui fascicul de fibrile axiale (axostil), situate intre corpul celulei si o membrana ce acopera celula. Se produc micari pulsatorii prin flexia si extensia celulei, precum si o micare concomitenta de rotatie in jurul axului longitudinal.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1431
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved