| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Multiplicarea prin diviziune
Din punct de vedere structural, mecanismul diviziunii simple si etapele desfasurarii sale sunt bine cunoscute.
La bacteriile cilindrice, diviziunea simpla se face dupa un plan transversal, perpendicular pe axul lung al celulei. Foarte rar, diviziunea poate avea loc dupa un plan longitudinal. La coci, diviziunea se poate realiza dupa 1, 2 sau 3 planuri perpendiculare unul pe celalalt, rezultand diplococi, tetrade sau coci asezati in pachete cuboidale.
Nu exista un model unic al procesului de diviziune bacteriana. Studiile s-au facut la un numar limitat de specii, apartinand celor doua grupe majore: Gram pozitive si Gram negative.
Bacilii Gram pozitivi se divid dupa urmatorul mecanism: in regiunea ecuatoriala a celulei se formeaza un sept transversal, alcatuit din membrana citoplasmatica si peretele celular. Septul creste progresiv centripet, ca o adevarata diafragma si separa cele doua celule surori ce vor rezulta din diviziune.
La microscopul electronic se evidentiaza clar etapele succesive ale cresterii centripete a septului ecuatorial. Membrana citoplasmatica, uneori, precede usor septul parietal, dar niciodata nu progreseaza pentru a realiza singura separarea completa a celor doi protoplasti (fig 43).
La bacilii Gram negativi, diviziunea celulara are loc prin constrictia (strangularea) treptata a celulei la nivelul zonei ecuatoriale. Constrictia implica toate cele trei straturi: membrana externa, stratul peptidoglicanic si membrana interna. Se produce o crestere treptata a curburii spre interiorul celulei si o ingustare a ei in regiunea ecuatoriala.
Relatia dintre celula care se divide si celulele rezultate din diviziune este, o celula mama - doua celule surori. Celula care se divide dispare. Ea isi pierde individualitatea in cele doua celule intinerite. Diviziunea celulei este precedata de sinteza unor proteine, care se concentreaza la situsul formarii septului sau al constrictiei.
b
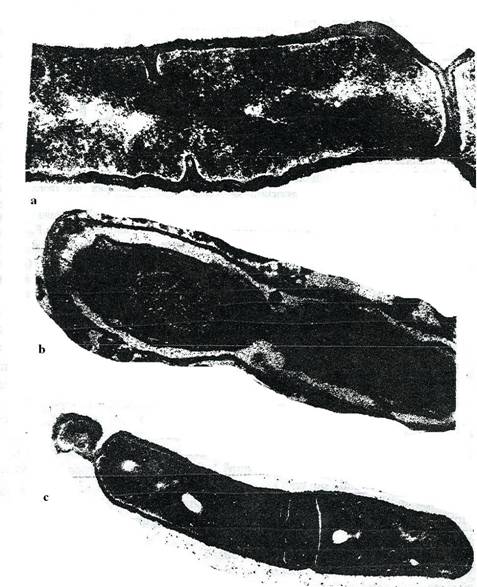
Fig. 43. Imagini electrono-optice in transmisie a celulelor bacteriene in diviziune. a. La B. subtilis, diviziunea are loc prin formarea unui sept de origine parietala. b. La bacteriile Gram negative diviziunea se face prin constrictia treptata a celulei la nivelul zonei ecuatoriale. c. Diviziunea asimetrica. La extremitatile ambelor celule de B. subtilis, diviziunea inegala genereaza minicelule (sageti) (original).
Mecanismul molecular al diviziunii
Procesul de diviziune este controlat de un set de 7 sau mai multe proteine cu functie specifica denumite proteine de diviziune. Proteinele care regleaza diviziunea, ca si cele care controleaza segregarea cromosomilor, motilitatea, nu sunt distribuite uniform in citoplasma, ci ocupa situsuri speciale: in membrana, in spatiul periplasmic etc.
Proteinele de diviziune se agrega in zona ecuatoriala a celulei si se asambleaza intr-o ierarhie bine determinata, formand o structura multimerica inelara - inelul Z, denumita septalsom, divisom sau septator. Diviziunea celulei se produce in zona inelului Z. La bacteriile Gram pozitive, pentru inelul Z este preferat termenul de septalsom (deoarece diviziunea este precedata de formarea septului), iar la bacteriile Gram negative se foloseste termenul de divisom, pentru ca diviziunea progreseaza prin constrictie.
Moleculele implicate in diviziunea celulara si in segregarea cromosomilor s-au evidentiat prin metoda microscopiei cu fluorescenta. La E. coli, ele se asambleaza la nivelul situsului de diviziune si formeaza ansamblul functional de forma inelara - divisomul. Toate proteinele reglatoare ale procesului de diviziune sunt proteine termosensibile, deoarece la temperatura nepermisiva, functia lor este alterata, nu mai controleaza procesul de diviziune si determina aparitia mutantelor filamentoase. Celulele filamentoase se formeaza ca rezultat al cresterii celulei, inotita de replicarea repetata a materialului nuclear, dar celula nu se divide. De aceea, multe proteine si mutantele pe care le genereaza au prefixul Fts (filamente termosensibile).
Majoritatea proteinelor Fts au un domeniu ancorat in membrana celulei, iar celelalte domenii se gasesc in citoplasma sau in spatiul periplasmic.
S-au identificat functiile biochimice pentru 2 proteine : FtsZ si PBP (penicilin binding protein).
Cea mai importanta cantitativ (de ordinul miilor de molecule), dintre proteinele de diviziune este FtsZ, cu localizare ecuatoriala, care in forma polimerizata, localizeaza procesul de diviziune. Fts Z este o proteina citosolica si se gaseste virtual la toate bacteriile si in organitele celulei eucariote. Este omologa tubulinei si se poate polimeriza ca si tubulina, pentru a forma o structura asemanatoare unui inel (inelul Z), la situsul de diviziune. Inelul Z are un rol esential in constrictia membranei celulare si in coordonarea intregului proces de diviziune. Fts Z este o GTP-aza si poate sa se polimerizeze atat in vitro cat si in vivo.
Proteina care leaga penicilina (PBP) are rolul de sinteza a peptidoglicanului parietal la noii poli ai celulei (pentru organismele care au perete peptidoglicanic). PBP sunt enzime care catalizeaza reactia de legare incrucisata intre polimerii peptidoglicanului: sunt transpeptidaze care leaga incrucisat catenele tetrapeptidice asociate acidului N-acetilmuramic. Legarea antibioticelor b-lactamice la PBP implicate in formarea septului de diviziune are ca rezultat formarea filamentelor lungi. Inactivarea altor PBP duce la liza rapida a celulei bacteriene, deoarece celula ramane osmotic neprotejata de perete.
Diviziunea celulei parcurge cateva etape :
- selectia situsului de diviziune, de obicei la mijlocul celulei, intre nucleoizii recent segregati;
- asamblarea aparatului citoplasmatic, care aproape todeauna implica FtsZ si la majoritatea organismelor, FtsA. Aceste proteine leaga si hidrolizeaza nucleozidtrifosfatii, eliberand energia necesara remodelarii celulei.
In celulele bacilare gata sa inceapa diviziunea sunt 3 situsuri la care procesul se poate initia: unul in zona centrala a celulei, intre cromosomii replicati si segregati si cate unul la polii celulei.
Evenimentul central al inceperii diviziunii este formarea inelului Z. Localizarea sa la mijlocul celulei este controlata prin doua mecanisme:
mecanismul inchiderii nucleoidului, se bazeaza pe observatia ca diviziunea celulei este initiata in spatiile libere, in care ADN lipseste ;
sistemul min, care controleaza pozitia inelului Z, denumit astfel pentru ca mutantele acestor gene produc minicelule prin localizarea inadecvata, la polii celulei, a situsului de diviziune.
Forta motrice a citochinezei pare a fi depolimerizarea controlata a inelului format de proteinele Fts, deoarece diametrul divisomului scade progresiv in timpul diviziunii, dar si cresterea centripeta a stratului de peptidoglican.
Diviziunea asimetrica
De cele mai multe ori, diviziunea este simetrica sau izomorfa, adica din procesul de diviziune rezulta doua celule de dimensiuni egale (fig. 42c).
Uneori, diviziunea celulara este asimetrica, rezultatul fiind formarea a doua celule inegale. Cea de dimensiuni mici este o minicelula. Diviziunea asimetrica este caracteristica bacteriilor cilindrice si se produce frecvent la mutante de E. coli si B. subtilis.
Minicelulele (bacterii miniaturale) au fost descrise in l967 de Adler ca bacterii foarte mici, care apar la extremitatea celulelor de E. coli, dupa diviziune.
Minicelulele apar foarte rar la bacteriile din mediile naturale, dar sunt frecvente la liniile mutante, crescute pe mediile artificiale. In general, minicelulele sunt lipsite de material nuclear, sau contin o cantitate foarte mica de ADN. Nu pot creste si nici nu se divid, dar au activitate metabolica: realizeaza procese oxidative, produc energie sub forma de ATP.
Adeseori, minicelulele contin plasmide. In cazul in care plasmidele le confera calitatea de donoare de material genetic, minicelulele pot realiza procese conjugative de transfer de material genetic, unor celule receptoare. Ele pot fi infectate de fagi, dar elibereaza un numar mic de fagi progeni.
Initial li s-a atribuit numai o importanta de ordin teoretic, minicelulele fiind structuri ce permit analiza unor procese biologice importante: diviziunea celulara, studiul fenomenelor de transport celular prin membrana. Interesul fata de ele a crescut, legat de posibilitatea de a le folosi in viitor pentru producerea de vaccinuri, chiar in cazul in care provin de la bacterii patogene, deoarece fiind lipsite de capacitatea de diviziune, nu pot initia procese infectioase.
Mecanismul formarii minicelulelor nu este elucidat. Una din teoriile care explica diviziunea asimetrica presupune ca in fiecare celula bacteriana cilindrica exista trei situsuri la care se poate initia formarea unui perete despartitor: unul situat central si celelalte doua, distribuite simetric la cei doi poli. In mod normal, bacteriile cilindrice se divid printr-un perete transversal ce se formeaza la nivelul situsului central. Cele doua celule surori mostenesc cate un situs polar. Pe masura ce cresc, situsul polar dobandeste o pozitie centrala. Formarea unei minicelule ar fi consecinta activarii premature a unui situs polar, la nivelul caruia se formeaza un sept de diviziune, inainte ca procesul de crestere celulara sa se realizeze complet.
Aceasta teorie nu explica modul de formare concomitenta a minicelulelor "in siraguri", ceea ce presupune activarea simultana a tot atatea situsuri de diviziune.
Formarea minicelulelor pare a fi rezultatul unor mutatii la nivelul genelor reglatoare ale procesului de diviziune.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1518
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved