| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Oncogenele retravirale
Toate oncogenele retravirale sunt derivate din genomul gazdei si reprezinta o achizitie recenta.
Cele mai productive surse de oncogene virale au fost animalele la care infectia cu virusurile leucemice este comuna: puiul de gaina, pisicile de casa care fac frecvent tumori. In tabelul de mai jos sunt redate denumirile celor mai reprezentative oncogene retravirale.
|
Oncogena |
Virusul transductor |
Sursa de celule maligne |
|
Abl |
Virusul leucemiei murine Abelson (MuLV) |
Limfomul celulelor B de soarece si om |
|
Erb A, erb B |
Virusul eritroblastozei aviare (VEA) |
Fibrosarcom si leucemie de pui de gaina |
|
|
Virusul sarcomului felin |
Fibrosarcom felin |
|
Fos |
Virusul osteosarcomului murin (FBJ MuSV) |
Condrosarcomul de soarece |
|
Fps |
Virusul sarcomului Fujinami (FuSV) |
Sarcom de pui de gaina |
|
H-ras |
Virusul sarcomului murin |
Sarcom de sobolan, carcinom uman si de sobolan |
|
K-ras |
Virusul sarcomului murin Kirsten (Ki-MuSV) |
Sarcom de sobolan, carcinom si sarcom uman, leucemii umane |
|
Myb |
Virusul mieloblastozei aviare (AEV) |
Celule leucemice de pui de gaina si celule leucemice umane |
|
Rel |
Virusul reticuloendoteliozei |
Leucemie limfoida de curcan |
|
Sis |
Virusul sarcomului simian (VSS) |
Sarcom de maimuta |
|
Src, ski, yes |
Virusul sarcomului Rous |
Sarcom de pui de gaina |
Denumirile protooncogenelor sunt prescurtari sau acronime pentru virusurile sau tesuturile de origine. Aceste gene codifica sinteza unor tirozin-kinaze (src), treonin-kinaze, a unor proteine care se asociaza cu ADN si au rol in reglarea
Studiile de hibridare moleculara in situ au argumentat ca gena v-src a virusului sarcomului Rous (VSR) este de origine celulara: gena v-src a fost incorporata de virus din ADN celular si astfel virusul a dobandit proprietati oncogene.
Ipoteza originii celulare a v-src este sprijinita de faptul ca mutantele virale rezultate prin deletia genei src, se multiplica normal. Rezultatele se pot extrapola pentru toate oncogenele transduse de retravirusuri.
Genele retravirale oncogene nu contin introni, in timp ce oncogenele celulare sunt totdeauna discontinui. Exista si alte gene care si-au pierdut intronii, dar nu se cunoaste nici una care sa-i fi dobandit.
Virusurile transductoare au o eficienta foarte mare de transformare a celulelor in vitro (24-48 de ore) iar in vivo produc tumori la cateva zile dupa inoculare.
Virusurile transductoare sunt defective, deoarece in capsida poate fi impachetata o cantitate limitata de ARN genomic. O parte a informatiei genetice virale, este inlocuita de genele transduse. Din aceasta cauza, virusurile transductoare sunt mutante defective ale ciclului de replicare (fig. 103), deoarece oncogena celulara a inlocuit total sau partial o secventa esentiala pentru replicare. Retravirusurile defective codifica majoritatea etapelor ciclului de multiplicare: revers-transcrierea, integrarea, sinteza ARN si traducerea (fig. 104) . Ele produc transformarea maligna a celulelor pe care le infecteaza, dar nu se asambleaza virus progen, deoarece nu se sintetizeaza proteine structurale. Virionii progeni se asambleaza numai daca aceiasi celula este infectata de un retravirus cu ciclu complet de replicare, care suplineste deficientele virusului defectiv.
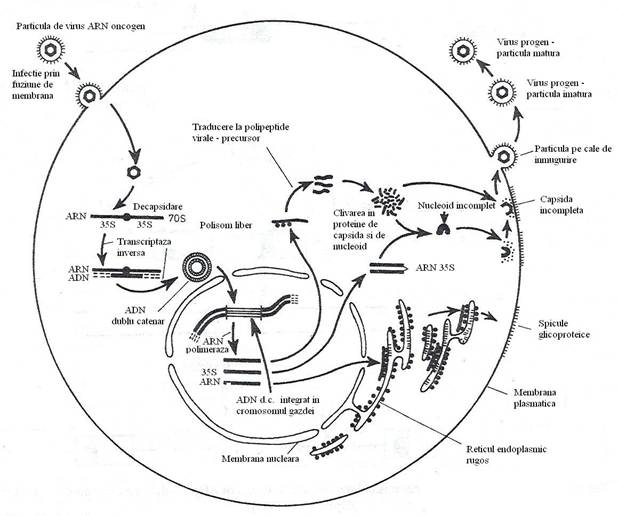
Fig. 103. Reprezentarea schematica a principalelor etape ale ciclului de replicare a virusurilor tumorale ARN.
Virionul are rolul unui vehicul transductor pentru oncogenele de origine celulara. Dupa ce au fost transduse intr-o noua celula, transcrierea lor este controlata de genomul viral.
Gena v-src transdusa de VSR este o exceptie. Ea se adauga intregului set de gene virale si virionii transductori nu sunt defectivi pentru ciclul de replicare. Deletia genei v-src nu influenteaza capacitatea de replicare a VSR.
Fenomenul transductiei este un eveniment rar. Virusurile transductoare nu pot fi considerate ca agenti infectiosi naturali, ci ca accidente ale ciclului de replicare, deoarece virionii transductori sunt defectivi.
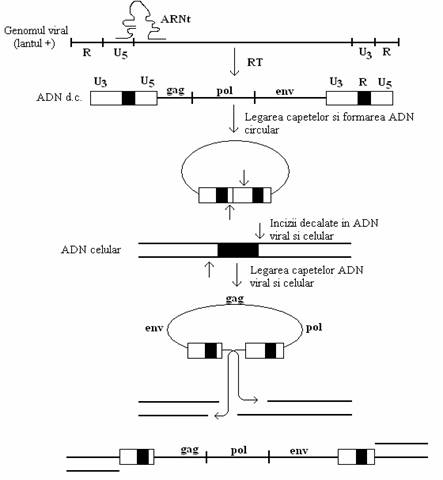
Fig. 104. Ilustrarea schematica a mecanismului molecular al integrarii ADN viral intr-un cromosom al celulei (dupa Coffin, 1991).
Mecanismul transductiei genelor celulare este ipotetic. Se admite ca integrarea genomului viral ca provirus, are loc in proximitatea unei oncogene celulare, in amonte, in raport cu promotorul ei, in aceiasi orientare de transcriere. Transcrierea genomului viral, catalizata de aparatul enzimatic al celulei, se extinde asupra oncogenei celulare, deoarece gena c-onc trece sub controlul promotorului viral. Astfel, este transcrisa o molecula himera de ARN, care contine atat copia provirusului cat si a oncogenei celulare. Copia de ARN este prelucrata prin mecanismul clivarii si inadirii (intronii sunt excizati) si ARN himeric va forma genomul virionilor progeni.
Oncogenele de origine celulara sunt transduse sub forma unei copii de ARN. Un virus transduce o singura gena, rareori doua. Genele onc transduse de retravirusuri sunt cei mai eficienti agenti ai transformarii maligne.
Oncogenele de origine celulara s-au gasit numai la subfamilia Oncovirinae, dar lipsesc la Lentivirinae si Spumavirinae, desi treptele replicarii sunt aceleasi.
Mecanismele posibile ale transformarii maligne sub actiunea retravirusurilor transductoare, sunt multiple.
1) In acord cu teoria supradozajului, transformarea maligna este rezultatul sintezei in exces a oncoproteinelor codificate de genele transduse. Eficienta transformanta a retravirusurilor transductoare, deriva din faptul ca genele v-onc, fiind sub controlul promotorului viral, sunt transcrise cu o rata superioara, comparativ cu genele c-onc. Daca oncogena celulara codifica un factor stimulator al sintezei ADN, cresterea ratei diviziunii celulare este prima treapta spre transformarea maligna.
2) Sinteza oncoproteinelor modificate. In celulele normale ale muschiului pectoral de gaina, gena c-src codifica o tirozin-kinaza, o fosfoproteina de 60 kDa, care se insera in membrana citoplasmatica si nu are, in mod obisnuit, proprietati transformante. Ea fosforileaza resturile de tirozina ale catorva proteine din categoria integrinelor. Proteinele tirozin-fosforilate, in celula normala, au rolul de a lega Ca2+ si functioneaza ca receptori pentru fibronectina. Activitatea tirozin-kinazei codificata de c-src este de numai 10% din aceea a enzimei codificata de gena v-src.
Genele v-onc transduse, codifica sinteza unor oncoproteine modificate. De exemplu, gena v-src codifica o tirozin-kinaza usor modificata fata de tirozin-kinaza codificata de gena c-src: ultimii l9 aminoacizi ai capatului C-terminal al proteinei C-src sunt inlocuiti cu o secventa de l2 aminoacizi in proteina V-src. Modificarea are consecinte asupra efectului sau reglator, dar gena v-src este transcrisa cu o rata net superioara, fiind sub controlul promotorului viral.
3) Modificarea pozitiei genei este unul dintre mecanismele prin care o gena celulara transdusa devine oncogena. Genele v-onc, originare in celula, la un nou ciclu infectios se integreaza concomitent cu provirusul, intr-o noua localizare cromosomala, in ambianta altor gene celulare.
Oncogeneza cu retravirusuri netransductoare. Multe retravirusuri care nu transduc oncogene celulare, induc tumori dupa inoculare experimentala la animale: sarcoame, leucemii, carcinoame. Diferenta majora fata de virusurile oncogene transductoare, consta in perioada de latenta care precede aparitia tumorilor: de la cateva saptamini, la cateva luni. Nici unul dintre virusurile netransductoare nu induce transformarea maligna a celulelor in vitro. Cel mai cunoscut reprezentant al acestui grup este virusul leucozei aviare. Tumorile induse de retravirusurile netransductoare au cateva proprietati definitorii:
toate celulele unei tumori contin genomul integrat ca provirus
provirusul este integrat in acelasi situs cromosomal, in toate celulele tumorii
celulele tumorii sunt clonale (identice din punct de vedere genetic), avand originea intr-o singura celula transformata.
Integrarea provirusului se face la situsuri specifice ale unui cromosom, in imediata vecinatate a oncogenelor celulare, ca si genomul virusurilor transductoare. Consecinta directa a integrarii genomului viral, este mutageneza prin insertie.
Integrarea genomului viral este mutagena, deoarece modifica secventa de ADN. Mutatiile induse in oncogenele celulare pot avea doua efecte: stimuleaza activitatea unei gene sau o inactiveaza.
Modificarea activitatii oncogenelor celulare constituie esenta mecanismului molecular al transformarii maligne si a fost explicata prin mai multe teorii:
l) teoria supradozajului considera ca transformarea maligna este rezultatul stimularii activitatii unei oncogene si a sintezei in exces a oncoproteinei pe care o codifica. Gena c-onc din proximitatea situsului de integrare a provirusului, trece in subordinea promotorului viral si va fi transcrisa cu o rata superioara celei normale. Oncoproteina corespunzatoare se sintetizeaza in exces, are actiune stimulatoare si activeaza necontrolat rata diviziunii celulare.
2) teoria activarii genelor c-onc sub actiunea inductoare a provirusului considera ca genele c-onc, in mod obisnuit sunt partial sau total represate, dar se activeaza dupa integrarea provirusului in imediata lor vecinatate. Se sintetizeaza oncoproteine, inductoare ale transformarii maligne. Acesta este fenomenul activarii prin insertie sau al cis-activarii oncogenelor celulare, consecutiv trecerii lor sub controlul promotorului viral.
3)inactivarea antioncogenelor este un alt efect al mutagenezei prin insertie. Antioncogenele sunt inactivatoare ale protooncogenelor. Ele codifica proteine supresoare ale protooncogenelor celulare, al caror rol este mentinerea functiilor normale ale celulei. De exemplu, retinoblastomul (o tumora retiniana ce apare in copilarie) se datoreaza pierderii ambelor copii ale genei rb. Fosfoproteina RB, cu localizare nucleara, are rol important in reglarea ciclului celular, controland intrarea celulei in ciclul mitotic. Absenta proteinei RB, are ca efect proliferarea necontrolata a celulelor retiniene.
Gena p-53, supresoare a oncogenelor poate fi inactivata prin insertia unui provirus la ambele alele, sau prin combinarea unui eveniment de insertie, cu pierderea celeilalte alele prin deletie.
Genele c-onc se pot activa independent de infectia virala sub actiunea mutatiilor, chiar punctiforme, care apar in interiorul lor. Substitutia unui singur aminoacid al oncoproteinei este suficienta pentru a converti o oncoproteina reglatoare, intr-o proteina cu caracter transformant.
Activarea oncogenelor ar putea fi rezultatul translocatiei unei protooncogene intr-o noua localizare cromosomala. De exemplu, in celulele limfomului Burkitt, schimbul de fragmente intre cromosomii perechii 8 si l4 are o frecventa de 90%. Schimbul intre cromosomii perechilor 8 si 2, ca si intre perechile 8 si 22 are o frecventa de 5%. In toate cazurile are loc translocatia unui fragment al unui cromosom din perechea numarul 8. Acest fragment contine protooncogena c-myc, care astfel este translocata adiacent fata de o gena reglatoare a sintezei anticorpilor. In noua lcalizare a genei, oncoproteina c-myc este sintetizata in exces.
Oncogenele celulare mediaza transformarea maligna indusa de virusuri, dar si de factorii fizici si chimici cu actiune mutagena.
Infectiile cu retravirusuri potential oncogene, la om si animale sunt mult mai frecvente decat incidenta tumorilor. Pentru ca transformarea maligna a celulei sa se produca, este necesar ca simultan cu infectia virala, sa actioneze alti factori cu efect sinergic, denumiti factori cocarcinogeni. Acestia pot fi factori intrinseci (genetici, hormonali, imunologici) sau factori ai mediului extern (substante chimice potential cancerigene, radiatii). Factorii cocarcinogeni interni sau externi potenteaza evolutia spre malignizare a infectiei cu virusuri oncogene.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1888
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved