| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Din numarul mare de microorganisme existente in natura, un numar limitat sunt capabile sa se dezvolte in asociatie cu organismele superioare, iar dintre acestea, un numar restrans (cateva sute) sunt patogene.
Bacteriile patogene au doua proprietati definitorii: patogenitatea si virulenta. Cei doi termeni au semnificatie distincta.
Patogenitatea este proprietatea unui microorganism de a determina in conditii naturale sau experimentale, aparitia unui proces infectios, decelabil din punct de vedere clinic, la o gazda receptiva.
In mod obisnuit, patogenitatea este asociata, inexact, cu modul de viata parazitara a microorganismelor, dar calitatea de patogen nu se asociaza totdeauna cu parazitismul. Majoritatea bacteriilor patogene sunt parazite, iar in vitro se dezvolta ca saprobionte pe medii organice, uneori cu compozitie chimica complexa. Unele bacterii, desi sunt strict saprobionte, adica se dezvolta pe materia organica in mediile naturale (de exemplu, Cl. botulinum), dar nu sunt parazite, produc totusi efecte patologice prin intermediul exotoxinelor pe care le secreta.
Patogenitatea este un caracter de specie. De exemplu, B. anthracis, C. diphteriae, S. typhi, M. tuberculosis sunt patogene sau conditionat patogene, adica determina aparitia unui proces infectios numai daca sunt inoculate in doze foarte mari sau daca organismul are o rezistenta generala scazuta. Cel mai ilustrativ exemplu de patogenitate conditionata este oferit de Ps. aeruginosa, o bacterie saprobionta in mediile naturale: organismul uman impiedica, in conditii obisnuite, colonizarea pseudomonadelor, dar la persoanele imunocompromise, in ranile provocate de arsuri sau de traumatisme sau in tesutul pulmonar al pacientilor cu fibroza chistica, Ps. aeruginosa totdeauna produce infectii.
Alte specii nu sunt niciodata patogene, pentru ca nu au capacitatea de a stabili interactiuni cu organismele superioare: Azotobacter, Nitrobacter, Acetobacter etc.
Patogenitatea implica existenta a patru proprietati esentiale:
a) capacitatea de a patrunde si de a se localiza in organismul gazdei;
b) proprietatea de a se multiplica in tesuturile acesteia si de a le invada;
c) rezistenta la mecanismele de aparare ale gazdei;
d) producerea leziunilor tisulare care se pot exterioriza clinic.
Virulenta este capacitatea unei tulpini a unui microorganism patogen aflat intr-o anumita faza de crestere, de a se localiza (a coloniza), de a se multiplica si eventual de a invada celulele si tesuturile gazdei si/sau de a produce toxine, determinand o stare patologica la o gazda receptiva.
Virulenta este o proprietate biologica corelata cu anumite caractere de structura (prezenta flagelilor, fimbriilor), cu prezenta unor proteine membranare sau cu unele particularitati fiziologice si de sinteza (ca de exemplu sinteza exotoxinelor sau a exoenzimelor).
Virulenta exprima cantitativ gradul de patogenitate a unei tulpini pentru o anumita gazda si este o proprietate multifactoriala:
infectiozitatea (sau capacitatea de colonizare)
agresivitatea (sau invazivitatea)
toxigenitatea
a) Infectiozitatea reprezinta capacitatea unui microorganism de a depasi mijloacele de aparare a organismului, de a se implanta si de a coloniza tesuturile sanatoase, adica de a stabili o localizare si de a forma un focar primar de infectie.
Colonizarea tesutului de catre un microorganism patogen este o faza initiala critica pentru evolutia procesului patologic si este un fenomen complex, care implica atat implantarea cat si utilizarea substantelor nutritive disponibile in mediul gazdei.
Bacteriile produc o larga varietate de factori, care usureaza capacitatea lor de a invada gazda, de a se raspandi in tesuturi si de a supravietui mecanismelor de aparare ale gazdei. Moleculele au fost grupate in 4 clase: adezine, invazine, agresine si impedine.
Adezinele sunt proteine bacteriene care mediaza interactiunea celulei bacteriene cu o varietate de celule ale gazdei, in special epitelii, ca o prima treapta de colonizare a gazdei. Invazinele sunt proteine bacteriene care permit celulei sa intre in celula eucariota. Agresinele sunt molecule, de exemplu toxine si proteaze, care produc leziuni ale gazdei ori favorizeaza raspandirea infectiei. Impedinele sunt componente bacteriene care inhiba actiunea mecanismelor de aparare, fara sa produca leziuni.
Prima etapa a colonizarii unui tesut este aderenta celulelor bacteriene de celulele gazdei
Legarea bacteriilor patogene de celulele sensibile este etapa initiala a procesului infectios. Aderenta impiedica indepartarea bacteriilor prin fluxul secretiilor, prin tuse, prin motilitatea cililor sau prin peristaltism.
Aderenta este mediata de structuri specializate ale suprafetei bacteriene, cunoscute sub denumirea generica de adezine.
Structurile bacteriene de aderenta, anatomice sau moleculare sunt, de cele mai multe ori, adaptative. Ele dispar prin cultivarea succesiva in vitro. De aceea, tulpinile bacteriene de laborator sunt mai putin aderente de suport, comparativ cu izolatele bacteriene recente.
Aderenta asigura colonizarea anumitor situsuri din organism, multiplicarea bacteriilor, sinteza toxinelor si desfasurarea reactiei inflamatorii de aparare.
Bacteriile adera in special de epiteliile mucoaselor, dar si de epiteliile keratinizate, de endotelii, de tesutul osos, de smaltul dentar etc. Aderenta implica interactiunea situsurilor complementare de pe suprafata celor doua tipuri de celule: epiteliala si bacteriana.
Aderenta este conditionata de complementaritatea sarcinilor electrice ale celor doua suprafete. Cele mai multe bacterii au o sarcina neta negativa a suprafetei lor, dar au si zone limitate electropozitive, precum si molecule cu caracter hidrofob.
Aderenta reprezinta un avantaj ecologic major pentru bacteriile patogene, cu privire la asigurarea nutrientilor, protectia fata de anticorpi si lizozim etc. Multiplicarea lor, dupa aderenta, are loc cu o rata net superioara fata de a celulelor neaderente.
Mecanismele aderentei bacteriene in vivo au fost studiate in special pentru bacteriile generatoare de carii dentare. Aderenta celulelor bacteriene patogene, de celulele sensibile implica participarea a doi factori: o adezina localizata pe suprafata bacteriei si un receptor de pe suprafata celulei gazda.
Adezinele bacteriene se impart in doua mari categorii: a)adezine de natura proteica; b) adezine neproteice.
Grupul adezinelor proteice cuprinde fimbriile. Acestea sunt structuri filamentoase, neflagelare ale suprafetei celulare si sunt diferite de pilii participanti la conjugare. Fimbriile fac parte din categoria lectinelor si se gasesc numai la bacteriile Gram negative. Ele se leaga de receptori specifci ai membranei celulare. Fimbriile care apartin aceluiasi tip sunt formate din molecule proteice identice. Diferitele variante fimbriale se deosebesc prin lungimea filamentului si prin gr. mol. a monomerilor polipeptidici constitutivi.
In aceiasi categorie sunt incluse o categorie de proteine din structura membranei externe a bacteriilor Gram negative, care indeplinesc functia de adezine.
Grupul adezinelor neproteice. Cele mai comune adezine neproteice sunt polizaharidele care intra in structura unei retele de tip glicocalix, de tip capsular sau a unei structuri neorganizate de tipul stratului mucos. Ele sunt prezente la bacteriile Gram pozitive si Gram negative.
Exopolizaharidele formeaza structuri capsulare la multe bacterii patogene si au consistenta de geluri hidratate. Ca factori de virulenta, exopolizaharidele au doua roluri majore:
a) aderenta de substratul tisular. Dupa Gristina si colab. (l985), atat suprafata celulei bacteriene cat si a substratului tisular sunt polianionice. Sarcinile electrice asemanatoare determina respingerea celulei bacteriene de catre substratul tisular. Fortele de respingere sunt contracarate de fortele de atractie London-van der Waals. Ele permit realizarea unor interactiuni hidrofobe intre moleculele suprafetei bacteriene si ale celulei gazda. Exopolizaharidele bacteriene intensifica interactiunile dintre liganzii bacterieni si tisulari. Astfel, tesutul este acoperit de un biofilm, in care agregatul de celule bacteriene, inconjurat de o matrice moleculara adera strans de substrat.
b) Protectia celulei bacteriene fata de sistemele de aparare a gazdei. Capsula impiedica fagocitarea pentru ca bacteriile capsulate formeaza microcolonii si inglobarea lor de catre fagocite devine dificila. Cu cat suprafata celulei bacteriene este mai hidrofila, cu atat este mai greu fagocitata. Pe de alta parte, complementul si anticorpii nu au acces la suprafata celulei. Unele specii bacteriene produc o capsula, a carei compozitie chimica este asemanatoare componentelor tisulare. O astfel de capsula camufleaza celula bacteriana fata de sistemul imunitar. De exemplu, capsula de la Str. pyogenes si Pasteurella multocida contin acid hialuronic.
Matricea moleculara polizaharidica, cu structura fibroasa favorizeaza cresterea celulelor si stabilitatea biofilmului. Stabilitatea biofilmului asigura formarea continua de microcolonii, persistenta si extinderea infectiei cronice. Din structura sa "scapa" periodic celule bacteriene care trec in s@nge sau in lichidele tisulare adiacente, favorizand extinderea infectiei.
Alte adezine sunt acizii lipoteichoici, caracteristici bacteriilor Gram pozitive, ancorati de glicolipidele membranare. Sunt structuri moleculare filamentoase, foarte asemanatoare fimbriilor, cu rol de aderenta la substrat. La Str. pyogenes, acizii lipoteichoici au efect sinergic cu proteina M.
Adezinele neproteice sunt mai putin eficiente in medierea proceselor de aderenta bacteriana, comparativ cu adezinele proteice. Polizaharidele capsulare, in anumite cazuri favorizeaza aderenta, iar alteori au efect invers (de exemplu, polizaharidele impiedica aderenta celulelor de H. influenzae, N. meningitidis sau P. multocida tip A).
Receptori de adezine. Interactiunea dintre celula epiteliala sensibila si adezinele bacteriene este mediata de receptorii de adezine si se bazeaza pe principiul complementaritatii spatiale. Uneori, interactiunea este nespecifica, ceea ce explica faptul ca unele specii bacteriene (Salmonella, Yersinia) infecteaza atat celulele intestinale umane, cat si liniile celulare de Drosophyla. Alteori, interactiunea are un grad inalt de specificitate. De exemplu, Str. pyogenes se localizeaza la nivelul epiteliului faringian si foarte rar in tractul urinar, iar E. coli colonizeaza epiteliul intestinal si al cailor urinare, dar foarte rar pe cel respirator.
Helicobacter pylori, un patogen specific uman,
adera de celulele epiteliale ale mucoasei gastrice, dar nu de celulele
profunde ale mucoasei sau ale colonului. Pe celulele epiteliului, H. pylori recunoaste antigenul de
grup sanguin Lewis, dar probabil si alte molecule, deoarece H. pylori infecteaza si
indivizi negativi pentru acest antigen. Un factor esential pentru
colonizare este ureaza, pe care H. pylori o produce in
cantitati foarte mari. In vitro,
celulele de H. pylori
hidrolizeaza ureea, eliberand amoniul care neutralizeaza aciditatea
si permite colonizarea initiala. Amoniul generat prin hidroliza
ureii produce NH4OH care se disociaza si ionul
Receptorii de adezine ai celulelor epiteliale sunt molecule din familia integrinelor, care indeplinesc functii fiziologice importante: leaga proteine plasmatice, dar si proteine din matricea extracelulara (fibronectine, fibrinogen, laminina, colagenul de tip I etc.).
Receptorii de adezine sunt molecule ale suprafetei celulei sau se gasesc la nivelul mucopolizaharidelor care tapeteaza totdeauna epiteliile mucoase.
Din punct de vedere chimic, receptorii de adezine sunt glicoconjugate: glicoproteine ale membranei celulare sau ale stratului mucopolizaharidic si glicolipide din structura membranei.
Glicoproteinele si glicolipidele proemina la suprafata membranei celulare si expun la exterior catene oligozaharidice mai mult sau mai putin complexe, cu rol de receptori de adezine.
Glicoproteinele din stratul mucopolizaharidic sunt formate din catene polipeptidice legate prin punti S-S. Catena oligozaharidica este formata din N-acetil-galactozamina, N-acetil-glucozamina, fucoza, galactoza si acid sialic. Glicoproteinele din mucus se gasesc si in secretii (salivara, digestive).
Glicoproteinele membranare se deosebesc de cele din mucus: N-acetil-galactozamina lipseste, locul ei fiind luat de manoza si de acizii uronici. Catenele glicoproteice ale mucopolizaharidelor si ale suprafetei celulare sunt asemanatoare cu cele ce se gasesc pe suprafata eritrocitelor si formeaza antigenele de grup sanguin. Asa se explica proprietatile hemaglutinante pe care le poseda anumite adezine bacteriene, daca receptorul lor celular se gaseste pe hematii.
Glicoconjugatele pot avea rol de receptori pentru toxine. De exemplu, glicolipidul GM1, prin componenta sa oligozaharidica este receptor specific al toxinei holerice.
Detectarea localizarii receptorilor glicoproteici si glicolipidici pe suprafata celulei este posibila prin utilizarea lectinelor. Lectinele sunt glicoproteine ce se combina specific cu un rest oligozaharidic. Lectinele cuplate cu un derivat fluorescent sunt utilizate pentru a studia repartitia receptorilor suprafetei celulare.
Modularea expresiei proprietatilor de aderenta se poate face, teoretic, pe mai multe cai:
prin blocarea receptorilor celulari cu analogi structurali ai adezinei bacteriene. De exemplu, aderenta streptococilor poate fi inhibata in vitro, daca la mediul de reactie se adauga acid lipoteichoic, analogul structural al adezinelor acestor bacterii;
prin utilizarea analogilor structurali ai receptorilor celulari. D-manoza, adaugata in mediul de reactie inhiba aderenta S. typhimurium si Sh. flexneri de eritrocitele de cobai. Bacteriile poarta fimbrii de tip I, ai caror receptori celulari sunt glicoproteine cu manoza;
blocarea partiala sau totala a adezinei cu anticorpi specifici este considerata ca o modalitate eficienta de protectie. Astfel, starea imunitara postvaccinala sau consecutiva unei infectii, asigura o protectie buna fata de o infectie ulterioara cu acelasi agent sau cu alti agenti care au adezine comune;
utilizarea antibioticelor la doze subinhibitorii este o modalitate de a modifica expresia adezinelor. Dozele subinhibitorii blocheaza partial sau total, biosinteza unor constituienti ai suprafetei celulare, antrenand si modificari ale anumitor proprietati, inclusiv a celor de aderenta.
Receptorii celulari de adezine se modifica, atat cantitativ cat si calitativ.
Modificarile receptorilor celulei gazda pot favoriza aderenta si invazia tisulara. Un exemplu este infectia severa produsa de Str. pneumoniae, care determina infectii clinice(pneumonie, otita medie, septicemie). Sensibilitatea la infectie creste foarte mult consecutiv unei infectii virale a tractului respirator. Citochinele gazdei activeaza receptorul pentru PAF(platelet activating factor), pe suprafata celulelor epiteliului pulmonar. Pneumococii ambelor tipuri de colonii(transparente si opace) adera de suprafata celulei pulmonare, dar cei ce produc colonii transparente invadeaza circulatia sistemica prin capilarele pulmonare ori prin vasele limfatice. Asa se explica predispozitia pacientilor infectati cu virus, la complicatiile consecutive pneumoniei pneumococice(meningita, septicemie).
Interactia unui patogen cu celula gazda nu este mediata totdeauna de receptori celulari. Adeseori, bacteriile patogene activeaza caile de semnalizare ale celulei, direct prin componentele celulare sau prin intermediul citochinelor inflamatorii.
Sideroforii ca factori de virulenta
Sideroforii sunt molecule mici, difuzibile, secretate de multe microorganisme, care au in structura lor, grupari caracteristice de tipul catechol, fenolat sau hidroxamat. Structura moleculara caracteristica, definita functional ca o "cusca moleculara", are rolul de a lega Fe3+ (feric).
La bacteriile Gram pozitive, complexele Fe-siderofor sunt recunoscute selectiv de receptori membranari si transportate in celula cu consumul ATP. {n celula, complexul Fe-siderofor se disociaza prin hidroliza sideroforului si/reducerea ionului feric la Fe2+(feros), care este depozitat in bacterioferitina, utilizata ca un cofactor in cateva procese celulare vitale. Fe este legat in centrul sideroforilor cu o afinitate echivalenta celei de legare cu transferina. Sideroforii pot prelua ionii de Fe chiar din compusi minerali, ca hematita.
Multiplicarea bacteriilor in tesuturile gazdei este limitata de mecanismele de aparare ale gazdei si este conditionata de prezenta obligatorie a Fe2+ (forma accesibila metabolismului). Fe este necesar desfasurarii unor variate reactii enzimatice, esentiale pentru crestere, fiind un cofactor de sine statator sau parte a unui grup prostetic, in special de tip hem. Fe este un component esential al catenei de transport al electronilor in membrana celulara.
In mediile naturale, ca si in organismul uman si animal, Fe se gaseste in cantitati suficient de mari pentru a asigura cresterea microorganismelor, dar este in forme inaccesibile. Astfel, concentratia normala a Fe in plasma mamiferelor (mai mica de l0 mM) este satisfacatoare pentru cresterea si multiplicarea bacteriilor, dar numerosi factori il fac inaccesibil microorganismelor. Practic, Fe disponibil lipseste. Un deficit partial al Fe are efect bacteriostatic, iar deficitul major are efecte letale, prin inhibitia sintezei proteinelor. Fe anorganic este insolubil in aerobioza la pH fiziologic.
In mediile naturale, Fe este legat de agenti chelatori (compusi chimici in care un ion metalic multivalent este captat, sechestrat si legat in structura ciclica a agentului chelator). In organismul uman si animal, Fe este legat in complexe organice cu proteinele (feritina, hemosiderina, gruparea hem din hemoglobina, sau din mioglobina), cu glicoproteine extracelulare (lactoferina din lapte, saliva, lacrimi), precum si cu transferina din sange si limfa. Toate aceste molecule au o mare afinitate de asociere cu Fe si in mod normal sunt partial saturate. Capacitatea de legare a transferinei, proteina majora de transport a Fe in plasma sanguina, este ocupata in proportie de 30%. In ser, hemoglobina (Hbg) este complexata cu alte proteine ca haptoglobina, iar cantitati mici de hem pot fi legate de albumina sau de hemopexina. Fierul intracelular poate fi asociat cu Hbg, transferina sau cu lactoferina sau este depozitat sub forma feritinei.
Proteinele de legare reduc concentratia Fe3+ liber in organism, la valori foarte scazute. Nivelul Fe biodisponibil este estimat la 10-18 M, de cateva ori mai mic decat nivelul cresterii normale. Dupa administrarea parenterala a Fe la cobai, sensibilitatea la infectia cu K pneumoniae a crescut dramatic, ceea ce demonstreaza rolul aprovizionarii cu Fe asupra patogenezei bacteriilor. De aceea, serul sanguin uman este bacteriostatic.
Din aceste cauze, bacteriile si-au elaborat mecanisme adaptative prin care mobilizeaza Fe si il disponibilizeaza pentru activitatile celulei. Bacteriile cu localizare intracelulara dobandesc Fe din rezerva celulei. L. monocytogenes produce un reducator al Fe3+ la Fe2+ si astfel Fe este eliberat din glicoproteinele chelatoare intracelulare.
Fe intracelular poate fi obtinut prin liza celulei gazda, mediata de citolizine sau hemolizine. Se elibereaza Fe, hemul sau Hbg. Unele bacterii folosesc Fe din celulele care degenereaza la suprafata epiteliilor mucoase. Neisseria si Prevotella (Bacteroides) elimina proteaze specifice care degradeaza transferina si elibereaza Fe.
Cel mai raspandit mecanism prin care bacteriile isi obtin Fe in organismul gazda, este sinteza agentilor chelatori care intra in competitie cu mecanismele gazdei care sechestreaza Fe. Denumirea generala a agentilor chelatori bacterieni este cea de siderofori.
In conditiile deficitului de Fe, enterobacteriile sintetizeaza o varietate de siderofori. Grupul cel mai comun este de tip fenolat, cel mai cunoscut reprezentant fiind enterobactina (enterochelina). La E. coli, in conditiile de restrictie a Fe, translocatia Fe3+ in celula si conversia sa in forma utilizabila (Fe2+) se face prin doua mecanisme:
l) Sinteza si secretia sideroforilor. Sideroforii sunt agenti chelatori de mare afinitate ai Fe, cu gr. mol. mica (600 D), capabili sa preia competitiv Fe legat de proteinele gazdei. Complexul Fe3+ - siderofor este recunoscut de un receptor situat in membrana externa si nu poate trece prin canalele membranei celulei bacteriene datorita dimensiunilor sale. Transferul membranar al Fe3+ mediat de receptor, este asociat cu reducerea lui la forma feroasa (Fe2+), iar sideroforul ramane la nivelul receptorului.
Sideroforii se gasesc nu numai la bacteriile patogene, ci constituie mecanisme eficiente de preluare a Fe in tot regnul procariotelor. In functie de specie, ei sunt factori cu diferite grade de virulenta.
2) Un alt mecanism de preluare a Fe este propriu unor bacterii patogene care colonizeaza mucoasele (Neisseria, Bordetella). Ele nu secreta siderofori, dar preiau Fe pe o cale directa, prin contactul selectiv intre componentele membranei externe, cu proteinele care leaga Fe (transferina, lactoferina sau chiar hemoglobina). Neisseria sp. si H. influenzae utilizeaza ca surse de Fe numai transferina si lactoferina umana, ceea ce ar explica specificitatea lor pentru gazdele primate.
Fe are nu numai un rol fiziologic deosebit, dar multe bacterii il utilizeaza pentru activarea unor factori de virulenta: toxine, adezine, invazine.
b) Agresivitatea (Invazivitatea)
Puterea de invazie (agresivitatea sau invazivitatea) reprezinta capacitatea agentilor patogeni de a depasi prin mecanisme specifice, barierele epiteliale, de a patrunde in tesuturile gazdei si de a se multiplica, producand efecte patologice. Microorganismele invazive au capacitatea de a patrunde prin mijloace proprii in tesuturile gazdei sau de a stimula functia endocitara a substratului si de a-si pastra viabilitatea in mediul gazdei.
Invazia este modalitatea prin care microorganismele infectioase sparg barierele epiteliale ale gazdei. Multe bacterii patogene au capacitatea de a supravietui in interiorul celulei eucariote. Ele patrund in celule care, in mod obisnuit nu sunt fagocitare(celulele mucoase, celulele endoteliale ale vaselor sanguine). Mediul intracelular ofera protectie microorganismului, care ori se replica ori persista.
In general, organismele invazive adera de celula gazda, prin intermediul unei clase de molecule de adezine, denumite invazine, care orienteaza intrarea bacteriei in celula. Mecanismele de aderenta declanseaza sau stimuleaza semnalele celulare, care direct sau indirect usureaza intrarea bacteriei. Invazia este un eveniment activ, sustinut de functiile normale ale celulei. Suportul procesului de invazie si inglobare este citoscheletul celulei gazda.
Un numar mic de specii bacteriene par sa forteze intrarea directa in celula gazda, printr-o digestie enzimatica locala a membranei celulei gazda, dupa aderenta. De exemplu, R. prowazecki secreta fosfolipaze care produc degradarea localizata si controlata a membranei celulei gazda. Prin leziunile membranare, agentul patogen intra direct in citoplasma.
In categoria invazinelor intra o categorie de proteine asociate suprafetei celulare, dar si apendicele celulare evidentiate la microscopul electronic (fimbrii si flageli) sau cele care constituie un strat fin, inalt organizat, pe suprafata celulei. Invazinele usureaza raspandirea bacteriilor in organism, dupa fixarea si multiplicarea lor la poarta de intrare.
Flagelii ca organite de motilitate sunt in acelasi timp invazine, deoarece confera un avantaj evident celulelor bacteriene care trebuie sa traverseze stratul vascos de mucus, pentru a ajunge la celulele epiteliale ale tractului digestiv sau respirator.
Fimbriile, la bacteriile patogene, au rol de adezine bacteriene, dar in aceiasi masura ele constituie si un factor de agresivitate, deoarece confera un grad de protectie a celulei fata de factorii de aparare a gazdei.
Unele invazine produc efect necrotic sau determina alte modificari care favorizeaza colonizarea tisulara progresiva. De exemplu, unele bacterii produc hialuronidaza si alte enzime care hidrolizeaza polimerii din substanta fundamentala a tesutului conjunctiv, favorizand astfel intrarea bacteriilor in vasele sanguine si limfatice.
Enterobacteriile au mecanisme proprii de invazie. Agentii patogeni enterici strapung bariera mucoasei intestinului subtire, prin motilitate flagelara, la nivelul unor celule epiteliale specializate, denumite celule M (Microfold) care acopera placile Peyer.
c) Toxigenitatea
Toxigenitatea (toxigeneza) reprezinta capacitatea unui agent patogen de a elabora in cursul cresterii sale, una sau mai multe substante toxice. Toxigeneza este o proprietate esentiala a patogenitatii bacteriene.
Sub denumirea de toxina sunt cuprinse toate substantele toxice de provenienta biologica (sintetizate de bacterii, fungi, celule vegetale sau animale). Termenul de "toxina" deriva din cuvantul grecesc "toxicon", care inseamna otrava.
Se cunosc circa 140 de toxine proteice, din care 2/3 sunt produse de bacteriile Gram pozitive. Unele specii bacteriene produc intre 5 si l0 tipuri de toxine.
Capacitatea de a elabora toxine nu este limitata la bacteriile patogene propriu-zise. Numeroase saprobacterii produc substante toxice, uneori foarte puternice (de exemplu, toxina botulinica produsa de Cl. botulinum). Toxigenitatea bacteriana variaza in cadrul aceleiasi specii si este dependenta de conditiile de mediu.
Bacteriile patogene sintetizeaza o varietate de substante care, direct sau indirect, sunt toxice pentru celulele si tesuturile gazdei. Unele sunt secretate de celulele bacteriene in cantitati foarte mici, fiind proteine cu actiune predominant enzimatica si poarta denumirea generica de exotoxine. Altele, de natura lipopolizaharidica (LPS), care nu au actiune enzimatica si sunt biologic active la concentratii mult mai mari, apartin endotoxinelor. Se cunosc circa 140 de toxine proteice, din care 2/3 sunt produse de bacteriile Gram pozitive. Unele specii bacteriene produc intre 5 si l0 tipuri de toxine.
Capacitatea de a elabora toxine nu este limitata la bacteriile patogene propriu-zise. Numeroase saprobacterii produc substante toxice, uneori foarte puternice (de exemplu, toxina botulinica produsa de Cl. botulinum). Toxigenitatea bacteriana variaza in cadrul aceleiasi specii si este dependenta de conditiile de mediu.
Clasificarea toxinelor bacteriene
Pentru a-si exercita efectul, toxinele trebuie sa se elibereze din celule si sa se solubilizeze in umorile organismului. Dupa sinteza, toxinele pot ramane asociate permanent sau temporar cu celula sau sunt eliminate la exterior. Aceste diferente sunt dependente, in primul rand de specia producatoare, precum si de fazele succesive ale evolutiei unei populatii bacteriene: faza exponentiala, stationara sau de declin. Chiar toxinele care in faza exponentiala a culturii bacteriene sunt asociate celulei, in faza de declin se gasesc libere in mediul extracelular datorita lizei celulelor. Din aceasta cauza, studiul raportului topologic intre celula si toxina are semnificatie numai pentru faza de evolutie a culturii bacteriene. Din acest punct de vedere se disting urmatoarele categorii de toxine:
a) Toxine localizate in celula (citoplasmatice) produse de bacterii Gram negative (Sh. dysenteriae, Y. pestis, B. pertusis, E. coli etc.) si Gram pozitive (enterotoxina produsa de Cl. perfringens, streptolizina S, pneumolizina produsa de S. pneumoniae, o toxina sintetizata de Cl. difficile). Toate sunt de natura proteica. Sunt localizate in citoplasma sau sunt asociate membranei citoplasmatice si pot fi eliberate dupa indepartarea peretelui prin liza mecanica sau prin extractie chimica.
b) Toxinele constitutive ale peretelui celular, produse numai de bacteriile Gram negative si care corespund endotoxinelor clasice. Sunt situate in afara membranei citoplasmatice si fac parte din structura membranei externe a peretelui celular. Din punct de vedere chimic, endotoxinele sunt complexe glicolipidice sau glicolipoproteice. Ele nu sunt niciodata eliberate in cursul fazei de crestere exponentiala, ci numai prin dezagregarea peretelui celular.
c) Toxinele eliminate in mediul extern sau exotoxinele propriu-zise sunt de natura proteica si sunt produse mai frecvent de bacteriile Gram pozitive (toxina difterica, toxinele stafilococice, toxina de B. anthracis etc.), dar si de cele Gram negative (V. cholerae, Ps. aeruginosa). Exotoxinele indeplinesc in mod obligatoriu, 3 conditii:
- se gasesc in mediul de crestere, independente de celula care le-a produs;
- eliberarea lor in mediul extracelular nu necesita autoliza celulei care le secreta. Celula ramane viabila, continua sa creasca si sa se multiplice;
- nu se acumuleaza in celula.
Aceste criterii se aplica celulelor in faza de crestere exponentiala si stationara, deoarece in faza de declin, celulele pot prezenta fenomene de autoliza, independente de sinteza toxinei.
d) Toxinele cu localizare mixta (endocelulara si exocelulara), in functie de faza de evolutie a culturii bacteriene. In faza exponentiala, toxinele sunt secretate partial in mediu, dar o fractie semnificativa ramane in interiorul celulei si este eliberata prin autoliza (toxinele tetanica, botulinica).
Mult timp s-a considerat ca exotoxinele sunt de natura proteica, iar endotoxinele - de natura lipopolizaharidica. Aceasta diferentiere nu este neta, deoarece unele toxine proteice apartin categoriei endotoxinelor, fiindca raman asociate celulei producatoare si invers, uneori, toxinele de natura lipopolizaharidica sunt eliminate la exterior ca exotoxine. De aceea, clasificarea toxinelor trebuie sa se faca dupa criteriul compozitiei chimice. Din acest punct de vedere, majoritatea sunt de natura proteica, iar majoritatea endotoxinelor sunt de natura lipopolizaharidica.
Raynaud si Alouf (1970) au clasificat toxinele in raport cu compozitia chimica si cu localizarea celulara.
Grupa I cuprinde toxinele proteice intracitoplasmatice ale bacteriilor Gram negative, eliberate dupa ruperea mecanica, enzimatica, prin autoliza invelisurilor sau prin extractie chimica. Sunt sintetizate de B. pertussis, Sh dysenteriae (neurotoxina), Y. pestis.
Grupa a II-a cuprinde toxinele lipopolizaharidice sau glico-lipo-peptidice (endotoxine), componente structurale ale peretelui celular Gram negativ. Cele mai tipice endotoxine se gasesc la enterobacterii.
Grupa a III-a cuprinde exotoxinele proteice propriu-zise, produse de V. cholerae, Cl. tetani, C. diphteriae.
Grupa a IV-a cuprinde toxinele proteice cu localizare intra- si extracelulara, in cursul fazei logaritmice de crestere: toxinele produse de Cl. tetani, Cl. botulinum, Cl. oedematiens, Cl. sordelii.
Din punctul de vedere exclusiv al compozitiei chimice, se disting:
1) Toxine proteice, reprezentate de exotoxinele produse de bacteriile Gram pozitive si de toxinele proteice intracelulare ale unor bacterii Gram negative: B. pertussis, Y. pestis, Sh. dysenteriae (neurotoxina).
2) Toxinele glico-lipo-polipeptidice, corespunzatoare endotoxinelor.
In raport cu tropismul lor se disting neurotoxine, enterotoxine, cardiotoxine, cu efecte limitate la tesutul respectiv sau toxine pantrope, cu efecte asupra multor categorii de tesuturi.
Toxicitatea exotoxinelor
Unele toxine au efect toxic foarte puternic. De exemplu, toxina botulinica de tip D este de 3 milioane de ori mai puternica decat stricnina, luata ca etalon, iar toxinele tetanica, neurotoxina de Sh. dysenteriae si cea botulinica de tip A sau B, de un milion de ori.
Efectul toxic este variabil nu numai in functie de specia bacteriana producatoare, ci si de tulpina, chiar in conditiile unui mediu optim. Adeseori, toxigeneza se atenueaza sau chiar dispare, in subculturi repetate.
Efectul toxinelor este variabil in functie de natura, varsta, greutatea, sexul, linia genetica a animalelor pe care sunt testate, ca si de calea de administrare a toxinei.
Toxicitatea diferitelor probe ale aceleiasi toxine poate prezenta variatii foarte importante: de exemplu, unele probe de toxina botulinica sunt de 6000 de ori mai toxice pentru cobai decat pentru soarece, iar altele numai de 3 ori.
Exotoxinele au, uneori, un rol determinant in patogeneza unor agenti infectiosi, datorita potentialului lor toxic foarte inalt. De exemplu, toxinele purificate reproduc in general, manifestarile clinice ale infectiei cu V. cholerae, C. tetani sau C. diphteriae. Dar la majoritatea agentilor patogeni, exotoxinele actioneaza sinergic cu alte componente celulare pentru a produce efectele potentiale asupra gazdei. Sinteza si secretia toxinelor este reglata de alti factori de virulenta. De exemplu, capacitatea de aderenta creeaza un contact strans intre agentul patogen si celula sensibila, ceea ce permite transferul toxinei si exprimarea intregului potential toxic. Desi toxina holerica poate reproduce multe din simptomele procesului infectios, pentru exprimarea integrala a virulentei bacteriene este necesara adezina care leaga celula bacteriana de celula mucoasei intestinale. Alteori, toxina are alte functii suplimentare celei de toxicitate, care amplifica virulenta: toxina de B. pertusis are si rolul de proteina de aderenta a bacteriei, de celulele mamaliene.
Receptori celulari pentru toxine
Interactiunea toxinelor cu celulele sensibile, asemenea altor biomolecule (hormoni, antigene, lectine, fatori reglatori etc.) este mediata de molecule specifice denumite receptori, situate in stratul extern al membranei citoplasmatice. Celulele sensibile nu au receptori specifici pentru toxine. Toxinele imprumuta receptorii, care in mod obisnuit au rolul de a ingloba molecule utile metabolismului celular.
Se cunosc receptorii prin intermediul carora, cateva toxine interactioneaza cu suprafata celulei. De exemplu, receptorul celular pentru streptolizina O (SLO), pentru listeriolizina (LLO) si pentru pneumolizina (PLO) este colesterolul. Aceste toxine se fixeaza pe suprafata celulei prin intermediul colesterolului membranar si nu depind de alti receptori de suprafata. De aceea, ele pot sa lizeze membranele, teoretic, ale oricarei celule animale.
Toate toxinele acestui grup sunt alcatuite dintr-o singura catena polipeptidica, a carei lungime variaza de la 471 aminoacizi (pentru pneumolizina) pana la 571 pentru streptolizina O. Variatia lungimii se datoreaza in intregime secventelor localizate la capatul N-terminal, a caror functie ramane necunoscuta. Cea mai lunga secventa, cu omologie aproape perfecta la toxinele grupului este de 11 aminoacizi si este bogata in triptofan.
Secventa comuna pentru toate acete molecule si esentiala pentru activitatea citolitica corespunde celei mai mici secvente - a pneumolizinei. Pneumolizina se deosebeste de celelalte toxine ale grupului, prin absenta peptidului semnal secretor si din aceasta cauza este eliberata numai prin liza celulelor de Streptococcus pneumoniae.
Aceste toxine nu se leaga de membranele care nu contin colesterol sau un compus inrudit cu colesterolul. Interactiunea cu colesterolul se produce in absenta altor lipide. Toxinele interactioneaza chiar cu sterolul pur in solutie sau in suspensie si rezultatul este inhibitia activitasii litice.
Alte cateva toxine au ca receptori moleculele de gangliozide. Acestea sunt glicolipide, mai abundente in neuroni, alcatuite dintr-o componenta oligozaharidica legata de un ceramid (acid stearic si sfingozina). Gangliozidele difera intre ele, prin numarul si secventa resturilor glucidice componente, in special a acidului N-acetil neuraminic (NANA) sau de acid sialic.
Toxina tetanica se leaga de gangliozide, in special de di- si tri-sialogangliozide, care contin doua si respectiv trei resturi de acid sialic, atasate de galactoza. Intre toxina tetanica si hormonul tirostimulator (TSH) exista o competitie de legare strict reciproca, pe membrana celulelor tiroidiene. Astfel s-a dedus ca receptorul neuronal pentru toxina tetanica este asemanator cu receptorul celulei tiroidiene pentru TSH.
Receptorul celular pentru toxina holerica, ca si pentru toxina termolabila de E. coli, foarte asemanatoare toxinei holerice este gangliozidul GM1. S-a evidentiat o corelatie directa intre continutul membranar in GM1 si sensibilitatea tisulara la toxina.
Receptorul celular al toxinei de B. pertussis pare a fi tot o gangliozida care contine acid sialic, iar receptorul toxinei difterice ar fi o glicoproteina.
In raport cu localizarea tintei lor moleculare se disting doua nivele de actiune a toxinelor:
cele care au tinta finala la nivelul membranei;
toxine a caror tinta finala este citoplasmatica.
Tinta finala este structura a carei interactiune cu toxina produce efectul toxic, in timp ce receptorul are uneori rolul de tinta intermediara, desi alteori este chiar tinta finala.
Toxinele care actioneaza la nivelul membranei sunt active prin modificari structurale pe care le induc in membrana citoplasmatica, urmate de ruperea acesteia. Consecinta este citoliza ori moartea celulei. Toxinele care consecutiv actiunii lor produc dezorganizarea membranei se numesc citolizine (respectiv hemolizine, daca celula tinta este eritrocitul).
Toxinele a caror tinta finala este intracelulara, mai intai traverseaza membrana si ajung in citoplasma.
Moleculele suprafetei eritrocitului, cu rol de receptor pentru toxinele bacteriene, sunt glicolipidele. Legarea toxinelor de glicolipide pare a fi mai avantajoasa decat legarea de glicoproteine, pentru interactia cu membrana, care este esentiala pentru actiunea toxica.
Patrunderea moleculelor de toxina in celula Toxinele bacteriene a caror tinta este intracelulara sunt molecule bifunctionale, ca si toxinele vegetale (ricina, abrina), bacteriocinele sau hormonii glicoproteici. Toate aceste categorii de molecule sunt alcatuite dupa acelasi model functional, fiind monomere (de exemplu, toxina difterica, exotoxina de Ps. aeruginosa) sau dimere (de exemplu, enterotoxina holerica si cea termostabila de E. coli). Cele monocatenare au o secventa COOH-terminala, prin care se leaga la nivelul receptorului si o secventa NH2-terminala, care patrunde in citoplasma si interactioneaza cu tinta intracelulara. Pentru toxinele dublu catenare, functiile de legare B (Binding) si de activitate propriu-zisa A (Activity) sunt realizate separat de fiecare catena.
Patrunderea moleculei de toxina in celula este un proces complex pentru majoritatea toxinelor cu actiune intracelulara si se face prin unul din urmatoarele mecanisme:
- endocitoza mediata de receptori;
- pinocitoza nespecifica in faza lichida;
- transferul direct al moleculei prin membrana citoplasmatica.
Inglobarea prin mecanismul endocitozei mediate de receptori confera specificitate si eficienta actiunii moleculelor mari. Receptorii celulari glicoproteici, dupa interactiunea cu moleculele de toxina, sufera o dinamica accentuata. In mod normal, moleculele cu rol de receptor sunt uniform distribuite in planul membranei sau sunt concentrate in teritorii specializate denumite zone tapetate cu clatrina* (coated pits), un invelis de natura proteica pe fata citoplasmatica a membranei. Dupa legarea moleculei de toxina, complexele receptor-toxina se aglomereaza in zonele tapetate cu clatrina, situate la baza microvilozitatilor. Zonele respective se intruzeaza si formeaza vezicule acoperite cu clatrina, cu rol de transport. In citoplasma, invelisul de clatrina se dezorganizeaza si receptorii celulari deveniti disponibili sunt reciclati spre suprafata celulei, iar molecula de toxina este eliberata spre un situs intracelular specific.
*Clatrina este o proteina mare, oligomerica, ce formeaza o retea pe suprafata interna a membranei plasmatice, favorizand intruzia membranei si formarea unei vezicule tapetata cu reteaua moleculara, care ulterior poate fuziona cu alte organite celulare.
Mecanisme generale de actiune a toxinelor
Efectul biologic al exotoxinelor este specific si este datorat afinitatii lor caracteristice pentru anumite celule-tinta ale organismului.
Unele toxine (tetanica, difterica, botulinica, holerica, eritrogena) au un rol determinant in patogenitatea bacteriana. Primele patru enumerate mai sus, sunt factori unici ai patogenitatii pentru bacteriile producatoare. In cazul holerei, gravitatea infectiei este consecinta efectelor fiziopatologice nespecifice ale toxinei asupra mucoasei intestinale ce constau in pierderea apei si sarurilor minerale la acest nivel.
Eritemul specific scarlatinos, produs de Streptococcus pyogenes se datoreaza efectului primar al toxinei eritrogene streptococice.
Enterotoxinele stafilococice si enterotoxina de Cl. perfringens produc efecte la nivelul mucoasei intestinale, ca rezultat al intoxicatiei alimentare.
Toxina produsa de Sh. dysenteriae este cauza dizenteriei, deoarece enterocitele si celulele endoteliale ale vaselor mici se lizeaza sub actiunea toxinei, rezultatul fiind ulcerarea mucoasei intestinale.
Exotoxina de B. anthracis produce un edem local si hemoragie, iar in cazul septicemiei, efectul letal pare a fi datorat neurotropismului sau, cu actiune in special asupra centrului bulbar al respiratiei.
Uneori, toxine foarte asemanatoare, cauzeaza boli foarte diferite. De exemplu, toxina botulinica este ingerata oral si produce paralizia flasca a muschilor striati, prin blocarea transmiterii potentialului la nivelul placii motorii. Toxina tetanica, produsa in ranile profunde infectate cu Cl. tetani, produce paralizia spastica, prin SNC.
Efectele biologice ale toxinelor se produc la nivelul diferitelor structuri celulare. De exemplu, hemolizinele si leucocidinele actioneaza la nivelul membranei citoplasmatice, iar alte toxine sunt active asupra organitelor, asupra catenei transportoare de electroni s.a.m.d.
Toxinele perturba functiile celulei gazda: influenteaza caile de transducere a semnalelor, rearanjeaza citoscheletul si traficul vacuolar.
Toxinele bacteriilor Gram pozitive, in general, nu necesita activare. Multe toxine ale bacteriilor Gram negative sunt sintetizate in forma inactiva si necesita o etapa de prelucrare, care consta in clivajul proteolitic, pentru a genera forma activa.
Toxinele cu actiune enzimatica (toxina Shiga, toxina holerica, toxina pertusica, toxina difterica si exotoxina A de Ps. aeruginosa) sunt clivate proteolitic pentru a produce fragmentul A catalitic activ.
Multe toxine neenzimatice care se insera in membrana celulei eucariote, necesita clivajul proteolitic pentru a permite oligomerizarea si formarea porilor: hemolizina de V. cholerae El Tor este clivata N-terminal, iar aerolizina, toxina α de C. septicum si citotoxina de Ps. aeruginosa sunt clivate C-terminal. Hemolizina de E. coli, formatoare de pori, reprezinta o clasa unica de toxine ce necesita prelucrarea posttraducere pentru activare.
Unele toxine au un plan structural si mecanisme de actiune comune, ceea ce permite gruparea lor in cateva familii.
Toxine cu activitate ADP-ribozilanta Multe exotoxine au o structura de tip A-B, adica sunt formate din doua componente: subunitatea B (binding), care mediaza legarea moleculei si patrunderea ei in celula sensibila; subunitatea A (activity), care determina efectul enzimatic (toxic) specific. Domeniul B are cea mai larga variatie, probabil pentru a conferi specificitate de legare de substratul celular sensibil.
Activitatea enzimatica a subunitatii A produce efecte foarte variabile, de la cea ADP-ribozilanta (toxinele holerica, pertusica, difterica, toxina termolabila de E. coli, exotoxina A de Pseudomonas, ADP-ribozil-transferazele - C2 si C3 produse de Cl. botulinum), pana la actiune proteolitica (toxinele tetanica si botulinica). Toxinele ADP-ribozilante au ca situs comun de legare NAD-ul si transfera ADP-riboza de la NAD si o leaga covalent la proteine ale celulei eucariote, dupa reactia globala:
NAD+ + Proteina tinta ----- ADP-riboza- Proteina + Nicotinamida + H+.
Unele dintre aceste exotoxine se asociaza cu proteine cu rol important in fiziologia celulara, care au capacitatea de a lega nucleotidele si produc ADP-ribozilarea (fig. 163):
- unor proteine heterotrimerice care leaga GTP;
- unor proteine mici care leaga GTP;
- actinei;
- altor proteine ale celulei eucariote, care nu au fost inca identificate.
Exotoxinele ADP-ribozilante au trei tipuri de organizare A-B:
- proteine polipeptidice unice, cu componentele A-B legate covalent;
- complexe cu componente A-B localizate pe proteine separate, legate prin interactiuni necovalente;
- complexe multiproteice cu componente A-B localizate pe doua proteine separate.
Pentru unele exotoxine ADP-ribozilante, nu s-a evidentiat organizarea structurala de tip A-B.
Mecanismele activarii in vitro a exotoxinelor ADP-ribozilante cu organizare de tip A-B sunt:
- proteoliza partiala si generarea peptidului A activ cu activitate catalitica;
- reducerea legaturilor S-S;
- activarea alosterica de catre nucleotide sau de catre proteinele accesorii ale celulei eucariote.
Modelul de actiune ADP-ribozilant s-a elaborat pe baza proprietatilor moleculare ale toxinei difterice,
exotoxinei A de Ps. aeruginosa si enterotoxinei termolabile de E. coli.
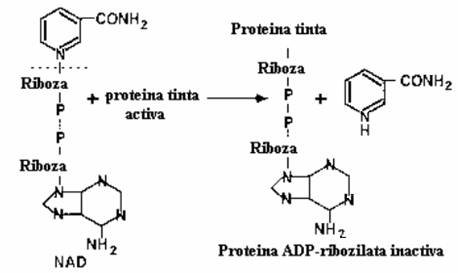
Fig. 163. Reprezentarea reactiei de ADP-ribozilare
Toxina difterica si exotoxina A de Ps. aeruginosa produc ADP-ribozilarea proteinelor mari care leaga GTP-ul, cum ar fi factorul de elongare al catenei polipeptidice EF-2, la resturi de acid glutamic diferite (148 si respectiv 553), dar omologe din punct de vedere functional, deoarece ambele leaga inelul nicotinamidei din NAD printr-un mecanism dependent de UV.
Proteinele familiei Rho leaga GTP si mediaza polimerizarea actinei celulare, avand rol in organizarea citoscheletului, in formarea veziculelor de transport, in cresterea si diviziunea celulei si desfasurarea ciclului celular, participa la procesele de transductie a semnalelor, sunt implicate in endocitoza mediata de receptor si secretie, controleaza transcrierea, apoptoza si transformarea celulara si sunt tinta actiunii toxinelor bacteriene.
Proteinele Rho sunt GTP-aze mici, controlate prin ciclul GTP-azei: sunt inactive in forma legata
de GDP, dar se activeaza dupa trecerea GDP in GTP. Invers, hidroliza GTP legat, inactiveaza proteinele.
Toxine proteolitice
Toxinele cu structura asemanatoare pot genera manifestari clinice foarte diferite. De exemplu, toxina botulinica ingerata odata cu alimentele, produce o paralizie flasca datorata actiunii la nivelul terminatiilor nervoase periferice, iar toxina tetanica, eliberata in ranile profunde contaminate cu C. tetani produce paralizia spastica prin intermediul sistemului nervos central. Ambele toxine actioneaza prin acelasi mecanism: blocarea eliberarii mediatorilor sinaptici prin clivarea sinaptobrevinelor, proteine componente ale veziculei sinaptice. Toxina tetanica blocheaza eliberarea mediatorilor glicina si GABA, in timp ce toxina botulinica blocheaza eliberarea acetilcolinei. Ambele proteine sunt metaloendoproteaze si se aseamana prin domeniile care leaga Zn. Modul de patrundere a toxinelor in organism determina manifestari clinice distincte, desi ele sunt foarte asemanatoare.
Toxine
formatoare de
Toxinele SLO, LLO, PLO apartin familiei toxinelor activate de gruparea thiol (Tabel ). Capacitatea lor de a forma pori depinde de o secventa conservata de aminoacizi la capatul C terminal, care contine resturi de Trp, esentiale pentru formarea porilor si un rest de Cys, care le face intolerante la conditiile oxidante.
Toxinele thiol -activate recunosc colesterolul din membrana celulelor eucariote si induc formarea unor pori de dimensiuni foarte mari (pana la 30 nm), inelari sau in forma de arc (acesti pori sunt de dimenisuni mai mici), alcatuiti din cca 50 de monomeri de toxina. Cantitatea de colesterol membranar trebuie sa depaseasca cu cateva ordine de marime cantitatea de toxina, pentru a avea loc o interactie stabila, probabil din cauza unor restrictii sterice sau termodinamice. Colesterolul nu are doar rol de receptor pentru moleculele de toxina, ci mediaza si oligomerizarea monomerilor de toxina, actionand probabil ca un efector alosteric.
Toxinele thiol-activate sunt alcatuite din 4 domenii, dintre care domeniul 4 este implicat in legarea de receptor (colesterol). Legarea initiala de colesterol este total reversibila, se realizeaza foarte rapid si independent de temperatura. Dupa legarea initiala, domeniile 1 si 3 implicate in oligomerizare sufera modificari alosterice care conduc initial la aparitia unui pre-por metastabil, care ulterior este stabilizat prin forte de tensiune superficiala cu aparitia porului stabil, in forma de inel. Porii incompleti, in forma de arc, permit de asemenea insertia moleculelor de toxina in membrana.
Rolul biologic major al acestor toxine este de a permite patrunderea in interiorul celulei eucariote a altor toxine sau enzime bacteriene, prin porii de dimensiuni mari. In cazul bacteriilor intracelulare, aceste toxine (ex. listeriolizina) permit propagarea infectiei de la o celula la alta.
Toxinele thiol-activate se gasesc la bacterii care au un mod de viata asemanator. Listeria si Streptococcus produc infectii invazive, iar toxinele lor sunt determinanti de patogenitate. Aceste bacterii produc enzime depolimerizante (proteaze, nucleaze) si sunt secretate concomitent cu toxinele. Deoarece porii formati de toxinele activate de thiol sunt mari, ei constituie calea de acces a acestor enzime in interiorul celulei. Toxina si enzimele coopereaza pentru degradarea celulelor animale, in beneficiul agentului patogen.
In cazul infectiilor clostridiene - gangrena gazoasa sau tetanosul, cele doua moduri de viata prezinta convergente evidente: toxinele pot fi implicate atat in stadiul initial al lezarii tesutului, necesar pentru realizarea unui mediu anaerob, cat si in stadiul tardiv (postmortem), caracterizat prin distrugere tisulara rapida si masiva.
Listeriolizina este produsa intracelular si se abate de la conditiile de actiune ale altor lizine, prin necesarul unui pH acid optim pentru actiunea ei. Mediul acid este creat in fagosom, din care Listeria se elibereaza singura prin secretia toxinei.
Unele toxine functioneaza prin insertia in membrana celulei sensibile. Rezultatul este formarea unui por sau canal, care duce la liza celulei prin mecanisme osmotice. O astfel de familie este formata de toxinele RTX (repetead toxins - denumire datorata repetarii unei secvente de 9 aminoacizi in fiecare toxina), produse de bacteriile Gram negative. Toxinele au fost grupate pe baza efectelor toxice si litice asupra celulelor gazda ale mamiferelor. Prototipul acestei familii este hemolizina de E. coli.
Hemolizina de E. coli. Hemolizina A (Hly A) de E. coli este un factor important al virulentei in infectiile extraintestinale, asa cum sunt cele ale tractului respirator superior, produse de E. coli. Este reprezentantul unei familii de toxine bacteriene care necesita modificarea posttraducere pentru dobandirea activitatii biologice.
Hly A face parte dintr-o familie care mai cuprinde leucotoxina de Y. haemolytica, hemolizinele si leucotoxinele de Actinobacillus actinomycetemcomitans, toxina bifunctionala (adenilat-ciclaza/ hemolizina) de B. pertussis si hemolizinele de P. vulgaris, Morganella morganii si Moraxella bovis, fata de care exista o identitate a secventei aminoacizilor in proportie de 30-75%. Toate se matureaza dupa sinteza si au un domeniu C-terminal care leaga Ca2+, format din secventele nonapeptidice repetitive bogate in glicina acida, ceea ce a dus la denumirea RTX. Toate se exporta din celula pe calea sistemului de secretie de tip I. Modificarea post-traducere este o caracteristica unica a acestor toxine, dar legarea Ca2+ si secretia de tip I sunt comune si altor proteine bacteriene.
Cand leaga Ca2+, secventele repetitive RTX formeaza scurte lanturi -pliate, organizate intr-o structura neobisnuita de suprahelice-
Legarea Ca2+ este o necesitate absoluta pentru o activitate citotoxica si se produce dupa exportul proteinei. Nivelul intracelular al Ca2+ este prea mic (0,1 uM) pentru a activa Hly A.
Toate sunt citotoxice pentru diferite tipuri de celule nucleate. Hemolizina de E. coli lizeaza in cateva minute, eritrocitele diferitelor specii: soarece, iepure, berbec, bovine, cal, om.
Hly A are spectru larg de actiune: lizeaza eritrocitele, granulocitele, monocitele, celulele endoteliale, celulele epiteliale renale de soarece, rumegatoare si primate. Absenta receptorilor specifici poate sa explice spectrul larg al actiunii Hly A.
Eritrocitele expuse actiunii Hly A de E. coli sufera schimbari majore ale citoscheletului care se exteriorizeaza prin formarea proiectiilor de suprafata. Liza eritrocitelor poate sa semnifice eliberarea Fe, iar liza leucocitelor poate sa aiba semnificatia unui factor de virulenta, care impiedica fagocitoza.
Unele citolizine sunt fosfolipaze: LLO de L. monocytogenes, citolizinele produse de Cl. perfringens si Ps. aeruginosa sunt fosfolipaze C, iar cea de Corynebacterium pseudotuberculosis este o fosfolipaza D.
Fosfolipazele bacteriene cuprind un grup heterogen de proteine-enzime, care produc o varietate de efecte in vivo si in vitro, de la alterari celulare minore in structura si functia membranei citoplasmatice, pana la efectul letal.
Listeriolozina O (LLO) produsa de L. monocytogenes, este singura toxina activata de gruparea tiol, a carei activitate optima este la pH-ul acid din vacuola intracelulara. pH-ul acid este declansator al lizei vacuolei.
LLO este singura toxina-enzima din familia citolizinelor, produsa de un patogen intracelular, care are rolul de a liza fagosomul. Acidifierea continutului vacuolar declanseaza efectul litic al LLO, limitat la membrana fagosomului.
LLO este reprezentata de doua fosfolipaze C, cu specificitate pentru fosfatidil-inozitol si fosfatidil-colina.
Iesirea agentilor patogeni de Rickettsia din vacuola celulei este mediata de fosfolipaze, care fac parte din aceiasi familie a citolizinelor.
Trypanosma cruzi invadeaza celulele, cu formarea vacuolelor acide intracelulare si produce o toxina activa la pH 5,5, in stadiile intracelulare ale ciclului. Blocarea acidificarii reduce capacitatea T. cruzi de a liza vacuola si de a trece in citosol.
Toxinele termostabile produse de E. coli, Yersinia, Citrobacter freundii, Vibrio mimicus actioneaza prin acelasi mecanism: activarea guanilat-ciclazei.
Enterotoxinele sunt proteine secretate care se leaga de un receptor celular, intra in celula si produc cresterea nivelului AMPc.
O mare parte dintre enterotoxinele diareice se leaga la receptorii de natura glicosfingolipidica. Unele dintre aceste toxine sunt capabile sa formeze canale permeabile pentru ionii de Ca, care patrund in celulele epiteliului intestinal si actioneaza ca mesager secundar care moduleaza procesele de transport ionic. Toxinele care nu formeaza astfel de canale, determina, in urma interactiuii cu receptorii specifici, eliberarea altor mesageri secundari inductori ai secretii ionilor de Cl, care la randul lor antreneaza secretia ionilor de Na si a apei, ca si a altor anioni, ceea ce conduce la acumularea apei in lumenul inestinal, la dezechilibrarea balantei hidro-electrolitice si generarea diareii secretorii. Activitatea enterotoxinelor se poate evidentia in vitro prin alungirea celulelor CHO (Chinese hamster ovary), rotunjirea celulelor Y-1 (mouse adrenal tumor cells) sau prin determinarea AMPc in celulele expuse actiunii toxinei. O metoda independenta de celulele cultivate este ELISA (enzyme-linked imunosorbent assay). Receptorii specifici (ex. gangliozidele GM1) sunt fixate pe faza solida pentru a lega enterotoxina, care poate fi detectata cu antiserul specific. In varianta alternativa a metodei ELISA sandwich, faza solida consta din fragmente F (ab') 2 ale anticorpilor anti-toxina. Ambele metode au sensibilitate si specificitate inalte. In vivo, pentru detectarea activitatii enterotoxinelor se foloseste testul ansei ileale de iepure sau de sobolan.
Citotoxinele sunt definite ca proteine care omoara celulele tinta. Ele pot actiona intracelular sau la nivelul membranei si formeaza pori. Citotoxinele cu actiune intracelulara se leaga la nivelul receptorilor membranari si inainte de a ajunge in citoplasma sunt prelucrate.
Mecanismele de actiune ale citotoxinelor sunt diferite:
- inhibitia sintezei proteinelor celulare
- inhibitia formarii filamentelor de actina
- formarea porilor membranari.
Formarea porilor in structura membranei tinta este un mecanism major de actiune a citotoxinelor. Astfel de citotoxine se pot detecta prin activitatea lor litica asupra eritrocitelor, motiv pentru care se mai numesc hemolizine. Liza eritrocitelor poate fi un mecanism de dobandire a Fe.
Formarea porilor induce un set de larg de reactii secundare in celulele nucleate: eliberarea citokinelor, disfunctia citoscheletului, sinteza mediatorilor lipidici. Liza leucocitelor poate determina scaderea reactivitatii imunitare.
Comparativ cu alte substante toxice, exotoxinele bacteriene au o toxicitate foarte inalta, atat in stare bruta sub forma filtratelor de cultura, dar mai ales in forma purificata. Astfel, toxina botulinica de tip D este de trei milioane de ori mai puternica decat stricnina luata ca etalon. Efectul toxic este variabil nu numai in functie de specia bacteriana producatoare, ci si de tulpina, chiar pe medii optime. In multe cazuri, toxigeneza se atenueaza sau chiar dispare prin subcultivare repetata.
In general, efectul toxinelor este variabil in functie de natura, varsta, greutatea, sexul si linia genetica a animalelor pe care sunt testate, ca si de tehnica si calea de administrare folosite.
Datele referitoare la toxigeneza se refera la administrarea lor parenterala, deoarece cu exceptia enterotoxinelor, toxinele uzuale sunt distruse de modificarile de pH sau de activitatea enzimelor digestive.
Potentialul toxic al toxinelor este ilustrat de date teoretice:
1) toxina botulinica de tip A purificata si cristalizata ar contine intr-un miligram, 1 000 000 DLM (doze limite mortale) pentru un kg corp cobai, ceea ce inseamna ca 1 mg de toxina poate sa omoare l 200 tone de cobai sau doua milioane de soareci;
2) 200 g neurotoxine (botulinica sau tetanica) ar putea omori intreaga populatie a globului.
Potentialul toxic al toxinelor bacteriene este dependent de calea de administrare. Dozele letale pentru caile respiratorie si digestiva sunt de l00 - l000 de ori mai mari decat pentru caile intravenoasa, intramusculara, intraperitoneala sau subcutanata. Pielea intacta este o bariera eficienta fata de toxine.
Determinismul genetic al sintezei toxinelor. Sinteza toxinelor bacteriene este codificata de gena tox, a carei localizare este cromosomala, plasmidiala sau fagica. La V. cholerae, subunitatile toxinei holerice sunt codificate de gene cromosomale, asociate cu alte gene ce codifica alti factori de virulenta. Astfel de grupe de gene de virulenta se numesc insule de patogenitate. Secventa lor difera mult de a altor gene cromosomale, ceea ce denota ca ele sunt achizitii relativ recente in evolutie. Ar putea sa aiba originea prin integrarea unui element genetic exogen - fagul.
La Cl. botulinum, gena tox, cu situs cromosomal, prezinta mai multe alele, care codifica sinteza a 7 serotipuri de toxina (notate A --- G), care nu dau reactie incrucisata semnificativa cu anticorpii obtinuti fata de una dintre ele.
Enterotoxinele si hemolizinele de E. coli, toxina dermo-exfoliativa stafilococica sunt codificate de gene plasmidiale.
Gena tox, codificatoare a toxinei difterice este localizata in genomul unor bacteriofagi ADN (β, P, 1, W). Activitatea ei este controlata de o gena situata pe cromosomul bacterian.
Fagii purtatori ai genei tox se gasesc in celulele de C. diphteriae in stare integrata (profag), de fag virulent replicativ sau sub forma de replicon autonom represat. Este cunoscut modul de functionare a genei tox, purtata de fagul β, ce se integreaza ca profag. Ea se gaseste aproape de situsul de insertie a genomului fagic in cromosomul bacterian, dar are propriul sau promotor si se poate exprima independent de alte gene fagice, dar poate fi represata fara sa afecteze biosinteza altor proteine fagice. Aceasta gena nu pare a fi esentiala pentru genomul fagc. Modificarea ei mutationala nu influenteaza ciclul de multiplicare a fagului. Se considera ca gena tox a acestui fag este de origine bacteriana, care a fost mobilizata prin procesul de transductie fagica.
Sinteza toxinei eritrogene streptococice (scarlatinoasa), precum si a toxinelor botulinice C si D este codificata de fagi temperati. La Cl. perfringens, toxigeneza este legata temporal de procesul sporularii. Sporularea este o conditie obligatorie, dar insuficienta pentru producerea toxinei. Toxina pare a fi un produs al unei gene de sporulare si este o proteina de structurata a tunicii sporale.
Endotoxinele
Termenul de "endotoxina" (introdus de Pffeifer, l892) este impropriu, deoarece lipopolizaharidele (LPS) sunt componente ale suprafetei bacteriilor Gram negative, dar este totusi folosit pentru ca, spre deosebire de exotoxine semnifica asocierea LPS cu celula.
Factorul toxic al bacteriilor Gram negative a fost extras de Boivin si Mesrobeanu (l933). Ei au identificat complexul glico-lipidic, cu antigenul somatic (O) al bacteriilor ce formeaza colonii netede (S-Smooth).
Endotoxinele LPS sunt componente structurale ale peretelui celular, la toate bacteriile Gram negative patogene si se elibereaza dupa dezintegrarea celulei. Sunt relativ termo-stabile si mai putin toxice decat exotoxinele, iar efectele lor sunt lipsite de specificitate. Din punct de vedere chimic, endotoxinele LPS sunt macromolecule complexe ce contin fosfolipide si polizaharide. Toxicitatea lor rezida in fractia fosfolipidica. Nu se denatureaza si nu rezulta anatoxine.
Endotoxinele LPS reprezinta 25% din moleculele de suprafata ale celulei si sunt esentiale pentru integritatea membranei externe.
Efectele endotoxinelor LPS
Termenii de "LPS" si "endotoxina" sunt folositi cu sensuri echivalente, dar LPS semnifica moleculele purificate, iar denumirea de "endotoxina" desemneaza complexul format din LPS si proteinele asociate din membrana externa.
LPS sunt molecule amfifile, cu o parte hidrofoba, capabila sa se dizolve in lipidele membranare si o regiune hidrofila, care poate ramane in faza apoasa. O prima treapta a actiunii LPS o constituie interactiunea dintre molecula amfifila si suprafata celulei sensibile. Molecula LPS poate fi inserata in membrana celulei prin jumatatea hidrofoba sau se ataseaza de receptorii membranari prin jumatatea hidrofila. O modalitate distincta a interactiunii LPS cu macrofagele, este aceea mediata de o proteina plasmatica de faza acuta denumita LBP (LPS binding protein).
Sursele majore de endotoxine sunt urmatoarele: 1)septicemia cu bacterii Gram negative; 2)bacterii originare in microbiota intestinala, datorita leziunilor mucoasei.
O particularitate a LPS, in contrast cu actiunea exotoxinelor, consta in aceea ca simptomele toxice nu se datoreaza actiunii lor directe asupra celulelor sensibile, ci sunt mediate, in mare parte, de citochinele sintetizate de novo de macrofage, ca raspuns la LPS.
LPS sunt molecule foarte imunogene prin stimularea nespecifica, policlonala a limfocitelor B. Anticorpii au specificitate fata de polizaharidul extern si precipita LPS, dar nu neutralizeaza efectele lor toxice. Raspunsul imun este de tip primar, deoarece molecula de LPS este un antigen timo-independent, cu grupari antigenice repetitive.
Lipidul A are proprietati imunogene, iar anticorpii specifici reactioneaza incrucisat cu lipidul A al altor endotoxine, datorita uniformitatii structurii sale.
Antigenul polizaharidic, obtinut prin hidroliza acida isi pastreaza proprietatea de specificitate, dar nu este imunogen, avand proprietatea de haptena.
Efectele endotoxinei se manifesta atat in stare fizica legata de celula, cat si dupa ce a fost eliberata prin moartea si liza celulei sau printr-un proces de "inmugurire", care nu afecteaza viabilitatea acesteia. Din aceasta cauza, cele doua stari, endotoxemia si bacteriemia pot sa coexiste sau sa se manifeste in etape distincte ale procesului infectios. Proprietatile endotoxice ale celulelor bacteriene, vii sau omorate sunt aceleasi cu ale preparatului de endotoxina. In organismul infectat nu este o corelatie lineara intre nivelul bacteriemiei si endotoxemiei si respectiv, intensitatea efectelor endotoxice. Capacitatea organismului de a detoxifica este un factor modulator esential al manifestarilor endotoxice. Glucocorticoizii au efect protector anti-endotoxic, probabil prin modificarea permeabilitatii capilare.
Endotoxinele nu au rol in colonizarea si penetrarea suprafetei mucoase, deoarece lipidul A are o localizare profunda in membrana externa. Tulpinile invazive si neinvazive de Salmonella produc cantitati similare de endotoxina. Endotoxina are rol in initierea infectiei, probabil prin inducerea unei scaderi tranzitorii a capacitatii de aparare a gazdei, prin intarzierea declansarii unui raspuns inflamator.
Dozele mici de toxina maresc rezistenta organismului la infectiile bacteriene si virale prin stimularea activitatii fagocitare si respectiv, a producerii de interferon.
Endotoxinele initiaza calea alterna a activarii complementului, dar efectul nu este bacteriocitoliza, deoarece reactia se produce la distanta de membrana externa. In vitro au efect mitogenic asupra limfocitelor B. LPS au proprietati adjuvante, deoarece stimuleaza raspunsul imun specific fata de un antigen administrat simultan.
Dozele mari de endotoxine determina o serie de manifestari patologice sistemice, nespecifice:
hipertermie (febra), prin actiunea lor asupra centrilor termoreglarii, chiar dupa administrarea unor doze minimale. Inducerea febrei este semnul marcant al toxicitatii LPS. Febra este efectul indirect al actiunii LPS asupra macrofagului, care secreta cateva citochine: IL-1, TNF β, IFN , cu actiune directa asupra hipotalamusului. Starea febrila se instaleaza brusc, este monofazica, mediata de prostaglandine;
leucopenie urmata de leucocitoza. Efectul se exercita direct si rapid asupra leucocitelor. Ele parasesc patul vascular si se retrag aproape instantaneu in plaman si in alte tesuturi, in proportie de circa 60%. Dupa circa 4 ore, numarul leucocitelor circulante creste peste limitele normale. Creste numarul hematiilor circulante, datorita eliberarii in circulatie a rezervelor celulare din centrele de formare si depozitare a eritrocitelor;
modificari cardiovasculare: legarea endotoxinei de celulele hepatice sau de alte celule induce eliberarea rapida a aminelor biogene (histamina, serotonina) si a peptidelor (bradikinina) din depozitele celulare. Aminele produc o hipertensiune tranzitorie, urmata de hipotensiune severa, hipovolemie si formarea cheagurilor vasculare de fibrina. Endotoxinele activeaza sistemul de coagulare sanguina;
efecte metabolice: inhiba sistemele enzimatice ale gluconeogenezei si ale sintezei glicogenului.
Din punct de vedere clinic, dozele mari de endotoxina produc urmatoarea secventa de modificari: frison, febra, somnolenta, dispnee, modificari ale dinamicii tranzitului intestinal, diaree sanguinolenta, hiperglicemie incipienta urmata de hipoglicemie, paralizie, coma, moarte. Acest tablou de modificari se succede in circa 24 de ore dupa injectarea preparatului la animalele sanatoase si corespund starii de soc endotoxic, consecinta a endotoxemiei. Endotoxemia este rezultatul supraincarcarii celulelor cu rol de aparare, ca urmare a revarsarii endotoxinelor in circulatie. Starea de endotoxemie nu este totdeauna urmata de soc si moarte. Nu se cunosc cauzele marilor diferente de raspuns si reactivitate, la starea de endotoxemie. Socul fatal este consecinta interactiunii endotoxinei cu sistemul de coagulare a sangelui, pe care il activeaza si il amplifica. S-a emis ipoteza ca instalarea socului ireversibil este favorizata de absorbtia in sange a endotoxinelor produse de microbiota intestinala. In mod normal, mecanismele de detoxifiere a organismului sunt eficiente, dar starea de soc se instaleaza consecutiv diverselor tulburari functionale, care impiedica o reactivitate optima fata de infectia cu bacterii Gram negative.
Conditiile de aparitie a procesului infectios
Simpla prezenta a unui microorganism patogen in mediul inconjurator nu este suficienta pentru a produce o infectie la o gazda sensibila. Pentru initierea procesului infectios sunt necesare urmatoarele conditii obligatorii: existenta unui izvor de infectie, a unei cai de eliminare a agentului infectios, a unei cai de transmitere si a unei porti adecvate de intrare in organismul sensibil. Domeniul specializat al patologiei infectioase care studiaza caile de eliminare, transmitere si patrundere a agentilor patogeni in organism se numeste epidemiologie.
a) Izvorul de infectie
Necesitatea existentei izvorului de infectie deriva din faptul ca mediul extern (sol, apa, aer) este nefavorabil cresterii si multiplicarii majoritatii agentilor patogeni. Mediul extern asigura, cel mult, conditii de supravietuire temporara a microorganismelor si de pastrare a infectiozitatii virusurilor. Perpetuarea microorganismelor patogene in natura este conditionata de existenta unui izvor sau rezervor de infectie, in care agentul patogen se multiplica. Aici se realizeaza procesul de acumulare naturala, de unde agentul patogen se disemineaza si contamineaza alte organisme.
Izvorul de infectie este diferit, in functie de agentul patogen si de spectrul sau de gazda. Unele microorganisme si virusuri infecteaza numai gazdele animale si produc zoonoze, omul fiind infectat in situatii accidentale. Alteori, agentii patogeni produc boli infectioase (transmisibile) strict caracteristice omului (denumite antroponoze) fara sa infecteze organisme animale. In acest caz, omul reprezinta unicul izvor de infectie in natura, pentru viroze (variola, varicela, oreionul, poliomielita, guturaiul, gripa, hepatite) sau pentru bacterioze (dizenteria bacteriana, febra tifoida, febrele paratifoide, tusea convulsiva, sifilisul, gonoreea, lepra). Pentru unele maladii infectioase (lepra, sifilisul, variola, rujeola), omul bolnav este unicul rezervor natural al infectiei. In alte cazuri (de exemplu, febra tifoida, scarlatina, difteria, poliomielita, holera), agentul infectios este transmis atat de omul bolnav, cat si de cel sanatos, purtator de agenti patogeni. In unele cazuri, purtatorii sunt fostii bolnavi care s-au imunizat, nu mai prezinta nici un simptom clinic, dar pastreaza in organism agentul patogen, temporar sau pentru totdeauna, pe care il elimina in mediu. Purtatorii sunt foarte importanti pentru diseminarea agentilor patogeni in mediul extern, deoarece fiind aparent sanatosi sunt greu de depistat si se deplaseaza liber in colectivitati, unde pot transmite infectia la organismele receptive.
O alta categorie de boli infectioase sunt comune omului si animalelor si sunt cunoscute sub denumirea generica de antropozoonoze. Izvorul de infectie este reprezentat de animale domestice sau salbatice: cainele pentru rabie, febra butonoasa, leptospiroze; bovinele pentru salmoneloze, vaccina, bruceloza, tuberculoza, febra Q; sobolanul pentru salmoneloze, leptospiroze, febra muscaturii de sobolan (febra sodoku), turbare, pesta (ciuma), tifos murin; pasarile pentru encefalite virale, gripa, ornitoze, salmoneloze etc.
Controlul antropozoonozelor este foarte greu pentru ca in circuitul agentului patogen intra animalele salbatice. De exemplu, turbarea este raspandita la vulpe, veverita, liliac, de unde este transmisa la caine, iar de aici la om. Chiar daca incidenta bolii scade dupa imunizarea cainilor prin vaccinare, virusul nu se poate elimina din populatiile de animale salbatice.
Unele microorganisme patogene se dezvolta, in primul rand in mediile naturale (apa, sol) si numai accidental infecteaza gazdele: de exemplu, agentii tetanosului si ai gangrenei gazoase. Agentii se multiplica in sol si pentru a se mentine nu este necesara transmiterea la o gazda. Clostridium botulinum, agentul botulismului, se dezvolta numai in sol, de unde ajunge in conservele alimentare, se multiplica si produce toxina, care dupa ingestie produce manifestari patologice.
b) Calea de eliminare a agentilor patogeni
Agentii patogeni care se multiplica in izvorul natural de infectie sunt eliminati in mediu si pot sa infecteze gazde noi. Calea de eliminare este conditionata de localizarea lor specifica in organism. Principalele cai de eliminare a agentilor patogeni sunt cea intestinala si cea respiratorie.
Calea intestinala este comuna bacteriilor enterotrope, prin intermediul materiilor fecale, care pot contine intermitent sau continuu, cantitati mari de agenti patogeni ai febrelor tifoide si paratifoide, dizenteriei, holerei, ai toxiinfectiilor alimentare, precum si virusuri (virusul hepatitei A, virusul polio).
Calea respiratorie asigura raspandirea agentilor patogeni ai infectiilor respiratorii (difteria, tusea convulsiva, rujeola, variola, varicela, oreionul, gripa, guturaiul), prin intermediul secretiilor nazofaringiene si bucale, proiectate in timpul tusei, stranutului, vorbirii. Pe aceasta cale, omul bolnav sau infectat raspandeste in jur o pulbere fina de aerosoli constituiti din picaturi microscopice de secretii incarcate cu agenti patogeni. Intr-un stranut se pot elimina intre l0 000 - l00 000 celule bacteriene. Cele rezistente la uscaciune isi pastreaza viabilitatea pentru perioade lungi de timp, atasate de particulele inerte.
Alte cai de eliminare: rani si supuratii (in cazul infectiilor produse de stafilococi, streptococi, bacilul piocianic, gangrena gazoasa); calea urinara (pentru agentii febrei tifoide, leptospirozei, febrei Q, tuberculozei renale); secretia lactata (agentul brucelozei, tuberculozei, febrei Q).
Pentru agentii patogeni care produc infectii generalizate si care nu au o cale naturala de eliminare din organism, iesirea se realizeaza prin intermediul unui artropod hematofag care se hraneste cu sangele infectat (de exemplu, agentii tifosului exantematic, ai pestei, ai encefalitelor virale, ai paludismului) sau iesirea se face pe cale artificiala, prin seringa sau instrumente chirurgicale infectate (agentii hepatitelor virale B, C, D, ai sifilisului).
c) Calea de transmitere a agentilor infectiosi
Dupa ce au fost eliminate din organismul bolnav sau purtator, agentii patogeni trebuie sa fie transmisi la o noua gazda receptiva. Transmiterea se realizeaza in mai multe modalitati.
Transmiterea prin contact direct are loc in cazul in care, intre organismul infectat si cel receptor exista o legatura directa si se face prin muscatura (agentii turbarii, ai febrei muscaturii de sobolan), prin supt (la animale, agentii febrei Q, tuberculozei, iar la om, M. tuberculosis, agentii infectiilor piogene), prin sarut(agentii tuberculozei pulmonare, sifilisului, mononucleozei infectioase), prin contact cutanat direct (agentii dermatomicozelor, furunculozei), prin contact sexual (agentii sifilisului, gonoreii). Cand agentul cauzal este transmis prin caile genitale, infectia se numeste venerica.
De cele mai multe ori, transmiterea infectiei este indirecta, prin interpunerea mai mult sau mai putin evidenta, a unui factor de mediu neanimat denumit vehicul, sau a unui organism denumit vector, care se interpune intre izvorul de infectie si organismul receptor:
transmiterea prin intermediul produselor alimentare - carne, lapte, oua, in cazul in care provin de la animale infectate sau sunt contaminate prin manipulare de catre o persoana (bolnava sau purtatoare) care elimina agenti patogeni. Bacteriile patogene, de regula, se multiplica in produsele alimentare. Astfel se transmit agentii febrei tifoide, ai toxiinfectiilor alimentare (cu Staphylococcus si Salmonella), ai febrei Q, ai infectiei carbunoase, ai botulismului, ai tuberculozei;
transmiterea prin intermediul obiectelor folosite de un bolnav (vesela, rufe, imbracaminte, carti, jucarii etc) este posibila un timp limitat dupa contaminarea acestora, proportional cu rezistenta agentilor patogeni la conditiile de mediu (agentii tuberculozei, difteriei, scarlatinei, variolei);
transmiterea prin vectori se realizeaza prin intermediul artropodelor, in special hematofage (insecte, capuse), care preiau agentii patogeni de la gazda infectata si ii transmit la o noua gazda, fie prin intepatura, fie depunandu-i pe tegumentul intact sau lezat. Uneori, vectorul are numai rol mecanic pentru agentul patogen, pe care il transporta in tubul digestiv sau pe suprafata corpului. Musca este vector mecanic pentru agentul patogen al febrei tifoide, al dizenteriei. Alteori, vectorul este el insusi infectat si realizeaza o transmitere biologica. In acest caz, agentul patogen se multiplica masiv in corpul vectorului (de exemplu, Ricketsia prowazeki, agentul tifosului exantematic) sau strabate o faza a ciclului sau vital (de exemplu, sporozoitul Plasmodium, in corpul tantarului. Flavivirusurile (agentii encefalitelor) se multiplica in organismul artropodelor vectoare, fara sa produca leziuni tisulare;
transmiterea prin intermediul aerului se face prin inhalarea picaturilor septice, raspandite de un bolnav sau de un purtator de agenti patogeni. Cele mai eficiente sunt picaturile septice foarte mici (nuclei), care pot ramane in suspensie in aer (ca aerosoli), mult timp dupa eliminarea lor din organism. Astfel se transmit agentii gripei, guturaiului, rujeolei, rubeolei, varicelei, scarlatinei, tusei convulsive;
transmiterea hidrica este proprie infectiilor intestinale (agentii febrelor tifoida si paratifoida, holerei, hepatitei A, enterovirozelor) si leptospirozelor si se produc prin contaminarea accidentala a unei surse de aprovizionare cu apa, cu agentii patogeni proveniti din dejectii umane sau animale;
transmiterea prin intermediul solului are loc in cazul unor plagi, de regula adanci, in care au fost antrenate granule de sol. Astfel se transmit infectiile cu B. anthracis, cu Cl. tetani, agentii gangrenei gazoase (Welchia perfringens, Cl. oedematiens, Cl. histolyticum).
d) Poarta de intrare in organism
Pentru ca agentii patogeni sa determine o boala infectioasa este necesar ca ei sa invinga barierele naturale protectoare ale gazdei. Poarta de intrare in organism corespunde, in general, caii prin care s-a facut eliminarea agentului patogen din organismul infectat.
De cele mai multe ori, poarta de intrare in organism este reprezentata de calea respiratorie (in infectii ca oreionul, guturaiul, gripa, rujeola, variola, varicela, difteria, tusea convulsiva, scarlatina), calea digestiva(agentii febrei Q, hepatitei virale A, poliomielitei), calea cutanata(pentru agentii infectiei carbunoase, tularemiei, febrei Q, pentru agentii transmisi prin muscatura, prin intepatura vectorilor, prin ranire).
Unii agenti patogeni patrund in organismul receptiv pe mai multe cai, din care una este principala, dar determina aceleasi manifestari indiferent de poarta de intrare. Alti agenti produc maladii cu caractere clinice diferite, in functie de poarta de intrare, ca unic factor determinant si fara relatie cu vreun caracter biologic particular al agentului patogen. De exemplu, B. anthracis, dupa patrunderea pe cale tegumentara produce carbunele cutanat, iar dupa ingestia alimentelor contaminate produce un proces patologic intestinal. Daca infectia se face pe cale respiratorie, rezultatul este infectia pulmonara.
Tipuri de infectii
Infectia reprezinta totalitatea proceselor biologice care se desfasoara in organismul uman sau animal, ca urmare a patrunderii si multiplicarii agentilor patogeni.
Pentru initierea procesului infectios, agentul patogen trebuie sa patrunda in organism pe o cale adecvata, iar organismul sa fie receptiv (sa permita multiplicarea agentului infectios). In conceptia moderna, procesul infectios trebuie interpretat ca rezultat al interactiunii dinamice dintre micro- si macroorganism. In conceptia ecologica, agentul infectios este forta motrice a procesului infectios, iar reactivitatea imunitara a organismului conditioneaza intensitatea, extinderea, gravitatea si insasi posibilitatea aparitiei procesului infectios.
In raport cu momentul in care se face infectia organismului uman si animal se disting patru tipuri de infectii.
a) Infectia germinala se produce in cazurile in care ovulul sau spermatozoidul sunt purtatorii agentului infectios, pe care-l transmit celulei-ou (pe verticala), odata cu caracterele ereditare. Acest mecanism de infectie nu s-a demonstrat la om si nici la animalele superioare. A fost descris la artropode (capuse din g. Ornithodorus, Dermacentor, Rhipicephalus), infectate astfel cu bacteria spiralata Borrelia recurentis, agentul patogen al febrei recurente de capuse. Ele devin astfel, rezervoare naturale de infectie.
b) Infectia transplacentara (intrauterina) este demonstrata si la om. Uneori este conditionata de producerea prealabila a unor leziuni placentare sub actiunea agentului patogen, ca de exemplu, T. pallidum, Brucella, Coxiella burnetii (agentul febrei Q). Alteori, infectia are loc fara modificari placentare (infectia variolica, rujeolica, hepatitica etc.).
c) Infectia intrapartum se produce in timp ce fatul traverseaza caile genitale (la expulzie), cu microorganisme patogene ale tractului genital matern (oftalmia si conjunctivita gonococica a nou-nascutilor).
d) Infectia postpartum se produce dupa nastere. In aceasta categorie sunt cuprinse majoritatea infectiilor omului si animalelor. Ele se manifesta in special dupa epuizarea imunitatii pasive, conferita de transferul anticorpilor materni prin placenta si colostru.
In raport cu originea microorganismelor care le produc, infectiile sunt de doua categorii:
exogene, provocate de microorganisme care patrund pe o cale adecvata;
endogene, provcate de microorganisme autohtone (indigene), potential patogene ale microbiotei normale, datorita scaderii rezistentei locale sau generale a gazdei, dupa iradiere, dupa tratamentul cu medicamente citotoxice, hormoni corticosteroizi, malnutritie, obstructia cailor de excretie. Aceste infectii nu induc un raspuns imun detectabil si au tendinta sa revina ciclic sau sa evolueze lent.
Din punct de vedere microbiologic, starea caracteristica a organismului infectat cu un agent patogen, inainte de declansarea simptomelor este aceea de contaminare.
Colonizarea sau multiplicarea are loc dupa patrunderea agentului infectios pe o cale adecvata. Multiplicarea realizeaza conditia cantitativa obligatorie a procesului infectios, deoarece, in mod obisnuit, doza contaminanta a agentului patogen este prea mica pentru a initia imediat procesul infectios. Cu cat tulpina este mai virulenta, cu atat doza contaminanta necesara initierii infectiei este mai mica.
Dupa contaminare, agentul patogen se multiplica la poarta de intrare sau este transporat prin sange si limfa, la nivelul unor situsuri specifice localizate la distanta.
Consecinta directa a colonizarii (multiplicarii) agentului patogen la poarta de intrare, local se constituie un focar de inflamatie, adica o reactie de aparare locala a organismului, cu mobilizarea leucocitelor. Rezultatul inflamatiei este supuratia (formarea puroiului), la nivel cutanat si la nivelul mucoaselor respiratorie, digestiva, urinara, genitala.
Modalitatile de manifestare a patologiei infectioase
In functie de natura agentului patogen, de preponderenta unuia sau altuia dintre factorii de virulenta, patologia infectioasa imbraca urmatoarele aspecte:
a) ntoxicatia de natura bacteriana fara infectie, este o situatie de exceptie, in cazul botulismului. Cl. botulinum se multiplica in alimentele conservate si produce toxina. Consumul unor astfel de alimente determina intoxicatia.
b) oxiinfectiile sunt procese alterative produse de bacteriile care se multiplica la poarta de intrare si elibereaza toxine difuzibile in organism. De exemplu, Cl. tetani se multiplica in plaga anaeroba provocata de ranire, iar toxina ajunge prin difuzie axonala retrograda, la motoneuronii din coloanele anterioare ale maduvei, unde isi exercita efectul. Asemanator, C. diphteriae se multiplica la nivelul mucoasei faringiene, iar toxina determina efecte patologice la distanta, asupra fibrei musculare cardiace, celulei nervoase sau glandelor suprarenale;
c) oxiinfectiile alimentare sunt rezultatul consumului alimentelor infectate (carne, lapte, oua) provenite de la animalele bolnave sau contaminate in timpul manipularii. De exemplu, Salmonella sp. si Str. aureus se multiplica excesiv in alimente (carne) si concomitent se acumuleaza endotoxine si respectiv enterotoxine. Consumul alimentelor echivaleaza cu ingestia culturii bacteriene. Concomitent cu efectele toxinelor se manifesta si procesul infectios initiat de celulele vii ingerate;
d) nfectia localizata (neinvaziva) este produsa de agentii patogeni care de obicei raman localizati la poarta de intrare in cursul intregului proces infectios. Toxinele eventual sintetizate, se raspandesc sistemic, pe cale sanguina (de exemplu, C. diphteriae, Cl. tetani). In aceste cazuri, infectia locala are caracterul unei toxiinfectii. O forma particulara a infectiei locale este infectia de focar. Aceasta este insotita de o reactie inflamatorie minima la nivelul focarului initial, dar se exprima prin fenomene morbide la distanta. Maladia reumatismala este considerata ca o infectie tipica "de focar", in timpul careia streptococul localizat la poarta de intrare (amigdale) elimina substante cu efect alterativ si alergizant. Uneori, infectia localizata este o caracteristica a microorganismelor lipsite de invazivitate
e) Infectiile invazive sunt produse de agentii infectiosi care, dupa o perioada initiala de multiplicare la poarta de intrare, se raspandesc la distanta pe diferite cai:
prin contiguitate, tipica agentilor infectiosi cu multiplicare intracelulara (virusuri, rickettsii). Dupa eliberare prin necroza celulei, prin inmugurire, prin permeabilizarea sau ruperea membranei, agentii infectiosi infecteaza celulele sensibile invecinate;
prin continuitate, in cazul agentilor patogeni care se multiplica intr-un tesut prevazut cu un sistem canalicular. Diseminarea se face din aproape in aproape. Astfel, infectiile respiratorii progreseaza spre bronhii (bronsite), tesutul pulmonar (pnreumonii), pleure (pleurite). De la nivelul apendicelui, agentul diseminat produce peritonita. La nivelul cailor urogenitale si biliare, agentul infectios intalneste o rezistenta minima conferita de scurgerea secretiilor si de unele manifestari de mobilitate locala, orientate in sens opus progresiei infectiei;
pe cale limfatica. De la nivelul focarului infectios primar, agentii infectiosi sunt preluati de curentul limfatic. Consecinta imediata este inflamarea vaselor limfatice si a ganglionilor regionali (limfadenita). Infectia poate sa progreseze pana la varsarea limfei in sange si rezultatul este bacteriemia sau viremia. Agentii infectiosi pot trece direct in sange, la nivelul leziunilor peretilor vasculari.
Bacteriemia semnifica prezenta unui numar mic de bacterii in torentul circulator, pentru o perioada strict limitata de timp. Sistemele de aparare, in special sistemul fagocitar, elimina bacteriile circulante. Bacteriemia este consecutiva proceselor infectioase acute pulmonare, interventiilor chirurgicale, extractiei dentare, absorbtiei intestinale si chiar periajului dentar. In sange, cu rare exceptii (bacilul carbunos, al pestei) bacteriile nu se multiplica. De aceea, prezenta lor in sange semnifica o eliberare dintr-un focar de infectie, atat de abundenta, incat, temporar, depaseste capacitatea protectoare a mecanismelor de aparare a organismului.
Extinderea infectiei pe una dintre caile mentionate poate determina fie o infectie regionala, care corespunde extinderii infectiei (diseminarii) pe teritorii intinse (de exemplu, sistemul limfatic regional si eventual vasele sanguine tributare zonei), fie o infectie generalizata.
Dintr-o infectie regionala, agentii patogeni diseminati pe o cale adecvata pot dobandi localizari secundare, in orice tesut sau organ, unde initiaza noi focare infectioase. Aceste localizari pot fi unice (de exemplu, pe valvulele cardiace, in plamani, meninge, ficat, vezica biliara, creier, rinichi etc.) sau multiple (miliare) denumite septicopioemii, care evolueaza spre infectii generalizate.
Diseminarea sanguina determina infectii generalizate. Ele sunt precedate de trecerea masiva a bacteriilor dintr-un focar infectios, in sange si semnifica incapacitatea mecanismelor de aparare a gazdei de a reactiona eficient fata de agentul infectios. Existenta stabila a unui numar mare de bacterii in sange caracterizeaza infectia septicemica. Septicemia este consecinta descarcarii masive de bacterii dintr-un focar de infectie, vascularizat. Starea septicemica poate fi insotita de toxemie, ca urmare a difuziei toxinelor din focar (abces, meningita).
Uneori, faza septicemica nu este precedata de multiplicarea agentului patogen la poarta de intrare. Agentii patogeni ai infectiilor transmise de insectele hematofage (tifos exantematic, febra recurenta) sau prin muscatura (leptospiroze) strabat mucoasele si trec direct in sange.
Infectia bacteriana produce modificari de tip alterativ ale organismului gazda. Ele sunt primare (directe), datorate factorilor de patogenitate bacteriana (intoxicatii, toxiinfectii, bacteriemii) si secundare, mediate de factorii umorali si celulari ai sistemului imunitar (starile de hipersensibilitate).
Posibilitati de evolutie a procesului infectios
Infectia poate evolua in doua modalitati: inaparent sau aparent, ca boala infectioasa.
Infectia inaparenta caracterizeaza acele situatii in care, procesul infectios nu se exteriorizeaza prin simptomatologie clinica. De exemplu, infectia tuberculoasa inaparenta se detecteaza prin reactia de hipersensibilitate intarziata la tuberculina. Reactia este pozitiva la un procent mare de indivizi, dar maladia tuberculoasa clinica se manifesta la un procent mic din totalul persoanelor pozitive pentru testul tuberculinei. De cele mai multe ori, infectiile inaparente se detecteaza prin reactii serologice in vitro sau prin reactii de hipersensibilitate in vivo.
Uneori, infectia asimptomatica poate fi mortala. Explicatia consta in paralizia mecanismelor de aparare a gazdei.
Infectia aparenta este cea care se manifesta printr-un ansamblu de semne clinice obiective si subiective, specifice si nespecifice, consecinte ale alterarilor produse de agentul infectios si de produsele activitatii sale.
Maladia infectioasa prezinta urmatoarele caracteristici; a)este produsa de microorganisme vii sau de toxine ale microorganismelor; b)poate realiza o imunitate specifica de durata variabila; c)organismul bolnav poate deveni sursa de imbolnavire a indivizilor sanatosi; d)este specifica, in sensul ca aceiasi maladie este produsa totdeauna de acelasi agent cauzal, desi forma clinica se poate prezenta sub aspecte variate.
Etapele maladiei infectioase
In evolutia sa, procesul infectios aparent parcurge mai multe etape distincte, separate in timp.
Perioada de incubatie sau perioada initiala este intervalul de timp scurs intre momentul patrunderii agentului patogen in organism si acela al debutului maladiei. Perioada este lipsita de simptome clinice evidente. Durata ei este variabila, in functie de natura agentului patogen, iar pentru acelasi agent, in raport cu doza infectanta, cu virulenta agentului patogen si cu reactivitatea imunitara a gazdei si se masoara in zile, saptamani, luni sau este foarte scurta. Pentru infectiile exogene, durata incubatiei se poate determina relativ precis, prin infectia experimentala a animalelor de laborator sau a voluntarilor. Pentru infectiile endogene, perioada de incubatie nu se poate determina. Din momentul contaminarii este posibila scurgerea unei perioade foarte lungi de timp pana cand agentul poate sa initieze un proces infectios. In aceasta perioada, agentul patogen se multiplica, se localizeaza la nivelul situsului receptiv si eventual elaboreaza substante toxice.
Cel mai adesea, agentii infectiosi manifesta un organotropism evident, deoarece, in general, isi gasesc conditiile optime de dezvoltare intr-un anumit tesut. De exemplu, indiferent de calea de patrundere, vibrionul holeric se localizeaza in intestinul subtire, bacilul dizenteric - in mucoasa intestinului gros, Brucella - in placenta bovinelor si ovinelor, datorita concentratiei mari de eritrol (dar nu in placenta umana), Rickettsia prowazeki - in celulele epiteliale ale capilarelor SNC, B. pertusis - exclusiv in mucoasa bronsica.
B. anthracis se multiplica in orice tesut, chiar si in sange.
Debutul bolii este marcat de momentul in care numarul de agenti infectiosi si cantitatea de toxine acumulate au atins un nivel critic. Debutul poate fi brusc sau lent si este caracterizat din punct de vedere clinic, prin instalarea (brusca sau gradata) a semnelor bolii: febra, cefalee, frisoane, algii musculo-articulare. Debutul marcheaza inceputul perioadei de invazie a agentului infectios de la locul multiplicarii primare sau a eliberarii unei cantitati prag de toxina, spre noi zone sensibile, iar din punct de vedere cronologic si clinic semnifica inceputul perioadei de stare.
Perioada de stare este intervalul de timp, in care maladiile infecto-contagioase isi desfasoara un tablou clinic cu simptome caracteristice, de amplitudine maxima, decisiva pentru evolutia ulterioara. In aceasta perioada poate sa survina decesul.
La sfarsitul perioadei de stare, simptomele dispar brusc - in crisis (de exemplu, in pneumonia bacteriana), insotita de sterilizarea bacteriologica sau lent - in lysis (in cazul febrei tifoide). Organismul se poate steriliza sau poate sa ramana infectat pentru o perioada nedefinita de timp.
In perioada de stare, organismul se imunizeaza abundent cu antigene ale agentului patogen si se sintetizeaza anticorpi. Imunizarea are ca rezultat, de regula, sterilizarea bacteriologica sau virologica a organismului. Uneori, organismul nu se sterilizeaza si in focarele de infectie greu accesibile efectorilor raspunsului imun, agentul persista, consecinta fiind o infectie cronica.
Convalescenta este perioada de timp in care organismul isi reface potentialul de activitate, anterior imbolnavirii.
Infectia cronica corespunde unei stari de echilibru intre agentul infectios si organismul gazda, caracterizata prin faptul ca vindecarea clinica (disparitia simptomelor) nu este insotita de sterilizarea organismului. Focarele de infectie cronica au localizari greu accesibile factorilor de aparare a organismului sau medicatiei: in SNC, in viscerele parenchimatoase (ficat, rinichi, splina), in sinusurile osoase, in glandele bine incapsulate (prostata). Infectia cronica poate sa persiste fara simptome clinice (sifilisul latent) sau poate evolua cu reacutizari intermitente la diferite perioade (bruceloza, tuberculoza, sifilisul tetiar).
Starea de purtator este, fie consecinta unei infectii cronice, fie a persistentei pentru o perioada de timp, a unui focar de infectie, in organismele clinic sanatoase. Uneori, fostii bolnavi tolereaza la nivelul mucoaselor, agentul patogen pe care il elimina timp de cateva saptamani (Str. pyogenes, C. diphteriae, V. cholerae). Alteori, bacteriile si virusurile sunt eliminate pentru perioade indelungate (de ordinul anilor): S. typhimurium ramane localizata in vezica biliara, de unde se elimina intermitent, iar bolnavii de difterie pot ramane purtatori ai bacilului C. diphteriae. Purtatorii creeaza probleme epidemiologice dificile, deoarece reprezinta izvoare de infectie greu de depistat. Purtatorii sunt foarte periculosi pentru sanatatea publica, mai ales daca vin in contact cu alimentele, ca manipulatori sau preparatori ai acestora.
Tipuri de evolutie epidemiologica a infectiei
Din punct de vedere epidemiologic, maladia infectioasa poate evolua pe mai multe nivele de extindere spatio-temporala si incidentala:
evolutia sporadica semnifica aparitia cazurilor izolate de imbolnavire, atat in timp cat si in spatiu;
evolutia endemica este definita prin aparitia cazurilor relativ rare de infectie, care se mentin numeric constante intr-o colectivitate si apar cu relativa regularitate, de obicei sezonier, la intervale variabile de timp. Majoritatea indivizilor populatiei sunt imunizati si deci indemni fata de agentul infectios;
evolutia epidemica, caracterizeaza evolutia rapida a unei infectii intr-un interval scurt de timp, cu un numar mare de cazuri clinice intr-o colectivitate (camin, cazarma, sat, regiune, tara). Se cunosc epidemiile de ciuma, gripa, rujeola, scarlatina;
evolutia pandemica este aceea in care maladia infectioasa se extinde foarte rapid pe un teritoriu foarte larg (tari, continente), cu un numar foarte mare de cazuri clinice. Este caracteristica infectiei gripale si este usurata de mijloacele de deplasare rapida, la distante foarte mari.
Fata de un agent patogen, absent in mod obisnuit dintr-un areal, populatia poate fi foarte susceptibila. La primul contact cu populatia receptiva, agentul patogen declanseaza o epidemie exploziva, dar odata cu instalarea starii de imunitate specifica, incidenta maladiei scade.
Daca agentul patogen isi modifica specificitatea antigenica, epidemia evolueaza si intr-o populatie imunizata, deoarece, fata de noile variante antigenice, populatia nu poseda anticorpi specifici.
Epidemiile sunt conditionate de standardul de viata al comunitatii. Maladia tuberculoasa este o reflectare a acestei conditii, in timp ce bolile venerice sunt favorizate de promiscuitatea sexuala.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3357
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved