| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
In prezent, plasmidele sunt definite ca fiind repliconi neesentiali, capabili sa se replice fizic independent de cromozom si transmisi in forma extracromosomala, pe verticala (de la o generatie de indivizi la alta) sau pe orizontala (de la un individ la altul in cadrul aceleiasi generatii). In general, plasmidele reprezinta un fenomen al lumii procariote. Cu toate acestea, au fost descrise structuri genetice plasmidiale si la unele grupuri de eucariote inferioare (drojdii, fungi, mucegaiuri) si chiar si la eucariote superioare (plasmidele din mitocondrii si cloroplaste la unele celule vegetale).
Exista cercetatori care incadreaza plasmidele in categoria organismelor acelulare. In contrast cu aceasta teorie, alti cercetatori considera plasmidele ca nefiind organisme, ci alaturi de virusuri, transposoni si secvente de insertie, ar apartine unor specii moleculare speciale, de elemente genetice mobile (ADN accesoriu) (P. Sykora 1992).
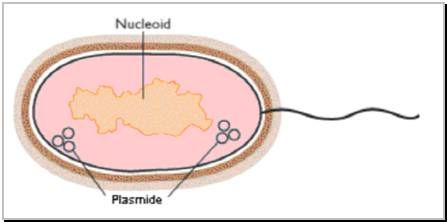
Figure 3.3. Reprezentare schematizata a nucleoidului si a plasmidelor bacteriene.
1 Principalele categorii de gene codificate de plasmide
Fiind repliconi fizic independenti, toate plasmidele poarta gene implicate in propria replicare, stabilitate si partitie. In afara de acestea, majoritatea genelor codificate de plasmide confera caractere adaptative, avantajoase gazdelor bacteriene. Cele mai frecvente categorii de gene purtate de plasmide sunt :
- gene de rezistenta la antibiotice
Plasmidele de rezistenta au fost descoperite in Japonia in anii 50 la izolate clinice de Shigella dysenteriae si s-a constatat ca aceste gene nu erau plasate pe cromosomul bacterian. Au fost denumite factori R. Folosirea abuziva si/sau incorecta a antibioticelor au determinat, pe de o parte, raspandirea in populatiile de bacterii patogene a determinantilor de antibiorezistenta, iar pe de alta parte, dezvoltarea unor mecanisme de antibiorezistenta extrem de eficiente.
In ceea ce priveste originea genelor de antibiorezistenta, au fost emise 2 ipoteze majore:
a-genele de antibiorezistenta existente in prezent in foarte multe bacterii patogene ar avea originea in gene similare prezente in bacteriile ce produc antibiotice (si care, de regula, sunt bacterii din sol); in aceasta situatie se presupune ca ar fi avut loc evenimente de transfer de gene pe orizontala intre bacterii din sol si bacterii patogene;
b-genele de antibiorezistenta ar fi evoluat din gene ²house-keeping² (de exemplu: proteinele ce efluxeaza tetraciclina ar putea sa fi evoluat din proteinele de transport al nutrientilor).
- gene de toleranta la metale grele (ioni mercurici, derivati organomercurici, nichel, cobalt, plumb, cadmiu, bismut, antimoniu, zinc, argint)
- gene de rezistenta la agenti intercalanti, radiatii UV, radiatii X, bacteriofagi, bacteriocine
- gene ce codifica factori de patogenitate si de virulenta
- gene ce codifica bacteriocine
- gene ce codifica antibiotice
- gene implicate in metabolismul opinelor (plasmidele Ti de la Agrobacterium)
- gene ce codifica capacitati metabolice deosebite, de exemplu folosirea unor substante xenobiotice ca sursa unica de carbon si energie la specii de Pseudomonas
- gene fixatoare de azot (genele nif de la Rhizobium si Bradirhizobium)
- gene ce codifica enzime proteolitice, amilolitice etc
- gene ce codifica transferul plasmidelor pe orizontala prin proces de conjugare bacteriana (regiunea tra)
Deci, in afara de genele implicate in propria replicare, stabilitate si partitie, plasmidele mai pot purta si gene ce codifica caractere - unele esentiale, altele neesentiale pentru bacteria gazda. Caracterele esentiale reprezinta cai metabolice centrale ale celulei bacteriene, plasmidele purtatoare fiind prezente la marea majoritate a izolatelor naturale ale unor tulpini bacteriene (de exemplu, unele plasmide mari de la Agrobacterium tumefaciens, Rhizobium leguminosarum).
Unele caractere neesentiale confera avantaje selective gazdelor purtatoare, permitand supravietuirea acestora in anumite conditii de mediu. In aceasta categorie intra genele plasmidiale ce codifica rezistenta la antibiotice si toleranta la metale grele (asemenea gene sunt prezente la multe tulpini de bacterii patogene). In absenta antibioticelor sau a metalelor grele ca factori de presiune selectiva, procesele de replicare, mentinere si partitie a plasmidelor reprezinta un efort metabolic suplimentar pentru celulele bacteriene. Ca urmare, in doar cateva pasaje, aceste plasmide sunt eliminate din populatia bacteriana prin subreplicare. Ele se mentin doar intr-o proportie scazuta a populatiei bacteriene, fapt ce ar putea fi considerat o ²economie² metabolica la nivelul intregii populatii bacteriene.
Exista insa si asa-numite plasmide ²criptice², care nu par sa ofere gazdei nici un caracter avantajos. Interesant este faptul ca, desi pot fi incadrate in categoria ²selfish DNA² ("ADN egoist"), totusi si aceste plasmide sunt mentinute intr-o anumita proportie din populatiile bacteriene.
In ansamblu, genomul bacterian se caracterizeaza printr-o intensa circulatie pe orizontala a genelor, intre diversi indivizi bacterieni (H.Tschape 1994). Aceasta circulatie se desfasoara prin trei procese centrale - conjugare, transformare si transductie - dintre care, plasmidele sunt implicate in primele doua.
Acest flux continuu de gene pare sa fie o caracteristica definitorie a lumii bacteriene si sa fi contribuit esential la extrema versatilitate metabolica a bacteriilor in aproape toate mediile naturale.
Plasmidele pot fi clasificate functie de diverse criterii :
a) criteriul autotransferabilitatii. Autotransferabilitatea reprezinta capacitatea unui element genetic (in speta, un plasmid) de a se autotransfera de la un individ bacterian la altul. De regula, genele implicate in autotransferabilitate sunt grupate intr-un operon (denumit operon tra), iar elementul genetic purtator este denumit conjugon. Conform acestui criteriu exista 3 clase de plasmide :
plasmide conjugative - sunt plasmidele care prezinta intreg operonul tra si sunt autotransferabile, putand genera proces de conjugare bacteriana;
plasmide non-conjugative - sunt plasmidele ce nu prezinta operonul tra, nu sunt autotransferabile (nu pot genera conjugare bacteriana) si nici mobilizabile;
3) plasmide mobilizabile - sunt plasmidele ce nu prezinta operonul tra (si, ca atare, nu sunt autotransferabile), dar au gene mob care le permit mobilizarea in evenimente de conjugare initiate de alte plasmide ce sunt conjugative.
b) criteriul structurii moleculare. Marea majoritate a plasmidelor sunt formate din ADN dublu catenar. Mai mult decat atat, din punct de vedere al structurii moleculare, exista doua clase de plasmide :
1) plasmide circulare. Plasmidele circulare sunt alcatuite din ADN d.c. circular, iar forma in vivo pare sa fie CCC ("circular covalently closed"); foarte rar si temporar se intalneste forma CO ("circular open"), in care una din cele doua catene nu este continua avand o legatura fosfodierica lipsa. Tot in vivo se mai intalnesc si concatemeri (oligo/multimeri).
2) plasmide lineare Asemenea plasmide sunt alcatuite din ADN d.c. linear (L). Au fost descrise doua tipuri de plasmide lineare :
Plasmide lineare cu telomere hairpin (telomere in ac-de-par). Acestea sunt intalnite cu precadere la plasmidele lineare de la Borrelia (Figura 3.3). Spirochetele din genul Borrelia au structura genomica lineara: toate speciile de Borrelia examinate au un cromosom linear de 950-1000 kb si mai multe plasmide lineare cu dimensiuni cuprinse intre 5 si 200 kb (unele au si plasmide CCC). Toate plasmidele lineare de la Borrelia descrise pana in prezent au telomere de tip hairpin. Telomerele hairpin se caracterizeaza prin palindroame terminale bogate in A/T, la capete existand o continuitate in legatura de tip covalent intre cele 2 catene. Asemenea structuri (adn d.c. L cu telomere de tip hairpin) mai exista si la ADNmt de la reprezentantii genului Pichia, la poxvirusuri si la plasmidele lineare din mitocondriile de fungi.
2. Plasmide cu telomere invertroni Asemenea telomere sunt intalnite la plasmidele lineare de la genul Streptomyces - plasmide cu dimensiune cuprinsa intre 9 si 600 kbp. Ca si cele hairpin, telomerele invertroni prezinta si ele palindroame terminale bogate in A/T. Diferenta consta insa in faptul ca nu exista legatura covalenta intre capetele celor 2 catene, si, ca urmare, capetele 5' si 3' sunt libere. La capul 5' al fiecarei catene este atasata covalent o molecula proteica denumita proteina TP ("Telomeric Protein") (Figura 3.3). Asemenea structuri ADN, cu palindroame terminale bogate in A/T si cu proteina TP atasata covalent la capul 5' au fost definite de K.Sakaguchi (1990) ca fiind elemente genetice mobile cu replicare autonoma si denumite invertroni, similare functional cu transposonii.
Telomerele invertroni sunt caracteristice repliconilor lineari de la eucariote, in mod special la plasmidele lineare din mitocondriile de fungi si plante si din citoplasma de fungi.
|
a)
|
Figura 3.4 Structura telomerelor hairpin la Borrelia (a) si invertroni la Streptomyces (b) (dupa H.Hinnebusch, 1992). Þ palindroame terminale, bogate in A-T
|
|
b)
|
c) criteriul incompatibilitatii plasmidiale
De-a lungul anilor, conceptul de incompatibilitate plasmidiala a ridicat foarte multe probleme. Actualmente, acest concept este definit ca fiind incapacitatea a doua sau mai multe plasmide de a se mentine si propaga stabil in aceeasi linie celulara. Plasmidele din acelasi grup de incompatibilitate ce contin origini de replicare similare (deci, au acelasi replicon-type), nu sunt mostenite stabil in descendenta si segrega una de cealalta. Cel mai probabil acest proces se datoreaza unei competitii moleculare intre plasmide cu acelasi replicon-type pentru aceleasi proteine implicate in replicarea respectivelor plasmide - proteine codificate cromozomal. La Enterobacteriaceae au fost descrise peste 30 de grupuri de incompatibilitate plasmidiala. Un tip de incompatibilitate s-ar baza pe competitia intre secventele repetate RS (la care se ataseaza proteina RepA), dintre diferite plasmide existente in celula bacteriana. Bazandu-se pe tipul de replicon a fost imaginata o metoda noua de identificare plasmidiala folosind o colectie de sonde rep (Sykora P. 1992).
Procesul de ²gene-flow² in lumea bacteriana ar putea avea un rol important in evolutia bacteriilor. Cu toate acestea, procesele de transfer de material genetic pe orizontala nu par sa afecteze stabilitatea speciilor taxonomice bacteriene. Din acest punct de vedere, plasmidele se gasesc intr-o situatie total diferita: ²gene-flow²-ul intre plasmide neinrudite pare sa fie atat de ridicat si de raspandit, incat nu se poate alcatui o identificare taxonomica a plasmidelor doar pe baza unei liste a caracterelor pe care le codifica. Se pare ca biologia plasmidelor nu implica mecanisme de izolare taxonomica asemanatoare cu speciile de plante si animale. In contrast, in lumea plasmidelor se observa un fenomen total diferit: izolarea plasmidelor individuale sau foarte asemanatoare. Acest fenomen este cunoscut ca fenomenul de incompatibilitate plasmidiala.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3037
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved