| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Premisele stiintifice ale ingineriei genetice la nivel molecular
Ingineria genica moleculara este un domeniu multidisciplinar, al carui scop este dezvoltarea bioindustriei in general si a microbiologiei industriale in particular. Evolutia sa pana la stadiul de corp unitar de metode si tehnici proprii este rezultatul descoperirilor in domeniul biochimiei enzimelor si acizilor nucleici de origine bacteriana. Progresele stiintifice ale microbiologiei din perioada l950-l965, au avut un impact hotarator asupra dezvoltarii biologiei in general, dar in special asupra dezvoltarii conceptelor biologiei moleculare. Termenul de "biologie moleculara" a fost introdus in l950.
Biologia moleculara nu este o disciplina de sine statatoare, ci reuneste intr-un corpus stiintific unitar, domenii care anterior erau complet independente (Microbiologia, Genetica, Biochimia). Biologia moleculara este un mod de a intelege desfasurarea proceselor biologice. Ea explica proprietatile si activitatile organismelor pe baze moleculare, aplicand sistemelor biologice, legi care guverneaza sistemele mai simple (fizice si chimice).
Orientarea moleculara a cercetarilor biologice, cristalizata in timp, a beneficiat de aportul Microbiologiei si Virologiei, domenii care au furnizat modelul experimental al cuplului E. coli - fag lambda. In acest context, Microbiologia a avut rolul unificator, deoarece diferitele tehnici (fizice, biochimice, genetice) si rezultatele obtinute au fost raportate la acelasi model experimental, facand posibila interpretarea lor corelativa. Sistemul biologic E. coli - fag lambda a devenit cel mai bine cunoscut din intreaga lume vie, fiind cunoscute toate detaliile de structura, precum si amplasarea pe cromosom a circa 800 de gene. Biologia moleculara este rezultatul utilizarii convergente a mai multor tehnici experimentale de varf, pentru studiul aceluiasi model experimental. Consecinta a fost consolidarea biologiei moleculare, concretizata in cateva realizari stiintifice fundamentale:
elucidarea structurii acizilor nucleici si a proteinelor;
descoperirea codului genetic
descoperirea fluxului informatiei genetice in celula;
descoperirea mecanismului molecular al biosintezei proteinelor;
descoperirea proceselor de variabilitate, de recombinare si de mutageneza;
descoperirea bazelor moleculare ale proceselor de crestere si diviziune celulara;
descoperirea mecanismelor de reglare a activitatii celulare.
Aceste descoperiri stiintifice de exceptie, au justificat definirea perioadei l950-'70, in termenii revolutiei biologiei moleculare. Conceptele majore ale celor mai multe stiinte traditionale s-au reformulat pe baze moleculare. Dupa l970, doua categorii de descoperiri stiintifice au influentat decisiv evolutia biologiei moleculare:
l) Descoperirile care au infirmat o serie de dogme:
dogma centrala a biologiei moleculare, odata cu descoperirea reverstranscriptazei si a transcrierii inverse (la retravirusuri);
dogma "o gena - o enzima", dupa evidentierea genelor suprapuse si a citirii defazate, la virusurile de dimensiuni mici;
dogma citirii secventiale a informatiei genetice (citirea fara pauze, odata cu descoperirea genelor discontinui);
2) Descoperirea enzimelor de restrictie si a plasmidelor a deschis calea elaborarii unor tehnici si tehnologii de biologie moleculara, cunoscute astazi sub denumirea de inginerie genetica, geniu genetic sau tehnologia ADN recombinant. Tehnicile de inginerie genetica se bazeaza pe posibilitatea sectionarii moleculei de ADN, de diferite origini (animala, vegetala, fungica, bacteriana, virala), cu ajutorul unor enzime speciale denumite endonucleaze de restrictie si a legarii reasortate a unor fragmente, prin intermediul actiunii altor enzime, denumite generic ADN-ligaze, in vederea transferului in alte celule.
Exista posibilitatea ca genele transferate, sa functioneze in celula acceptoare si sa determine sinteza de catre celula respectiva, a unor substante noi, pe care, in mod obisnuit, celula nu le produce. Microorganismele, carora in mod artificial li se grefeaza gene straine, functionale sunt reprogramate genetic.
Un interes deosebit pentru tehnicile de inginerie genetica, l-a starnit posibilitatea sintezei de catre celulele de E. coli reprogramate, a unor molecule de o deosebita importanta clinica: insulina, somatostatina, interferonii etc.
Tehnicile bazate pe metodologia ADN recombinant, au deschis calea unei noi revolutii - revolutia biotehnologica sau bioindustriala. Revolutia biotehnologica este ferm orientata spre industria genetica, deoarece utilizeaza, pe scara tot mai larga, microorganisme modificate genetic (reprogramate) prin tehnici de inginerie genetica.
Revolutia biotehnologica este considerata ca una din perspectivele de mare succes ale studiului microorganismelor. La baza ei stau descoperirile facute, in primul rand, pe celulele de E. coli. Cunoasterea aprofundata a celulei de E. coli a conditionat, ca majoritatea transferurilor de gene sa se faca la acest organism, care a devenit astfel un adevarat "cobai" al biologiei moleculare.
Enzimele de restrictie
Pana in l960, izolarea unei gene cromosomale parea imposibila. Prin procedeul fragmentarii mecanice a ADN, obtinerea unui fragment de lungimea unei singure gene din genomul mamiferelor are o sansa foarte mica (l/l00 000). Purificarea unui astfel de fragment este imposibila.
Solutia practica a izolarii genelor a fost adusa de enzimele de restrictie, care au activitate de endonucleaze. Studiul lor a inceput odata cu descoperirea faptului ca unele bacterii au capacitatea de a rezista infectiei fagice, deoarece poseda enzime care ataca si cliveaza ADN de provenienta exogena (straina).
Enzimele de restrictie se gasesc la cele mai multe microorganisme procariote, dar lipsesc la eucariote. Diferenta neta a distributiei lor este partial explicata, prin aceea ca la procariote exista un flux genetic deosebit de intens, fara echivalenta in restul lumii vii. Enzimele de restrictie au rolul de a recunoaste si de a degrada ADN exogen, limitand replicarea sa si propagarea in generatiile celulare ulterioare.
Enzimele de restrictie au urmatoarele proprietati definitorii:
recunosc ca straine secventele din molecula de ADN exogena si o cliveaza, facand-o nefunctionala, fiind enzime cu activitate endonucleazica;
enzimele de restrictie sunt asociate in celula bacteriana, cu enzimele de modificare. Ultimele recunosc aceleasi secvente de ADN strain si le modifica, astfel incat nu mai pot fi clivate de endonucleaze.
Enzimele de restrictie-modificare formeaza un sistem functional unitar si au rolul de "santinele genetice" ale celulei. Ele recunosc aceiasi secventa de ADN, dar realizeaza activitati enzmatice diferite.
Se cunosc trei clase de enzime de restrictie, notate cu I, II si III. Molecula de enzima de restrictie de tip I cumuleaza functiile de nucleaza (produce clivarea) si de modificare prin metilare. Trei situsuri distincte ale aceleiasi molecule realizeaza functiile de recunoastere, clivare si modificare. Fragmentele de ADN rezultate sunt heterogene ca marime si nu pot fi utilizate pentru clonare sau pentru analiza structurala a ADN.
Restrictazele de tip II au numai functie endonucleazica si cliveaza molecula de ADN in fragmente de dimensiuni relativ egale, utilizabile pentru clonare.
Restrictazele de tip III se deosebesc de cele de tip I, prin aceea ca nu au functia de modificare. Fragmentele de ADN care rezulta, sunt heterogene.
Nucleazele taie legaturile fosfodiester ale lanturilor polinucleotidice. Cele ce taie preferential legaturile interne se numesc endonucleaze, iar cele care sectioneaza nucleotidele terminale se numesc exonucleaze.
Mecanismul de actiune. Mult timp dupa descoperirea lor, s-a considerat ca endonucleazele actioneaza nespecific, adica cliveaza molecula de ADN in orice punct, indiferent de secventa bazelor. Endonucleazele de restrictie cliveaza fragmentele de ADN numai la nivelul unor secvente de 4-8 baze, care au caracter de palindrom, adica ordinea bazelor in directia 5' -- 3' pe o catena este aceiasi si pe catena opusa in aceiasi directie 5' --- 3'.
De exemplu, restrictaza Eco R1 recunoaste secventa*:
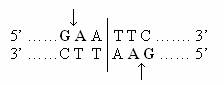
Situsul de actiune a enzimei, pe cele doua catene este decalat, fiind localizat intre nucleotidele marcate cu simboluri in relief.
Deoarece aceiasi secventa de baze, dar orientata in directii opuse se gaseste pe ambele catene, enzimele de restrictie creeaza totdeauna clivari dublu catenare. Ruperile se produc aproape simultan, deoarece multe enzime de restrictie, sub forma dimerilor, se leaga de ambele situsuri de recunoastere pe cele doua catene. Adeseori, situsul de recunoastere este si situs de clivare. Daca cele doua catene ale moleculei de ADN sunt sectionate la acelasi nivel rezulta fragmente cu capete taiate drept (blunt ended). Alte enzime sectioneaza decalat pe cele doua catene, la o oarecare distanta de axul de simetrie a palindromului si astfel rezulta fragmente de ADN cu extremitati monocatenare lipicioase (sticky ends), care se pot lega prin complementaritate si genereaza structuri moleculare circulare.
Palindroamele recunoscute de enzimele de restrictie sunt relativ scurte. Discontinuitatile create nu sunt reparate de enzimele de reparare.
Se cunosc peste l00 de endonucleaze cu specificitati diferite, izolate de la peste 300 de specii de microorganisme procariote. Pentru ingineria genetica, aceste enzime sunt instrumente esentiale de lucru, adevarate "bisturie biochimice", care sectioneaza moleculele de ADN. Din moleculele mari de ADN se obtin fragmente mici. Secventele scurte de diferite origini pot fi legate sub actiunea ADN-ligazei.
Enzimele de modificare
Modificarea este procesul biochimic prin care, unele enzime specifice sintetizate de bacterii modifica natura bazelor la nivelul unor secvente de ADN, recunoscute de enzimele de restrictie. Secventele modificate devin rezistente la actiunea endonucleazelor de restrictie.
Pentru fiecare endonucleaza de restrictie trebuie sa existe o enzima de modificare. Cel mai adesea, modificarea se realizeaza prin metilare la situsuri specifice sub actiunea metilazelor. De exemplu, restrictaza Eco R II recunoaste secventa:
G C A G G ...
G G T C C ....
Enzima de modificare metileaza citozina:
m
C C A G G ...
G G T C C ...
m
O secventa de nucleotide poate fi substratul actiunii unei enzime de restrictie sau de modificare, dar niciodata pentru ambele.
La procariote, circa l% din totalul bazelor sunt metilate. Cel mai adesea este metilata adenina (sub forma metil-adeninei) si cu o frecventa mai mica se metileaza citozina (sub forma metil-citozinei). Modificarea prin metilare se realizeaza pe ambele catene ale moleculei de ADN, in cursul replicarii acesteia.
Simultan cu enzimele de restrictie-modificare s-au descoperit alte categorii de enzime:
ligazele, enzime capabile sa adauge nucleotide la extremitatea moleculei de ADN, sa lege covalent cele doua extremitati si sa formeze o molecula lineara recombinata sau respectiv, o molecula circulara. Cea mai utilizata ligaza comerciala este cea codificata de fagul T4;
terminal-transferazele, enzime care catalizeaza adaugarea nucleotidelor la extremitatea 3' a moleculei de ADN in curs de sinteza, fara sa necesite matrita si chiar pot sa creeze extremitati adezive.
Restrictazele, ligazele si terminal-transferazele sunt instrumentele de lucru ale ingineriei genice. Restrictazele taie fragmentele de ADN, iar ligazele reunesc capetele rezultate. Foarte important si definitoriu pentru ingineria genica, este faptul ca fragmentele de ADN rezultate sub actiunea unei restrictaze pot fi reunite (recombinate), indiferent de provenienta lor. Se obtin astfel molecule himere.
Moleculele recombinate de ADN stau la baza restructurarii profunde a continutului microbiologiei industriale.
Cea de a II-a categorie importanta de molecule pentru biotehnologie sunt plasmidele. Ele au capacitatea de a incorpora in structura lor, gene de provenienta straina si de a le transmite cu o frecventa mare, de la o celula la alta.
In al II-lea rand, biotehnologia se bazeaza pe perfectionarea in vitro, a tehnicilor de transfer de material genetic la bacterii si la levuri, prin care se modeleaza mecanismele de transfer care au loc in mod natural.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 1369
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved