| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
REGLAREA EXPRIMARII GENELOR LA EUCARIOTE
La eucariote reglarea genetica are un caracter mult mai complex deoarece
materialul genetic este complexat cu histone, pentru a forma fibra de cromatina;
reglajul genetic la eucariote este mai complex din cauza existentei genelor in mozaic, astfel ca sinteza ARN se realizeaza in mai multe etape prin care se elimina intronii;
reglajul genetic la eucariote este afectat de faptul ca sinteza proteica se realizeaza in citoplasma, iar ARNm trebuie sa migreze din nucleu in citoplasma, la locul sintezei proteice;
reglajul genetic la eucariote are un caracter mai complex din cauza unei cantitati foarte mari de ADN in nucleu, din care insa numai o parte este informational.
La eucariote, genele nu sunt organizate in operoni, motiv pentru care reglajul genetic se realizeaza la nivelul genelor individuale. Ca urmare, fiecare molecula de ARNm poarta mesajul genetic pentru o singura catena polipeptidica el fiind monocistronic.
ADN de la eucariote este permanent complexat de histone care, pe langa rolul lor structural de a asigura stabilitatea fibrei de cromatina, joaca si rol de represori nespecifici generalizati, astfel ca genele se afla intr-o permanenta stare represata. Pentru a functiona, genele trebuie sa fie induse sa functioneze, atunci cand necesitatile celulei o cer. Activarea genelor se realizeaza cu ajutorul proteinelor nonhistonice. Deoarece la eucariote majoritatea genelor sunt inactivate in orice moment, reglajul genetic se realizeaza nu prin blocarea activitatii lor (represie) ci prin activarea unor anumite gene (inductie), activarea si inactivarea avand un caracter reversibil.
Reglarea activitatii genelor se poate realiza la mai multe nivele, evidente in cursul citodiferentierii:
Majoritatea genelor de la eucariote sunt inactive (tacute) pana in momentul in care sunt activate in mod specific. O prima conditie pentru ca procesul de transcriere sa poata fi initiat este ca nucleosomii sa poata fi indepartati de la nivelul regiunii de interes.
Histonele, la randul lor, pot sa influenteze proprietatile de transcriere ale ADN prin modificari chimice (fosforilare, acetilare, metilare) afectand astfel interactiunea ADN- histone. Proteinele nonhistonice indeplinesc rolul unor activatori specifici ai genelor asigurand transcrierea diferentiata a genelor , ele interactionand cu histonele pe care le indeparteaza de la nivelul genelor ce rmeaza a fi transcrise.
Reglarea transcrierii genelor la eucariote se mai poate realiza si prin procesul de metilare al ADN. Astfel, imediat dupa replicare, o parte din bazele azotate (in special citozina) sufera un proces de metilare (prin interventia unei metil transferaze). Gradul de metilare al citozinei variaza in functie de specie. Astfel, la mamifere de exemplu, 70 % din ADN este metilat.. S- a constatat ca ADN-ul genelor inactivate este mai puternic metilat decat cel al genelor active. Metilarea citozinei poate marca macromolecula de ADN in mod stabil, el fiind transmisa celulelor fiice la fiecare diviziune si poate indeplini functii multiple, incluzand mentinerea structurii cromosomului, controlul transcriptiei, transformarea oncogenica etc. Metilarea unor anumite secvente de ADN este asociata cu inhibitia transcriptiei, genele respective fiind insa inactivate in mod reversibil.
Datorita structurii discontinue a genelor (exoni + introni), se sintetizeaza ARNm precursor (premesager) ce contine atat secvente informationale cat si secvente non-informationale. Prelucrarea acestei molecule (splicing) pentru a obtine ARNm matur, functional presupune parcurgerea mai multor etape in care sunt implicate molecule de ARNsn precum si anumiti factori proteici.
S-a evidentiat faptul ca in anumite situatii prelucrarea ARNm se poate realiza intr-o asemenea maniera incat permite o asortare variata a ezonilor din aceeasi gena, proces denumit prelucrare alternativa (alternative splicing).
3. Reglajul genetic la nivelul transportului ARNm in citoplasma. Odata prelucrat, ARNm migreaza din nucleu in citoplasma, la nivelul porilor din membrana nucleara. Cercetarile au aratat ca numai o mica parte din ARNm sintetizat in nucleu, migreaza in citoplasma, cea mai mare parte din ARNm fiind degradat in nucleu cu ajutorul unor enzime. Reglajul genetic este acela care decide ce secvente de ARNm vor fi degradate in nucleu si care vor fi exportate in citoplasma, acolo unde se realizeaza sinteza proteica. Este vorba de enzime diferite ce opereaza in tipuri variate de celule, fapt ce determina ce sinteze proteice caracteristice vor realiza celulele respective.
4. Reglajul genetic la nivelul translatiei mesajului genetic. Consta in faptul ca nu toate moleculele de ARNm matur migrate in citoplasma vor fi utilizate in procesul sintezei proteice.
5. Rglajul genetic la nivelul degradarii ARNm are rolul de a selecta moleculele de ARNm matur migrate in citoplasma care vor fi degradate. In cazul genelor hemogobinei, de exemplu, ARNm are o mare stabilitate in timp, astfel ca el serveste repetat pentru sinteza proteica.
De asemenea, la eucariote se poate spune ca exista doua tipuri de reglare a activitatii genelor: reglare pe termen scurt- se bazeaza pe mecanime moleculare reversibile, reprezentate de modificari in activitatea unor gene, si reglare pe termen lung- care este de regula ireversibil si implica fenomene legate de diferentierea celulara ce au loc in cursul dezvoltarii ontogenetice.
La eucariotele superioare, reglarea pe termen scurt (la nivelul transcrierii genetice), este in mare parte mediata de hormoni. Un anumit hormon poate avea drept tinta una sau mai multe tipuri de celule, fiecare tip raspunzand diferit la acelasi hormon. Unii hormoni regleaza activitatea genelor influentand transcrierea, traducerea sau functionarea unor enzime cum este adenilat ciclaza. De exemplu, cei mai multi hormoni polipeptidici isi exercita efectele lor initiale la nivelul membranei celulelor tinta, stimuland activitatea adenilat- ciclazei. Aceasta converteste ATP la AMPc capabil sa stimuleze sinteza genelor, ca si in cazul procariotelor. Hormonii steroizi actioneaza direct la nivelul transcrierii genetice, fara interventia AMPc, in timp ce alti hormoni pot afecta histonele stimuland astfel procesul de transcriere.
Reglarea pe termen lung se realizeaza prin heterocromatinizarea sau condensarea mai pronuntata a cromatinei ceea ce conduce la o reducere sau chiar o blocare a functionarii ADN ca matrita pentru sinteza ARN.
La nivelul cromosomilor eucariotelor exista doua tipuri de heterocromatina, constitutiva si facultativa, care sunt implicate in activitatea genelor.
Astfel heterocromatina constitutiva se gaseste la nivelul regiunilor care nu sunt niciodata exprimate (transcrise); acestea includ satelitii si au un rol structural pentru cromosom. Deseori, secventele condensate sunt concentrate la nivelul unor regiuni specifice, de obicei in jurul centromerului, ele pastrand aceeasi configuratie in toate celulele organismului respectiv.
Heterocromatina facultativa reprezinta cromatina condensata doar in anumite celule, in timp ce in alte tipuri de celule regiunile corespunzatoare sunt decondensate. S-a dovedit faptul ca, in celulele embrionare cantitatea de heterocromatina facultativa este mica in timp ce in celulele specializate proportia sa creste foarte mult. Aceasta inseamna ca in cursul procesului de dezvoltare individuala (ontogenetica) are loc inactivarea prin heterocromatinizare a unui numar mare de gene.
Cel mai cunoscut exemplu de reglare prin heterocromatinizare de tip facultativ este cromatina sexuala ce reprezinta unul dintre cromosomii X de la femelele de mamifere care se inactiveaza la intamplare (fie cel de origine materna, fie cel de origine paterna), in mod definitiv.
Astfel, organismul femel poate fi considerat ca fiind un mozaic de clone celulare, fenomenul fiind evident in cazul unor gene mutante localizate pe cromosomul X:
in cazul unor celule gena mutanta se manifesta fenotipic, in timp ce in altele nu (cele ce contin cromosomul X heterocromatinizat).
Un exemplu este cel al pisicilor "calico" la care femelele au blana cu pete galbene si negre, fiind heterozigote (Cy/CB), genele pentru culoarea blanii fiind localizate pe cromosomii X.
In cursul dezvoltarii ontogenetice in unele celule este inactivat cromosomul X pe care se gaseste gena Cy in timp ce in alte celule se inactiveaza cromosomul X ce contine gena CB.
Rezultatul este aparitia unor pisici cu blana cu pete galbene si negre. La masculi, din cauza ca exista un singur cromosom X, fenomenul nu se realizeaza, ei fiind fie complet negri fie galbeni

Exemplul de mai sus este un tip de Reglare pe termen lung care se realizeaza prin heterocromatinizarea sau condensarea mai pronuntata a cromatinei ceea ce conduce la o reducere sau chiar o blocare a functionarii ADN ca matrita pentru sinteza ARN.
Heterocromatina, puternic condensata, este prezenta mai ales in celulele inalt diferentiate care sintetizeaza foarte putine proteine comparativ cu celulele nediferentiate.
De exemplu, in leucocitele normale apar mase mari de heterocromatina sub forma unor blocuri puternic condensate la periferia nucleului, in timp ce in cazul leucemiilor aceste aspecte sunt absente, celulele trecand in starea nediferentiata cu sinteze proteice intense si capabile de diviziuni celulare rapide.
Heterocromatinizarea poate avea loc la nivelul unor segmente cromosomale, a unor cromosomi intregi (X de la femelele de mamifere) si chiar a intregului genom (ex. La Planococcus citri), cand cromosomii de origine paterna sunt inactivi, formarea spermatozoizilor realizandu-se in urma diviziunii mitotice, ei fiind de 2 tipuri: unii functionali si altii nefunctionali (contin cromosomii paterni inactivi).
Un alt aspect al reglajului genetic s-a identificat odata cu descoperirea genelor cu structura discontinua de la eucariote. In acest caz, intr-o prima etapa este sintetizat un ARNm primar (ARN premesager) care contine mai multi exoni si introni dupa care, prin prelucrarea sa se poate forma ARNm matur care include toti exonii sau numai o parte dintre ei, proteinele rezultate fiind evident diferite (ex. Cazul formarii anticorpilor).
Citodiferentierea
Procesul de diferentiere celulara sau citodiferentiere este specific organismelor eucariote pluricelulare, el referindu-se la modificarile structurale si functionale pe care le sufera celulele ce rezulta (in urma unor diviziuni mitotice repetate) din celula-ou sau zigot.
La organismele eucariote, continutul in ADN este acelasi, indiferent de tipul celular, ceea ce inseamna ca diferentierea celulara si dezvoltarea organismului reprezinta rezultatul unei cascade de evenimente programate genetic ce determina activarea sau represia genelor.
Studiile efectuate asupra dezvoltarii ontogenetice la Drosophila melanogaster si la nematodul Caenorhabditis elegans au permis elucidarea unor aspecte legate de genetica dezvoltarii.
Fenomenul diferentierii celulare se realizeaza intr-o ordine cronologica stricta, dupa un program extrem de riguros a carui nerespectare conduce la modificari anormale ale organismului.
De exemplu, la om unde exista aproximativ 1013 celule, in cursul dezvoltarii ontogenetice are loc diferentierea a peste 150 tipuri diferite de celule, deosebite ca forma, marime, structura sau functie.
Studiul diferentierii celulare la mamifere este dificil, motiv pentru care cercetatorii au ales un sistem model la care dezvoltarea embrionara sa poata fi urmarita cu usurinta.
Acesta este cazul dezvoltarii embrionare la Drosophila melanogaster,
avantajele alegerii datorandu-se urmatoarelor caracteristici: genom
de dimensiuni mici numar
mic de cromosomi (patru perechi) si existenta cromosomilor politeni prezinta
mai multe stadii larvare, dezvoltarea fiind prin metamorfoza
(ou-larva-pupa-adult), intregul proces de metamorfoza (dezvoltarea de la ou
la adult) dureaza aproximativ noua zile.
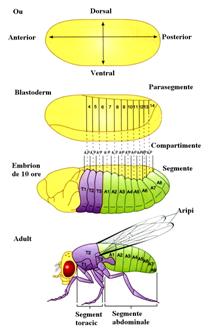
Reglarea procesului de citodiferentiere, aceasta se realizeaza la niveluri diferite:
la nivelul cantitatii de ADN si a replicarii materialului genetic, al transcrierii si traducerii la nivel cromosomal sau al intregului genom.
Astfel, desi se considera ca toate celulele unui organism contin aceeasi cantitate de ADN, s-a dovedit ca exista unele variatii ce se datoreaza unor procese de replicare diferentiata:
suprareplicare a anumitor regiuni cromosomale si subreplicarea altora (mai ales a regiunilor heterocromatice).
De exemplu, la plante exista diferente intre cantitatea de ADN din meristeme si cea din diferite organe: la nivelul celulelor meristematice cantitatea de ADN este mai mica in timp ce in alte organe, cum sunt cotiledoanele care sunt alcatuite din celule inalt diferentiate, cantitatea de ADN este mai mare.
In aceea ce priveste genele implicate in controlul dezvoltarii la D.melanogaster, acestea au fost identificate si studiate pe baza mutatiilor produse la nivelul lor, care au condus fie la aparitia unor structuri anormale fie la moartea organismului.
Aceste gene pot fi incluse in cel putin trei grupuri
gene de origine materna: se exprima in cursul ovogenezei si sunt responsabile de gradientul de proteine ce apare la nivelul oului si care sunt implicate organizarea spatiala a embrionului timpuriu
gene de segmentare: sunt exprimate dupa fecundare la nivelul zigotului si determina numarul si organizarea segmentelor corpului.
genele homeotice: se exprima dupa genele segmentare si determina identitatea fiecarui segment individual.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 4885
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved