| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
ROLUL MICROORGANISMELOR IN CIRCUITUL GLOBAL AL MATERIEI IN NATURA
Organismele fototrofe (plante, alge, bacterii fotosintetizante) sintetizeaza substante organice, pornind de la CO2, apa si saruri minerale, utilizand energia solara pe care o transforma in energie chimica.
Unele substante minerale au un rol biologic primordial (C, N, O, H, S, P, Na, K, Fe), iar altele sunt necesare numai in cantitati foarte mici (Mn, Mg, Zn, Cu, Co, Mo, B, etc.).
Biomasa produsa in fotosinteza este preluata partial de catre organismele heterotrofe. Resturile organice vegetale si animale se reintorc in sol si ape, dar sunt neutilizabile de catre plante. Solul pierde astfel permanent substante anorganice prin incorporarea lor in substanta organica. Se adauga pierderile determinate de eroziune, de depunerea unor compusi minerali in sedimente si roci sedimentare sau stocate sub forme ce evolueaza spre stadiul de combustibili fosili (carbune, titei). Continuarea acestui proces ar produce in timp, epuizarea formelor utilizabile, situatie incompatibila cu existenta vietii. Fenomenul epuizarii este inpiedicat de faptul ca microorganismele din sol si ape desfasoara permanent o puternica actiune de mineralizare a substantelor organice de provenienta vegetala sau animala.
Ciclurile biogeochimice insumeaza caile de circulatie a elementelor biogene in natura, prin care trec de la forma anorganica, la forma organica, pentru a reveni la forma anorganica din compartimentul abiotic. In ciclul biogeochimic intra in actiune reactii fizice (solubilizare, precipitare, volatilizare) si chimice (oxidari, reduceri, hidrolize), precum si activitati biologice (degradari si mineralizari, conversie organica prin biosinteze).
Ciclurile diferitelor elemente biogene sunt strans interconectate si interdependente, realizand in ansamblu, ciclul global al materiei in natura. De exemplu, bacteriile proteolitice, care descompun proteinele la CO2, NH3 si H2S sunt implicate in trei cicluri: al C, N si S.
Influenta omului perturba capacitatea proprie de echilibrare a unui ciclu, producand, uneori, modificari severe si greu reversibile ale mediului: cresterea concentratiei CO2 in atmosfera, ca o consecinta a arderii combustibililor fosili, poluarea sub diferite forme, cu un aport excesiv de materie si energie.
In circuitul materiei in natura, rolul microorganismelor este esential. Fara microorganisme n-ar putea exista plante, iar in lipsa lor, viata animalelor ar inceta. Microorganismele sunt tot atat de indispensabile vietii, ca si sursa de energie pe care o reprezinta soarele.
Azotul este un element esential pentru existenta vietii in biosfera, deoarece este inclus in structura tuturor proteinelor si acizilor nucleici. Desi reprezinta 79% din continutul atmosferei, azotul se gaseste aproape totdeauna in forme inaccesibile plantelor si animalelor.
Azotul atmosferic, diatomic (NsN) este un gaz inaccesibil majoritatii sistemelor biologice. Se adauga cantitati importante de azot organic din organismele vii si din humus, precum si din sedimentele din mari si oceane.
In circuitul azotului in natura, azotul mineral (NH4+, NO3-) este inglobat de plante sub forma constituientilor celulari. |esuturile vegetale si animale moarte sunt mineralizate in procesul de proteoliza si amonificare si convertite prin nitrificare, la forme din nou accesibile plantelor. Pierderile de azot din sol, legate de recoltarea plantelor de cultura, precum si de denitrificare si levigatie depasesc (pentru solurile cultivate) cantitatea de azot accesibil. De aici decurge necesitatea imbogatirii solului in azot combinat, fie pe cale naturala prin fixarea biologica a azotului atmosferic, fie artificial, prin adaugarea ingrasamintelor azotate. Se adauga azotul fixat pe cale abiotica prin iradieri, descarcari electrice din atmosfera si cel rezultat din procese de combustie (echipamente electrice, motoare cu combustie interna, etc.).
Circuitul biologic al azotului este un proces lent, care se desfasoara sub actiunea a peste 100 de genuri de bacterii. Azotul gazos atmosferic este convertit la forme fixe (NH4+, NO3-, NO2-), care sunt folosite de plante si incorporate sub forme organice in componente structurale.
Etapele esentiale ale circuitului azotului, unele desfasurate in aerobioza, iar altele in anaerobioza sunt: fixarea azotului molecular, amonificarea, nitrificarea (nitritarea si nitratarea), denitrificarea, toate mediate numai de bacterii.
Fixarea biologica a azotului este realizata in exclusivitate de bacterii chimiotrofe si fototrofe.
Din numarul total de 260 de genuri de bacterii chemotrofe, circa 10% contin nitrogenaza si au capacitatea de a fixa N2. Ele includ specii din genurile: Alcaligenes, Aquaspirillum, Arthrobacter, Azospirillum, Azotobacter, Bacillus, Beijerinckia, Campylobacter, Citrobacter, Clostridium, Derxia, Desulfotomaculum, Desulfovibrio, Enterobacter, Erwinia, Frankia, Klebsiella, Methylobacter, Methylosinus, Methylococcus, Mycobacterium, Propionibacterium, Rhizobium, Thiobacillus, Xanthobacter. Unele (Rhizobium, Frankia, Azospirillum) formeaza simbioze cu diferite grade de specificitate, iar celelalte sunt organisme libere, fixatoare de N2. Unele sunt aerobe (Azotobacter, Beijerinckia, etc.), altele anaerobe (Clostridium, Desulfovibrio, etc.), iar celelalte, in majoritate, aerobe facultativ anaerobe (Klebsiella, Citrobacter, Enterobacter).
Mulder si colab. (1965) considera ca bacteriile libere fixatoare de N2 au o contributie minora la economia globala a acestui element in natura si la fertilitatea solului: 2-3 kg N/ha/an.
Din cele 12 genuri de bacterii fototrofe identificate pana acum, 92% au capacitatea de a fixa N2: Chlorobium, Chromatium, Pelodictyon, Rhodospirillum, Thiocapsa, Thiocystis, etc. Ele fac fotosinteza anoxigenica si de aceea importanta lor este limitata la mediile anoxice, si anume la sedimentele din lacurile putin adanci.
Fixarea biologica a azotului in asociatie cu plantele este rezultatul unei cooperari mai mult sau mai putin intime cu planta gazda. In aceasta categorie sunt cuprinse: simbioza Rhizobium cu plante leguminoase si neleguminoase; simbiozele asociative; simbiozele foliare; actinorizele; cianobacteriile simbiotice fixatoare de N2, in asociatii cu fungii din clasa Ascomycetes sau Basiomycetes in licheni; cu muschii din grupul Hepaticae; cu pteridofitele in asociatia Azolla-Anabaena azollae; cu gimnospermele (Cycas, Ceratozamia, etc.); cu angiospermele (Gunnera sp.).
Simbioza Rhizobium-plante leguminoase este sistemul fixator cel mai eficient. De exemplu, capacitatea fixatoare a sistemului Rhizobium/lucerna sau Rhizobium/lupin poate reprezenta intre 350 si 600 kg N/ha/an.
Amonificarea
Cea mai mare parte a azotului din straturile superficiale ale solului este prezent sub forma de combinatii organice. El provine din proteinele vegetale sau animale, din biomasa microbiana, din excretele animale etc.
In climatul temperat, o rezerva majora de azot organic este asociata cu humusul si devine disponibila dupa mineralizare lenta. Extractele de sol contin, practic, toti aminoacizii existenti, legati in diferite combinatii organice si mici cantitati (2-400 ppb) ca aminoacizi liberi, glucide aminate, baze purinice si pirimidinice, etc.
Mineralizarea azotului organic din sol este un proces esential pentru ciclul acestui element in natura, deoarece asigura conversia la forme anorganice, utilizabile de catre plante si microorganisme.
Amonificarea evolueaza in doua etape:
- etapa nespecifica, de proteoliza, efectuata de microorganisme heterotrofe: Pseudomonas sp., Bacillus sp., Clostridium, Aspergillus, Mucor etc., carora li se adauga efectul proteazelor de origine vegetala sau animala. In aceasta etapa, moleculele organice complexe sunt hidrolizate la molecule mai mici care pot patrunde in celule, fiind utilizate de microorganisme sau sunt degradate in etapa urmatoare. Enzimele implicate in etapa nescpecifica sunt: endopeptidaze, care ataca lantul polipeptidic in interior, eliberand peptide de diferite lungimi. Unele endopeptidaze au actiune nespecifica, iar altele au actiune specifica deoarece recunosc anumite resturi de aminoacizi si hidrolizeaza legaturile peptidice din vecinatatea lor. Peptidazele sunt exopeptidaze (amino- sau carboxipeptidaze). Ambele tipuri de enzime elibereaza di- sau tripeptide, care sub actiunea di- sau tripeptidazelor elibereaza aminoacizi liberi. Acest proces intereseaza numai o mica fractie a azotului organic, restul fiind rezistent la degradare datorita formarii complexelor ligninoproteice sau datorita incorporarii in reteaua cristalina a argilelor minerale;
- etapa specifica este rezultatul actiunii microorganismelor din sol care asigura degradarea aminoacizilor, hidroliza acizilor nucleici si a ureii, a acidului uric, a glucidelor aminate si conversia lor la NH3.
In sens strict, amonificarea este procesul de eliberare a NH3, prin actiunea microorganismelor asupra moleculelor rezultate din descompunerea proteinelor, acizilor nucleici etc. Producerea si eliberarea NH3 sub actiunea microorganismelor de putrefactie nu este un proces de amonificare.
NH3 este rezultatul reactiilor de dezaminare, care se produc pe mai multe cai:
1) Dezaminarea oxidativa, descrisa la Proteus sp., Ps. aeruginosa, E. coli etc., dupa reactia:
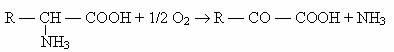
2) Dezaminarea reductiva, caracteristica aproape exclusiv bacteriilor anaerobe (Clostridium sp.), urmeaza calea:
![]()
3) Dezaminarea desaturanta, descrisa la E.coli, Proteus sp., Clostridium sp., Neurospora, evolueaza dupa reactia:
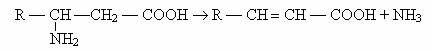
4)Dezaminarea prin deshidratare, descrisa la E. coli si N. crassa are loc printr-o serie de reactii cuplate:
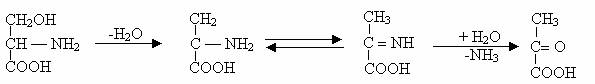
NH3 format, fiind volatil este partial eliberat in atmosfera, unde poate fi expus transformarilor fotochimice sau reactioneaza direct cu oxizii formand sulfat de amoniu. Acestia pot reveni in sol sau in apa, odata cu ploile sau prin depuneri de particule.
O alta parte din NH3 este adsorbita temporar pe complexele argilo-humice, sau este convertit in conditii de aerohioza la NH4+, care este oxidat la nitriti si ulterior la nitrati.
Amonificarea este un proces lent, care evolueaza in conditii optime in straturile superficiale ale solurilor bine structurate si aerate, la pH apropiat de neutralitate.
In faza nespecifica, acizii nucleici proveniti din celulele vegetale, animale si din microorganisme sunt clivati in polinucleotide si in final, rezulta mononucleotide. Degradarea este catalizata de ribonucleaze, dezoxiribonucleaze etc., sintetizate de bacterii.
In etapa urmatoare, purinele si pirimidinele neutilizate de alte microorganisme sunt degradate de bacterii pana la stadiul de uree, care este supusa procesului de amonificare.
Amonificarea ureei este rezultatul actiunii microorganismelor. Ureea din sol provine din excretii, din metabolismul microorganismelor si din degradarea bazelor purinice si pirimidinice.
Conversia ureei la NH3 decurge dupa reactiile:
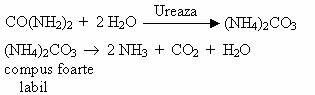
Activitatea amonificatoare a urobacteriilor este foarte importanta, deoarece ureea contine 47% N, care altfel ar ramane neutilizat de plante. Cantitatea de uree eliminata in natura este foarte mare (zeci de milioane de tone/an).
Amonificarea este o etapa esentiala a circuitului azotului in natura, prin care azotul organic este mineralizat in forma amoniacala, utilizabila ca atare sau dupa oxidarea la nitrati, in nutritia minerala a plantelor.
Nitrificarea
Nitrificarea este un proces biologic prin care NH3 sau alte forme reduse ale azotului anorganic, rezultate in procesul de amonificare sunt oxidate la nitrati, forma cea mai usor asimilabila de catre plante.
Nitrificarea este realizata de bacterii chemolitotrofe obligate, in doua etape succesive strans cuplate:
Bacteriile care oxideaza NH3 (nitrit-bacteriile sau nitrosobacteriile) sunt Gram negative (Nitrosomonas europea, Nitrosococcus nitrosus, Nitrosospira briensis, Nitrosolobus multiformis, Nitrosovibrio tenuis) si au sisteme membranare complexe, intracelulare.
Bacteriile care oxideaza nitritii (nitrat-bacteriile) sunt reprezentate de Nitrobacter winogradskyi, Nitrococcus mobilis, Nitrospira gracilis, N. marina.
Biochimia nitrificarii este relativ putin cunoscuta, deoarece bacteriile se dezvolta foarte greu pe medii artificiale (durata unei generatii este de 10 ore-cateva zile).
Se admite ca nitrificarea are loc dupa urmatoarele reactii:
1) NH4+ + 1/2 O2 NO2- + H2O + 2 H+ (Nitritarea)
2) NO2- + H2O NO3- + 2 H+ + 2e- (Nitratarea)
Nitritii apar constant in conditii de laborator ca produsi intermediari de nitrificare, dar existenta lor in sol nu a fost evidentiata. De aceea s-a presupus existenta unor bacterii care oxideaza NH3 direct la nitrati. In realitate, cele doua grupuri fiziologice de bacterii coexista in stransa asociere in sol si nitritii rezultati din oxidarea NH3 sunt utilizati imediat de nitratbacterii. Nitritii sunt toxici si de aceea, transformarea lor rapida are rol protector pentru organismele din sol.
Procesul nitrificarii este favorizat prin ameliorarea aerarii solului. Nitratii sunt usor levigati si ajung in panza de apa freatica, unde produc efecte negative: levigarea reprezinta o pierdere a azotului combinat din sol; nitritul reactioneaza cu compusi aminati din apa freatica si formeaza nitrozamine cancerigene; nitratul netoxic per se poate fi redus la nitriti de catre bacteriile din microbiota intestinala si determina fenomene toxice consecutive combinarii lor cu hemoglobina.
Denitrificarea
Denitrificarea este un proces biologic catalizat in exclusivitate de bacterii si consta in reducerea dezasimilatorie a unuia sau a ambilor oxizi ionici ai azotului (NO3- si NO2-), la oxizi gazosi, ca oxidul nitric (NO) sau oxidul nitros (N2O); acestia pot fi ei insisi redusi la N2. Simultan, are loc oxidarea materiei organice din mediu, cu producerea energiei.
Bacteriile denitrificatoare formeaza un grup biochimic si taxonomic heterogen, care reuneste peste 75 de genuri diferite. Unele sunt autotrofe si se dezvolta in prezenta CO2 si a H2 sau a compusilor redusi ai sulfului, dar cele mai multe sunt heterotrofe: Alcaligenes faecalis, Azospirillum sp., Pseudomonas aerogenes, P. denitrificans etc.
Nici o bacterie denitrificatoare nu este strict anaeroba. Ele prefera respiratia aeroba si au catena transportatoare de electroni.
Denitrificarea este mai activa in conditii de anaerobioza sau de tensiune scazuta a O2, in ape stagnante, in soluri umede sau inundate, in sedimente acvatice etc.
Biochimia denitrificarii este complexa si numai partial cunoscuta. Este un proces multienzimatic determinat de activitatea enzimelor nitrat-reductaza, nitrit-reductaza, oxidnitric-reductaza, care catalizeaza secventa de reactii:
NO3- NO2- NO N2O N2
Reducerea asimilatorie nitratilor este efectuata de un numar foarte mare de bacterii, fungi filamentosi, levuri, alge, plante superioare. Rezultatul este producerea de NH3, care nu se acumuleaza, deoarece este utilizat de celule pentru biosinteza constituientilor celulari.
Circuitul oxigenului
O2 reprezinta 21% din aerul atmosferic, dar se gaseste si intr-o serie variata de compusi organici si anorganici din organismele vii, precum si ca O2 "fosil", legat de zacaminte si depozite de combustibili fosili.
Acumularea de O2 liber in atmosfera, considerata cea mai profunda transformare biogeochimica, a inceput acum 1,8 miliarde ani. Fenomenul a avut o importanta covarsitoare, deoarece organismele multicelulare au evoluat numai dupa ce atmosfera s-a imbogatit cu acest element, prin fotosinteza. Cloud si Gilbert (1970) apreciaza ca cea mai mare parte, daca nu chiar tot O2 liber din atmosfera, este de origine biogena si provine prin fotoliza apei sub actiunea energiei luminoase, in cursul fotosintezei.
Oxigenul este produs prin actiunea plantelor verzi, a algelor si a procariotelor din grupul Oxyphotobacteria (Cyanobacteriales, Prochlorophytes si Halobacterium) in zona fotica a mediilor acvatice.
O parte din O2 eliberat in atmosfera este convertit sub actiunea radiatiilor ionizante la ozon (O3) si sustras din circulatia biogeochimica. O3 asigura functia de ecran protector, oprind UV daunatoare pentru organismele vii.
O alta parte din produsii primari ai fotosintezei sunt convertiti in biomasa vegetala - ca oxigen structural, component al moleculelor din organismele vii (glucide, aminoacizi, lipide, etc.). Oxigenul reprezinta 1/4 din atomii materiei vii.
In sfarsit, o parte din oxigen este scos din ciclul biologic ca parte a ionului carbonat (CO32-), fiind precipitat din solutie si stocat in rocile calcaroase.
Oxigenul provenit prin biofotoliza apei i se adauga cel provenit din surse "nebiologice" sub forma de CO2, apa si oxizi anorganici. Ionii nitrat si sulfat sunt surse de oxigen pentru unele organisme, care ii reduc NH3 si respectiv la H2S, care apoi sunt reoxidati, circuland in biosfera. Rezervele de oxigen cele mai circulate sunt cel atmosferic si dizolvat, din CO2 si din apa.
Rezervele de oxigen din materia organica vie sau neanimata, ca si cantitatile mari de oxigen din compusii minerali (inclusiv carbonati) sunt recirculate cu o rata foarte mica. Activitatile umane influenteaza ciclul CO2 in biosfera, prin combustia carbunelui, a petrolului si a gazelor care maresc continutul atmosferei in CO2.
Rolul microorganismelor in circuitul oxigenului in natura este mai evident in mediile acvatice. Apele repede curgatoare sunt bine oxigenate. Curentii oceanici asigura, in general, aportul de oxigen in apele oceanice si marine, cu exceptia apelor din golfuri, fiorduri sau din unele mari interioare.
O situatie speciala este aceea a unor bazine acvatice din regiunile temperate, in care vara are loc un proces de stratificare, determinat de temperatura. Apele mai calde de la suprafata sunt separate de straturile profunde situate sub termoclina (mai reci si mai dense). Straturile profunde devin anaerobe. Materia organica cade la fund, unde este folosita ca sursa nutritiva de organismele facultativ aerobe. Ele reduc si mai mult continutul in oxigen al apelor profunde. De aceea, in adancul acestor ape se gasesc numai microorganisme obligat anaerobe (cele care nu tolereaza concentratii de O2 mai mari de 1% din cele atmosferice) si numai uneori de microorganisme microaerofile. La inceputul iernii se realizeaza o "rasturnare" a straturilor de apa, care determina aerarea celor adanci: apele la suprafata devin mai reci si mai grele decat cele profunde si asigura omogenizarea conditiilor pe intreaga coloana de apa. Datorita acestor fenomene, unele lacuri din regiunile temperate prezinta un ciclu anual de trecere alternativa sezoniera a apelor profunde, de la aerobioza la anaerobioza si invers.
In raurile in care sunt deversate cantitati mari de substante organice sub forma de ape uzate sau de poluare industriala, concentratia oxigenului dizolvat scade semnificativ. Cantitatea mare de substanta organica determina cresterea excesiva a bacteriilor heterotrofe si un deficit de oxigen.
Circuitul carbonului in natura
Carbonul, element biogen esential pentru existenta sistemelor biologice, se gaseste in combinatii organice sau anorganice, repartizate in rezervoare cu circulatie rapida, precum si stocat in forme care circula foarte lent sau deloc.
Forma cea mai rapid circulanta este CO2 atmosferic, care reprezinta 0,032% (320 ppm), echivalent cu o cantitate globala de 70.000 tone C. In apa marii, C anorganic este prezent in solutie sub forma de CO2, H2CO3, HCO3- si CO32- sau precipitat sub forma de carbonati insolubili.
Atmosfera si hidro-ecosfera sunt strans legate, astfel incat asigura un schimb echilibrat de CO2. Se apreciaza ca in fiecare an, 100 miliarde tone de CO2 atmosferic sunt dizolvate in mare si o cantitate echivalenta de CO2 oceanic este transferata in atmosfera. Un echilibru asemanator caracterizeaza schimburile dintre litosfera si atmosfera: cantitatea de CO2 atmosferic fixata prin fotosinteza rezulta prin descompunerea substantelor organice [n etapa de mineralizare. Cu exceptia unor factori perturbatori de origine antropogena, concentratia CO2 atmosferic nu este expusa unor variatii semnificative.
Circulatia CO2 (atmosfera sol si atmosfera ocean), ca si circulatia CO2 C organic sunt foarte rapide, datorita faptului ca rezervorul de CO2 este limitat cantitativ. Dupa Stanier, Adelberg si Doudoroff (1970), in absenta recircularii, [ntreaga rezerva de CO2 ar fi fixata in fotosinteza in circa 20 de ani, sau dupa altii, in 40-50 de ani, dupa care orice forma de viata ar inceta sa mai existe.
Circulatia biogeochimica a C implica existenta a doua cicluri distincte: unul cu evolutie terestra si celalalt oceanic, interconectate dinamic la interferenta cu atmosfera.
Degradarea biologica a constituientilor vegetali
Substantele organice, componente ale peretului celular vegetal sunt polizaharidele, lignina si substantele accesorii nestructurale.
Polizaharidele sunt glucide cu greutate moleculara mare, cu rol predominat structural si sunt reprezentate de celuloza, hemiceluloze, substante pectice (cu rol de liant), manani, galactani, glucani si xilani etc (fig 164).
Lignina, cel mai putin definita structural dintre constituientii lemnului, reprezinta 20-35% din structura acestuia.
Substantele accesorii (nestructurale) sunt extractibile (terpeni, fenoli, rasini etc.) sau neextractibile (regasite in cenusi ca oxalati, carbonati, cristale de siliciu). Ele maresc rezistenta celulozei la degradare si la atacul insectelor.
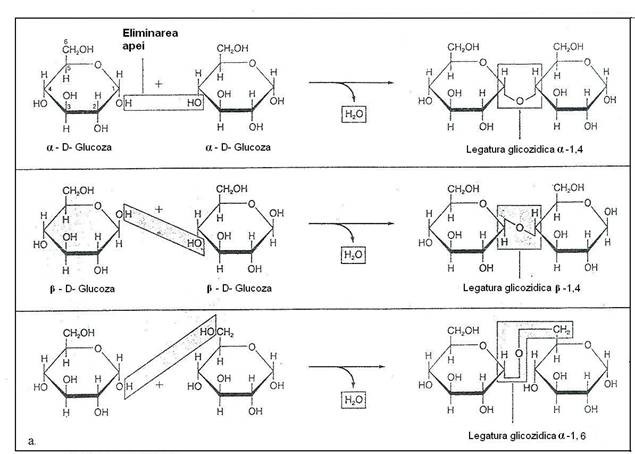
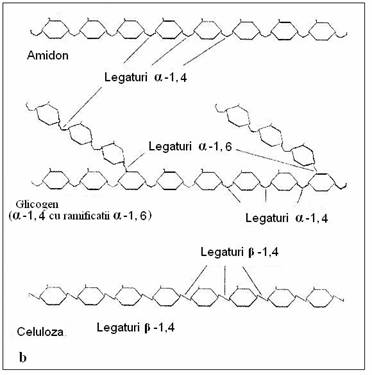
Fig. 164. a. Polimerizarea monozaharidelor. b. Structura polizaharidelor comune.
Structura si morfologia fibrelor de celuloza
Desi structura primara a celulozei este relativ simpla, structura sa tertiara (sau cuaternara) este foarte complicata, fapt care explica marea sa rezistenta la degradarea enzimatica.
In structurile naturale, circa 100 molecule de celuloza sunt legate impreuna pentru a forma fibrile elementare sau protofibrile. Ele au lungimea de circa 100 A, latimea de 40 A si grosimea de 30 A si sunt alcatuite din molecule de celuloza aliniate paralel si legate prin punti de H. Un numar de circa 20 ( 5) fibrile elementare se agrega intr-un fascicul lung si subtire numit microfibrila. Prin asocierea a circa 250 microfibrile rezulta o fibrila, iar din unirea a 1500 fibrile rezulta fibra macroscopica, vizibila cu ochiul liber.
De-a lungul microfibrilelor, moleculele de celuloza contin zone extinse cu organizare ordonata, corespunzand regiunile cristaline, ce alterneaza cu zone mai putin ordonate, numite paracristaline sau amorfe, usor atacate de enzime si agentii fizici si chimici.
O consecinta a modului de organizare foarte ordonata a fibrelor de celuloza este ca nici o molecula de apa sau de enzime nu poate intra in structura lor, motiv pentru care celuloza este inerta in intestinul multor animale. Pretratarea in vitro cu acizi sau alcali creeaza strucuturi deschise ce favorizeaza atacul enzimatic.
In celuloza nativa, regiunea cristalina poate reprezenta pana la 70% din lungimea microfibrilelor.
In plantele ierboase si in legume, celuloza reprezinta circa 15% din greutatea uscata, iar in plantele lemnoase, peste 50%.
Degradarea biologica a celulozei. Celuloza este constituient major al materialelor vegetale si cel mai abundent material organic prezent in natura. Degradarea biologica a celulozei are o importanta fundamentala pentru circuitul C in natura (fig. 165).
Din punct de vedere chimic, celuloza este un polimer linear, alcatuit din unitati repetate de celobioza (4-o-(b-D-glucopiramozil)-D-glucopiranoza, respectiv din 8000-12000 unitati de anhidro-D-glucoza, unite prin legaturi glicozidice 1,4-b-D. Numarul unitatilor si gradul de polimerizare sunt variabile in functie de provenienta moleculelor.
In conditii de laborator sau industriale se pot obtine celuloze modificate fizic sau chimic prin tratare cu acizi sau alcani, sau substituite chimic (de exemplu, trinitrofenil-celuloza sau carboximetil-celuloza).
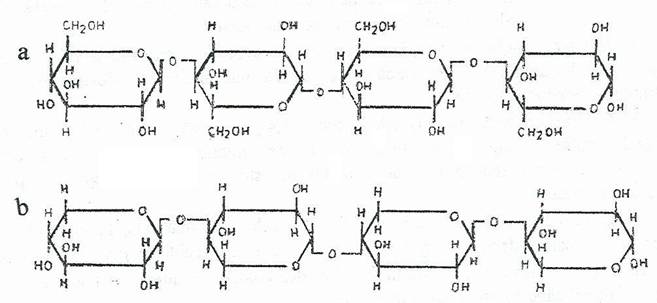
Fig. 165. Structura chimica a celulozei (a) si hemicelulozei (b).
Microorganisme celulozolitice
Peste 200 de specii de microorganisme (eubacterii, actinomicete, microfungi (levuri, si fungi filamentosi) si protozoare produc celulaze si degradeaza celuloza: Clostridium cellulovorans, C. cellulolyticum, C. thermocellum, Bacillus cereus, B. polymyxa, B. licheniformis, B. subtilis, Cellulomonas fini, C. flavigena, Cytophaga sp., Sporocytophaga, Aspergillus sp., Phoma, Trichoderma, Verticilium Alternaria, Botrytis, Cephalosporium, Chaetomium, Fusarium, Macrosporium, Penicillium, etc.
Toate microorganismele care degradeaza celuloza cristalina produc sisteme celulazice, formate dintr-o varietate de enzime care actioneaza cooperant. Sistemul enzimatic descris la Trichoderma reesi este alcatuit din trei tipuri majore de enzime:
1) Endo-b-1,4-glucanaza (EG) hidrolizeaza aleatoriu legaturile b-1,4-glicozidice din mijlocul moleculei de glucoza. Nu ataca celobioza, dar hidrolizeaza celodextrinele si celulozele substituite (carboximetilceluloza).
2) Celobiohidrolaza (CBH) este o exo-b-1,4-glucanaza. Actioneaza asupra celulozei, sectionand treptat unitati de celobioza de la extremitatea nereducatoare a catenei de polimer. Are o mare specificitate de substrat si este capabila sa degradeze peste 80% din celuloza cristalina.
b-glucozidaza hidrolizeaza celobioza si celooligozaharidele, la glucoza. Nu ataca celuloza si nici celodextrinele cu g.m. mare. Desi nu este o celulaza per se, ci, in special o celobioza, b- glucozidaza favorizeaza procesul de degradare enzimatica a celulozei. Hidrolizeaza celobioza, impiedicand acumularea ei in mediu.
Capacitatea de a degrada celuloza naturala implica sinteza intregului sistem enzimatic. Multe microorganisme pot degrada celuloza partial degradata, nu insa si celuloza nativa. Ele produc anumite enzime celulozolitice, dar le lipseste una dintre enzimele esentiale.
Hemicelulozele izolate din tesuturile vegetale, dupa tratamentul cu acizi minerali la cald elibereaza pentoze, hexoze si uneori, acizi uronici. Pentozele sunt reprezentate de xiloza si arabinoza, hexozele, de manoza sau galactoza, iar acizii uronici, de acidul glucuronic (C6H10O7) sau de acidul galacturonic (C6H10O5).
Hemicelulozele apar sub forma unor polimeri lineari alcatuiti dintr-o singura pentoza (pentozani) sau hexoza (hexozani). Pentozanii sunt xilani sau arabani, iar hexozanii sunt manani sau galactani. Hemicelulozele care contin acizi uronici se numesc hemiceluloze poliuronidice, iar cele lipisite de acestia, celulozani.
Cel mai important celulozan este xilanul, format exclusiv din 50-150 resturi de xiloza, legate β-1,4 sau contine cantitati mici de L-arabinoza.
Degradarea enzimatica a hemicelulozelor este realizata de enzime extracelulare si evolueaza in doua etape succesive:
- etapa conversiei moleculelor polimerice la oligoglucide sau la monomeri (xilotrioza, xilobioza, xiloza- in cazul xilanului);
- etapa utilizarii moleculelor simple, ca sursa de C si energie.
Dupa modul de actiune, hemicelulazele sunt de doua tipuri: unele xilanaze sunt de tip endo, deoarece scindeaza legaturile β-1,4 din interiorul catenei polizaharidice; altele sunt de tip exo si ataca polimerul la nivelul extremitatilor libere.
Microorganismele active sunt reprezentate de bacterii (Achromobacter, Bacillus, Cytophaga, Pseudomonas, Streptomyces, Vibrio, etc) sau fungi (Aspergillus, Chaetomium, Penicillium, Rhizopus).
Lignina este un polimer aromatic tridimensional, insolubil in apa, alcatuit din unitati fenil-propan, rezultat prin cuplarea aleatorie a trei tipuri de alcooli precursori:
1) Alcoolul p-cumarilic, care in polimer formeaza unitati de p-hidroxifenil;
2) Alcoolul coniferilic, care formeaza unitati de guaiacil;
3) Alcoolul sinapilic, in polimer formeaza unitati de siringil.
Copolimerizarea relativ aleatorie a acestor alcooli determina formarea unui polimer heterogen, cu o mare heterogenitate de structura si rezistenta la degradare. Unitatile de baza sunt mentinute asociat prin legaturi C-C sau aril-eter.
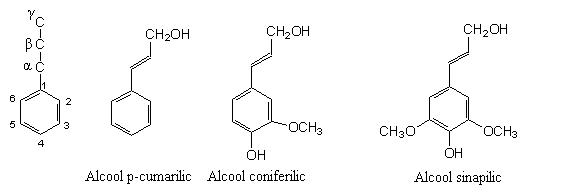
Alcoolii precursori ai ligninei si nomenclatura atomilor de carbon din structura lor.
Cei trei monomeri precursori sunt prezenti in proportii variabile in functie de specia plantei, de tipul de tesut etc.
Lignina este degradata de fungi si bacterii. Actiunea ligninolitica a bacteriilor este moderata. Principalii agenti ai degradarii ligninei in natura sunt fungii putregaiului alb. Cel mai studiat este Sporotrichum pulverulentum, care secreta enzime ce degradeaza lignina si alti constituienti ai lemnului. Eficienta deosebita a fungilor putregaiului alb, dupa Atlas si Bartha (1987), se datoreaza producerii unor agenti oxidanti ca H2O2, anionul superoxid (O2-), radicalul hidroxil(-OH), oxigenul singlet (1O2), care rup legaturile dintre subunitatile componente ale ligninei, producand depolimerizarea acesteia.
Enzima esentiala pentru degradarea ligninei este ligninaza (ligninperoxidaza), o oxireductaza, care contine hem. A fost definita ca o oxigenaza care necesita H2O2 (Tien si Kirk, 1984). In prezenta H2O2, ligninaza catalizeaza oxidarea ligninei si a compusilor inruditi, cu producere de radicali cationici.
Mn-peroxidaza, prezenta in mediul de crestere al fungilor putregaiului alb oxideaza fenolii si participa, probabil la producerea apei oxigenate.
Fenoloxidazele sunt enzime extracelulare produse de fungii putregaiului alb. Rolul lor in degradarea ligninei este putin cunoscut.
Descompunerea ligninei este un proces oxidativ, stimulat de cresterea tensiunii oxigenului si are loc numai in prezenta unei surse de C mai usor metabolizabila si in conditiile limitarii sursei de azot.
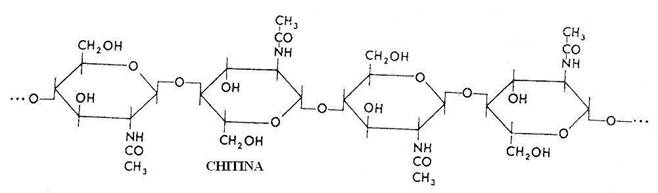
Structura chimica a chitinei.
Chitina este insolubila in apa, in solventi organici si in alcali concentrati. Poate fi solubilizata enzimatic sau cu acizi minerali concentrati. Se gaseste in peretele celular al fungilor (2,6-26% din greutatea uscata), la unele alge verzi, la nematode, anelide, moluste, celenterate, artropode. Chitina este degradata de unele bacterii de unii fungi.
Toate microorganismele chitinolitice produc o chitinaza extracelulara care ataca polimerul lung, producand separarea subunitatilor de baza de N-N-diacetilchitobioza. Acestea sunt atacate de chitobiaza si hidrolizate la glucozamina (2-amino-glucoza) si acid acetic, utilizabile ca sursa de C si energie de alte microorganisme.
Chitinoliza are o amploare deosebita in mediul marin si oceanic si este un proces semnificativ pentru ciclul biogeochimic al C.
Unele substante rezistente sau chiar "recalcitrante" la degradare sunt scoase pentru perioade mai lungi sau mai scurte din circuitele naturale. Scoaterea din circuit a unor cantitati mari de compusi ai C, organici sau anorganici, inaccesibile vietii reprezinta imperfectiuni ale ciclului.
Humusul constituie o mare parte a substantei organice din sol. El se formeaza prin conversia intermediarilor fenolici rezultati din degradarea ligninei. Humusul este foarte rezistent la degradarea microbiana, componentele sale avand o vechime cuprinsa intre 20 si 2 000 ani, stabilita prin studiile de datare cu C14. Substantele humice reprezinta o situatie intermediara in raport cu cea a combustibililor fosili.
Depozitele anorganice de C se formeaza prin precipitarea CaCO3, rezultat din combinarea acidului carbonic dizolvat in apa marii, cu Ca2+ in conditii slab alcaline.
Formaminiferele, molustele, coralii etc., formeaza "depozite biologice" de C anorganic.
Circuitul fosforului
Fosforul este un element esential pentru existenta sistemelor biologice, datorita prezentei sale in moleculele de ADN, ARN, in fosfoplipidele din structura membranelor celulare, in glicerofosfati, unele coenzime, precum si in moleculele de ADP si ATP. Fosforul este prezent in moleculele de fitina provenite din tesuturile vegetale, in compozitia oaselor, ca fosfat de Ca (85%) si ca fosfat de Mg(1,5%).
Rezervorul major de fosfor in natura este roca fosfatica apatita (3Ca3(PO4)3.CaF2), insolubil. In sol se gaseste sub forma fosfatilor hidratati (FePO4.2 H2O si AlPO4.2 H2O), ce provin din eroziunea lenta a apatitei. Fosforul din sol se gaseste in proportie de 95-99% in forme inaccesibile direct plantelor si microorganismelor. Din aceasta, 30-85% sunt compusi organici, proveniti din resturile vegetale, animale si din microorganisme (fitina, acizii nucleici, fosfolipide, glucide fosforilate).
Fosfatii anorganici sunt insolubili (fosfati de calciu, fier, aluminiu) si solubili. In fosfatii primari (NaH2PO4), numai una din cele trei valente ale PO43- este legata de un metal. Fosfatii secundari (Na2HPO4) au numai un atom de H, iar cei tertiari (Na3PO4) nici unul. Solubilitatea lor este progresiv mai mica. In mediile acvatice, concentratia fosfatilor este foarte mica, dar creste de cateva ori in celulele algelor si ale altor microorganisme. De aici sunt transferati sub forma combinatiilor organice la animale care se hranesc cu fitoplancton.
Microorganismele sunt implicate in patru mecanisme majore ale transformarii fosforului in circuitul sau in natura.
1) Mineralizarea compusilor organici ai fosforului din sol de catre microorganisme heterotrofe si regenerarea ortofosfatilor solubili. Degradarea implica participarea unui numar de enzime de tipul nucleazelor, fosfolipazelor, fosfatazelor, glicerofosfatazelor. Fitina, sarea de Ca si Mg a acidului fitic este degradata lent sub actiunea fitazei produsa de unele bacterii si microfungi. Compusii cu fosfor originari in celulele microbiene si animale sunt degradati mai rapid si este favorizat de autoliza celulelor moarte. Ortofosfatii solubili rezultati la sfarsitul procesului de mineralizare urmeaza doua cai: o parte sunt asimilati de microorganismele care i-au solubilizat; o alta parte este convertita la forme greu solubile (fosfati de Ca, Mg, Fe, Al).
2) Solubilizarea fosfatilor anorganici (Ca3(PO4)2 si a fosfatilor de Fe, Mg, Al) si eliberarea fosfatilor solubili accesibili plantelor. In sol procesul are loc cu precadere in rizosfera.
3) Imobilizarea fosforului in celulele microorganismelor. Procesul are loc si in organismele plantelor si animalelor, prin sinteza acizilor nucleici, fosfolipidelor, fitinei, glucidelor fosforilate, etc.
4) Procese de oxidare a P anorganic. Ortofosfatul este forma cea mai oxidata a P. Unele microorganisme aerobe pot utiliza fosfitii (HPO32-), convertindu-i la fosfat dupa reactia:
H3PO3 + 1/2/ O2 H3PO4
Acid fosforos Acid ortofosforic
In anaerobioza, Cl. butyricum face reactia inversa, de reducere a fosfatului la fosfit si la hipofosfit dupa reactia:
H3PO4 H3PO3 H3PO2
Acid hipofosforos
Recicularea fosforului in biosfera este numai partiala, deoarece este asociata cu pierderi continue, care sustrag cantitati importante din compusii necesari sistemelor biologice. Imperfectiunile ciclului determina fenomene de deficit de P in sol si in mediile acvatice cu consecinte negative asupra productiei primare a ecosistemelor respective.
O alta pierdere este transferul permanent, unidirectional de P din sol in mare. Cantitati importante de P din mediile acvatice se sedimenteaza in straturile profunde, devenind inutilizabile pentru viata.
Circuitul sulfului
Sulful este un component esential al biosferei, ca parte a structurii chimice a organismelor vii. Se gaseste in sol, ape si atmosfera si ocupa locul 13 in ceea ce priveste distributia cantitativa a elementelor in natura. Citoplasma contine intre 0,4-1% S sub forma unor compusi organici.
S este prezent in natura sub doua forme:
1) In forma anorganica, in depozitele de S elementar (S0) sau intr-o varietate de compusi redusi (H2S), sau oxidati ca SO2, SO3, tiosulfati (S2O32-), tetrationati (S4O62-), sulfati (SO42-) sau H2SO4.
2) In forma organica, in compozitia unor aminoacizi (cistina, cisteina, metionina), a glutationului.
Hidrogenul sulfurat (H2S) se gaseste in balti, izvoare sulfuroase, gaze naturale si in medii cu substante organice in curs de descompunere. Are origine vulcanica, hidrotermala sau biologica (produs al activitatii microorganismelor). H2S nu se acumuleaza in natura, deoarece in prezenta O2 se oxideaza rapid, spontan la SO2.
Dioxidul de sulf (SO2) rezulta din oxidarea H2S, din gazele industriale. SO2 reprezinta circa 95% din compusii S rezultati din arderea combustibililor fosili. SO2 este redus la H2S prin actiunea bacteriilor, iar in aer este oxidat la SO3 prin actiunea radiatiilor UV. SO2 poate interactiona cu O dupa reactia:
SO2 + O + M SO3 + M, in care M este o molecula de O2 sau N2 si are rolul de a prelua excesul de energie din reactie, impiedicand reversia ei.
Trioxidul de sulf (SO3) reactioneaza prompt cu vaporii de apa si formeaza acid sulfuric.
Sulfatii provin din roci sau din apa de ploaie, in care deriva probabil de la SO2 din aer. Sulfatul este extrem de stabil la reducere chimica. Cel mai adesea, reducerea sa este rezultatul actiunii directe a bacteriilor sulfat reducatoare. In anaerobioza, sulfatii sunt redusi la H2SO4 si S0.
Ciclul biologic al S in natura implica participarea a patru tipuri de reactii:
1) Mineralizarea compusilor organici ai S din componenta celulelor.
2) Oxidarea compusilor anorganici si din aminoacizi.
3) Asimilarea compusilor anorganici si incorporarea lor in materia vie.
4) Reducerea sulfatului si a S0 la sulfuri (reducerea dezasimilatorie si asimilatorie a sulfatului).
Mineralizarea compusilor organici ai sulfului
Compusii organici ai S depusi in sol, odata cu dejectiile animale si cu tesuturile vegetale si animale moarte nu pot fi utilizati ca atare de plantele superioare. Mineralizarea este rezultatul actiunii directe a unor microorganisme de putrefactie: E. coli, P. aeruginosa, Proteus sp., Clostridium sporogenes, etc. Reactia are la baza proteoliza urmata de eliberarea S din compozitia aminoacizilor cu S:
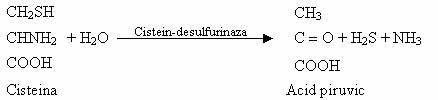
Produsii majori ai degradarii sunt, in anaerobioza H2S, iar in aerobioza H2S, mercaptanii si diferiti compusi minerali, partial sau complet oxidati (H2SO4).
Oxidarea compusilor anorganici ai S
H2S produs in natura pe diferite cai (descompunerea produsilor cu S, reducerea sulfatului, activitatea vulcanica, etc) este numai intr-o mica masura imobilizat ca sulfuri instabile sau oxidat spontan la S0 in prezenta O2. Cea mai mare parte a H2S, ca si a S0 sunt oxidate pe cale biologica.
Cele mai studiate sunt bacteriile din g. Thiobacillus cu speciile: T. thiooxidans, T. thioparus, T. ferooxidans, etc, precum si din g. Thiomicrospira si Sulfolobus. Traiesc in medii bogate [n S0 si H2S (lacuri, balti, mari, apa de canal, izvoare termale acide). Oxideaza S0 si diferiti compusi anorganici ai acestuia, dupa reactiile:
Th. thiooxidans: 2 S0 + 3 O2 + 2 H2O 2 H2SO4
Th. thioparus: H2S + 2O2 H2SO4
Na2S2O3 + 2O2 + H2O Na2SO4 + H2O
2Na2S4O6 + 7O2 + 6H2O 2Na2SO4 + 6H2SO4
Oxidarea So la sulfat este insotita de o producere masiva de H+. Mediul se acidifica pana la pH 1,0.
Th. ferooxidans este in acelasi timp sulf- si ferooxidanta, deoarece oxideaza So, H2S, S2O62-(tetrationat), SO32-(sulfit) la H2SO4 si respectiv la sulfati, dar si Fe3+:
2FeS2 + 2H2O + 7O2 2FeSO4 + 2H2SO4
4 FeSO4 + O2 + 2H2SO4 2Fe2(SO4)3 + 2H2O
Al doilea grup de bacterii sulfoxidante este reprezentat de bacteriile din genurile: Beggiatoa Thiotrix, Thioploca, Thiobacterium, Thiospira etc. Cele mai multe nu au fost izolate in culturi pure si se gasesc in medii care contin H2S si O2. Reactia majora este:
2H2S + O2 2So + 2 H2O
Sulful elementar este depus sub forma unor granulatii refrigente. Reactia este reversibila: dupa epuizarea H2S din mediu, So din incluziuni este oxidat la H2SO4.
Al treilea grup de bacterii sulfooxidante este reprezentat de bacteriile fotosintetizante sulfuroase. Ele apartin la doua categorii:
a) Bacteriile sulfuroase purpurii (Thiorhodaceae), care se deosebesc in functie de modul in care depun incluziile de So: Ectothiorhodospira depune So extracelular, iar Thiospirilium, Thiocystis, Thiocapsa, Chromatium, Lamprocystis si Rhodothecae depun So intracelular.
b) Bacteriile sulfuroase verzi - Chlorobiaceae (Chlorobium, Chloropseudomonas, Pelodictyon) utilizeaza H2S ca donor de e- pentru reducerea CO2. So format ca intermediar in oxidarea sulfurilor la sulfat este depus extracelular.
Bacteriile fototrofe purpurii si verzi utilizeaza compusi ai S (H2S, S2O32-, SO32-), iar unele chiar compusi organici. Energia stocata ca rezultat al fotosintezei este folosita pentru sinteza constituientilor celulari de la CO2, dupa reactia:
H2S + 2 CO2 + 2H2O H2SO4. + 2(CH2O)
Reducerea asimilatorie a sulfatilor este efectuata in exclusivitate de bacterii, de unele levuri si de plante, dar niciodata de organismele animale. Produsul final al reducerii asimilatorii a sulfatului este cisteina, de la a carei grupare -SH deriva ceilalti aminoacizi cu sulf, precum si biotina, tiamina, acidul pantotenic, glutationul.
Reducerea dezasimilatorie a sulfatilor semnifica utilizarea lor ca acceptor final de e- in procese de dezasimilare a unor substraturi organice in respiratia anaeroba. Procesul s-a denumit respiratia sulfatului. Numai o mica parte din sulfatul redus este asimilat de organism, cea mai mare parte fiind eliberat in mediul extern ca S2- sau ca H2S liber.
Hidrogenul necesar pentru reactie poate fi furnizat de compusi organici ca lactatul, dupa reactia:
2C3H5O3Na + MgSO4 H2S + 2C3H3O2Na + MgCO3 + CO2 + H2O
sau de H2, pentru reducatorii de sulfat autotrofi, dupa reactiile:
4 H2 + SO42- S2- + 4 H2O
3 H2 + SO32- S2- + 3 H2O
4 H2 + S2O32- S2- + H2S + H2O
9 H2 + S4O62- S2- +3 H2S +6 H2O
4 H2 + CaSO4 H2S + Ca(OH)2 + 2 H2O
Bacteriile sulfat reducatoare reprezinta principalul producator de H2S in natura.
Depozitele de sulf au cel mai adesea o origine biogena si s-au fornmat prin reducerea sulfatilor, urmata de o etapa de oxidare, la care au participat bacteriile sulfoxidante. Dovada o constituie existenta sulfuretelor in care se acumuleaza si in prezent.
Bacteriile sulfat-reducatoare ce se dezvolta in apele de canal, in apele stagnante si in apele din porturile intens poluate, determina mirosul neplacut datorita producerii intense de H2S. Reducerea masiva a sulfatilor in natura este rezultatul activitatii bacteriene sulfat-reducatoare.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5740
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved