| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
STRUCTURA CROMOZOMULUI
Plicaturarea ADN-ului in cromozomi si cromatina
Fiecare cromozom contine o singura molecula de ADN organizata intr-o succesiune de secvente plicaturate care construiesc un cromozom metafazic.
Lungimea cromozomului metafizic este 1/10000 din lungimea ADN-ului sau
Nivele de impachetare si organizare ale cromatinei
a. ADN-ul in cromozomi este asociat cu proteine de legatura. Complexul ADN-proteina se organizeaza in cromatina . Aceste proteine pot media transcriptia genica si replicarea, sau pot fi proteine structurale care intra in structura AND.
b. Histonele sunt proteinele structurale ale cromatinei si sunt cele mai abundente proteine din nucleu: cinci histone (H1, H2A, H2B, H3 si H4) furnizeaza cadrul pentru unitatea fundamentala de impachetare a cromatinei = nucleozomul
c. Fiecare nucleozom este format dintr-un cilindru strans de 8 histone (de doua ori H2A, H2B, H3, H4) infasurat de 2 ori in jurul suprafetei de helixul ADN
-miezul fiecarui nucleozom cuprinde 146 de perechi de baze ale helixului ADN si miezurile sunt legate de aproximativ 60 baze intr-o structura ca un sirag de margele
-largimea pachetului nucleosomal este de 11 nanometri (nm) comparativ cu 2 nm pentru dublul helix de ADN
-nucleozomii sunt plicaturati strans impreuna cu ajutorul moleculei histonice H1; se formeaza o fibra cu grosimea de 30 nm, care este unitatea cromatinica fundamentala, vizibila in nucleu in interfaza, cu microscopul electronic.
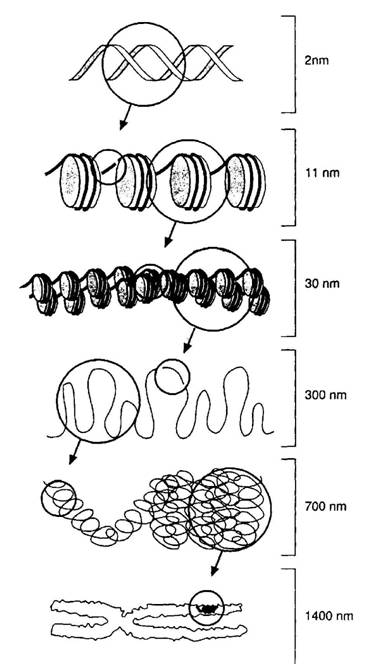
Fig. 7. Structura cromozomului
d. Fibra de 30 nm este ulterior impachetata intr-un sistem al domeniilor buclate
Inelele care contin de la 20.000 - 100.000 perechi de baze sunt formate de proteine nonhistonice legand situsuri specifice de-a lungul fibrei de 30 nm
Dovezile sugereaza ca inelele adapostesc unitati individuale de transcriptie si replicare ale AND-ului, de altfel, ele au fiecare o semnificatie structurala si functionala.
Organizarea superioara a ADN-ului
In metafaza cromatina este condensata mai puternic comparativ cu cromatina interfazica
Procesele condensarii din profaza mitozei sunt urmarea incolacirii si ingramadirii domeniilor buclate; in stare condensata inelele alaturate sunt tinute impreuna de interactiunea proteinelor care formeaza o plasa
Reteaua cromatinica este aranjata intr-o maniera helicoidala de-a lungul axului cromozomului
Grosimea (largimea) unei cromatide individuale in metafaza cromozomului este de aproximativ 700 nm, reflectand un grad crescut de comprimare a fibrei cromatinice de 30 nm
Benzile cromozomiale. Profaza si metafaza cromozomilor expune benzi alternative clare si intunecate in conditii specifice de colorare . Aceste benzi reflecta plicaturarea diferita a clusterilor domeniilor buclate si deasemenea definesc segmente ale genomului care au diferite proprietati si functiuni
Caracteristicile speciale ale cromozomilor
v Eucromatina formeaza corpul principal al cromozomului si are o densitate relativ crescuta a segmentelor codante a genelor.
a. Benzile R sunt domenii care :
-Se vizualizeaza ca pete luminoase in procedurile de bandare pentru benzile G
-ADN-ul replica devreme in faza S a ciclului celular
-au un continut relativ crescut in G si C
-au majoritatea SINES
-au cea mai mare densitate genica
b. Benzile G sunt domenii care:
-raman intunecate in proceduri de bandare pentru benzile G
-au replicare tarzie in faza S
-au un continut mai mare in A si T decat benzile R
-au majoritatea LINES
-au relativ mai putine gene decat benzile R
v Heterocromatina este cromatina care este lipsita de gene sau care are gene inactive. Segmentele heterocromatinice ale genomului raman mai condensate in interfaza decat eucromatina si se replica foarte tarziu in faza S.
a. Heterocromatina constitutiva este localizata in jurul centromerului tuturor cromozomilor, pe bratul lung al cromozomului Y, si in satelitii cromozomilor acrocentrici . Aceste domenii contin familii variate de elemente inalt repetitive de ADN cu functie necunoscuta. Poate fi puternic luminata in metafaza folosind procedurile de bandare pentru benzi C sau Q .
b. Heterocromatina facultativa este eucromatina intr-o stare inactiva transcriptional. Un exemplu elocvent este lyonizarea: unul din cei 2 cromozomi X ai femeii este inactivat in scopul compensarii dozei duble a genelor de pe cromozomii XX comparativ cu o singura doza pe perechea XY a barbatului
Mecanismul inactivarii nu este cunoscut; dar unul dintre rezultatele procesului este modificarea metilarii citozinei in secventele din apropierea genelor cu transcriptie regulata. Inactivarea unui X este de asemenea asociata de schimbarea momentului replicarii ADN-ului: mai tardiva in faza S. Cromozomul X inactivat ramane puternic condensat si se coloreaza intens in timpul interfazei formand corpusculul Barr.
v Centromerul apare ca un domeniu de constrictie a cromozomului metafazic. In timpul diviziunii celulelor centromerul este ultimul segment al cromozomului care se replica si separa
a. Structura: consta in secvente specifice de ADN care leaga proteine care pot fi identificate prin proprietatile lor antigenice
b. Functia: proteinele centromerice formeaza kinetocorul, care impreuna cu microtubulii fibrelor fusului de diviziune separa cromatidele surori si cromozomii in anafaza mitozei sau meiozei
v Telomerele : sunt secvente de ADN care se gasesc la capetele cromozomilor si care au rolul de a mentine stabilitatea cromozomilor. Cromozomul fara telomere are tendinta la recombinare cu alte segmente cromatinice si poate suferi rupturi si deletii.
a. Structura: segmentele terminale ale tuturor cromozomilor au o secventa ADN similara (TTAGGG) care este repetata in cateva mii de copii
b. Functia: structura intacta a telomerelor faciliteaza replicarea ADN-ului la capetele cromozomului.
Analiza cromozomilor si clasificarea lor
Studii citogenetice: analiza si diagnosticul citogenetic poate fi performant in metafaza sau prometafaza cromozomilor obtinuta prin diviziuni rapide celulare in culturi de celule sau in unele cazuri direct din tesuturi cu activitate mitotica crescuta (celule tumorale)
a. Sursa celulelor
Cromozomii din culturi limfocitare constituie cea mai rapida si convenabila metoda de rutina pentru analiza citogenetica. Aceste culturi pot folosi limfocite periferice care permit prepararea cromozomilor in metafaza in 3 zile.
Culturi celulare din alte tipuri de celule (fibroblaste, amniocite, celule ale vilozitatilor coriale, celule tumorale) necesita 1 - 2 saptamani pentru a cumula suficiente celule pentru analiza citogenetica. Fibroblastii si alte culturi de celule pot fi subcultivate si pastrate saptamani sau luni. De asemenea pot fi inghetate in azot lichid si pastrate ani pentru viitoarele analize.
Prepararea directa a celulelor pentru analiza cromo-zomilor poate fi facuta din celulele maduvei osoase a pacientilor cu leucemie sau pe tumori solide. Aceste tehnici au avantajul unei rate crescute a mitozei in aceste tesuturi.
b. Metode pentru analiza cromozomilor
Bandarea G este cea mai comuna metoda care foloseste coloratia Giemsa dupa tratarea enzimatica a cromozomilor metafazici pentru a diferentia benzile cromozomiale - vizualizati sub lumina microscopului cromozomii metafazici au benzi alternative luminoase si intunecate. Benzile intunecate, conventi-onal sunt numite benzi G, benzile luminoase sunt benzi R.
- In medie, in preparatele metafazice conventionale, aproximativ 400 de benzi intunecate si clare pot fi observate intr-un set haploid de cromozomi
Bandarea Q - quinacrina coloreaza cromozomii care sunt vizibili la microscopul cu fluorescenta. Alternarea benzilor luminoase si estompate fluorescente corespund celor vizibile si in bandarea G pentru cele mai multe clone vii ale cromozomilor. Benzile Q stralucitoare sunt echivalente benzilor G intunecoase. Domeniile lipsite de aceasta similitudine sunt numite benzi variabile
Bandarea R sau bandarea reversibila foloseste coloratia Giemsa la temperaturi ridicate sa vizualizeze in reversie benzile G sau Q. Coloratia fluorescenta cu o afinitate crescuta a ADN-ului bogat in GC (olivomicin) poate produce de asemenea benzi R
Benzile C apar la incalzire intr-o solutie alcalina si coloratie cu Giemsa; benzile C sunt regiuni de heterocromatina constitutiva localizate adiacent centromerilor tuturor cromo-zomilor si in bratul lung al cr. Y
Metoda de bandare cu rezolutie inalta prinde cromozomii in profaza sau prometafaza, cand ei sunt mai putin evidentiabili decat in metafaza. Metodele implica sincronizarea partiala a mitozei in cultura prin oprirea sintezei ADN, ceea ce face ca celulele sa se acumuleze in acest moment din ciclul celular. Deci mai multe celule pornesc in mitoza impreuna. In aceasta metoda, fiecare banda vizibila in metafaza cromozomilor poate fi impartita in subbenzi (800 sau mai multe). Analizele profazei cromozomiale cu rezolutie inalta (850 / 9000 benzi) nu este o tehnica de practica generala
Analiza situsului fragil poate fi realizata crescand cultura de celule in conditii specifice pentru inducerea in fiecare a situsului fragil. Ex: detectarea situsului fragil X sensibil la folat, care e asociat cu retardarea mentala X- lincata; celulele trebuie sa cresca in mediu fara acid folic sau tratate cu methotrexat sau (FdUr) fluordezoxiuridina, care interfereaza cu metabolismul folatului.
Hibridizarea in situ a probei de ADN din celulele metafazice si profazice
(a) Aceste metode implica hibridizarea in situ a probelor specifice de ADN etichetate in metafaza cromozomilor sau de cromatina interfazica.
(b) Probele de ADN sunt etichetate cu molecule "reporter" cum e biotina, digoxigenina si dimetrophenylul, care pot fi vizualizate sub microscop cupland moleculele "reporter" cu un semn fluorescent sau alt colorant. Agentii fluorescenti sunt cei mai folositi dand termenul de hibridizare fluorescenta in situ (FISH)
(c) Metoda FISH este foarte importanta . Poate fi utilizata in determinarea originii cromozomilor markeri mici care nu pot fi complet caracterizati prin bandare
(d) Cea mai comuna folosire a metodei FISH este pentru detectarea deletiilor submicroscopice cum ar fi cele asociate cu sindrom Prader/Willi.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 5374
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved