| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Vertebrate
Studiul vertebratelor fosile reprezinta un domeniu interdisciplinar care implica cunostinte temeinice de zoologie, fiziologie, anatomie comparata, embriologie, teorii evolutioniste, stratigrafie si geologie.
Istoria
evolutiva a vertebratelor a inceput in urma cu aproximativ
Evolutia vertebratelor poate fi interpretata din cel putin doua puncte de vedere:
- in sens istoric - prin urmarirea modificarilor de structura si adaptare in timp;
- din punct de vedere al modificarilor genetice, al proceselor de selectie naturala si de dinamica a populatiilor care explica cum apar aceste schimbari in timp.
Moduri de fosilizare ale vertebratelor - Procesele de fosilizare limiteaza foarte mult informatiile privitoare la organismele vertebrate din cadrul unor grupe extincte. Dupa moartea organismelor, tesuturile moi se descompun, putrezesc, dar oasele pot fi acoperite de sedimente in care se infiltreaza solutii de apa incarcate cu minerale solubile si sedimente ce umplu cavitatile mai largi ale oaselor si precipita in micile canale ocupate in timpul vietii de celule si vase de sange. Corpul moale poate fi mancat, sfasiat de pradatori, iar oasele pot fi sparte, dezarticulate. Probabil un numar extrem de mic de indivizi (numai un individ dintr-un milion, dupa unele opinii) a fost rapid acoperit de sedimente astfel incat a fost posibila fosilizarea completa a elementele scheletice. La majoritatea vertebratelor fosile, mineralele componente ale oaselor isi pastreaza in proportie destul de mare integritatea, astefl incat detaliile histologice si chiar compozitia chimica sunt relativ putin alterate chiar si dupa sute de milioane de ani.
Alte moduri de fosilizare: prin deshidratare rapida (mumifiere); in curgerile de lava; in chihlimbar; resturi carbonificate sub forma de impresiuni care pastreaza forma generala a corpului (in sedimente cu granulatie fina acumulate intr-un mediu anaerobic); prin metasomatoza (piritizare, silicifiere); urme la activitatii vitale: urme de pasi, coprolite, etc.
Scheletul vertebratelor - reprezinta partile tari, mineralizate ale corpului care au rol de a sustine corpul si care servesc ca locuri de insertie pentru muschi si ca elemente de protectie a organelor interne.
In functie de originea si pozitia sa scheletul poate fi: exoschelet (schelet extern) si endoschelet (schelet intern).
Exoscheletul este format din tesut osos care ia nastere din stratele superficiale ale dermei, fara a trece prin stadiul intermediar cartilaginos. Exemple de vertebrate cu structuri exoscheletice bine dezvoltate: agnathe, pestii placodermi, placodonti (reptile mesozoice). Solzii ososi reprezinta cele mai frecvent intalnite structuri exoscheletice la pesti, la unele agnathe fosile si la unii dintre amfibienii fosili (stegocephali).
Endoscheletul este generat de celulele osteogene ale mezodermului. Endoscheletul este alcatuit din doua tipuri de oase: oase dermice (de membrana, de acoperire) care iau nastere la fel ca si exoscheltul din tesuturile superficiale ale dermei si oase endochondrale (de cartilaj, de inlocuire) care se formeaza prin inlocuirea tesutului cartilaginos cu tesut osos (deci trec prin stadiul intermediar de cartilaj).
In general vertebratele prezinta diferite tipuri de tesuturi calcifiate cum ar fi dentina, emailul, diferite tipuri de oase, etc. Oasele endochondrale sunt insa unice deoarece ele isi incep dezvoltarea din cartilagii. La majoritatea vertebratelor bazale structurile cartilaginoase pot deveni superficial calcifiate in cel putin doua moduri diferite. De exemplu la Chondrichtyes (pesti cartilaginosi) se dezvolta placute osoase microscopice care acopera suprafata cartilajului generand astfel un cartilagiu calcifiat prismatic. Toate gnatostomatele (vertebrate cu maxilare) si probabil unele grupe bazale de vertebrate, pot produce de asemenea si straturi succesive de tesut osos care acopera suprafata cartilagiului, generand astfel un tesut osos dens, lamelar, denumit os perichondral.
Oasele endochondrale sunt generate de actiunea celulelor osteoblaste. La Osteichtthyes (pesti ososi) sistemul circulator invadeaza practic matricea cartilaginoasa. Acest fapt permite osteoblastelor sa continue formarea osului in interiorul cartilagiului, recrutand aditional si alte osteoblaste. Aceste celulele practic inlocuiesc gradat cartilagiul inconjurator din interior catre exterior, astfel incat, treptat ia nastere un tesut osos vascularizat, neregulat, adica un os "spongios" in interior, inconjurat la exterior de un tesut osos dens, lamelar, periostial, rezultand in final un os endochondral.
Voi prezenta in continuare o descriere succinta ("minimum minimorum") a principalelor grupe de chordate fosile si actuale. Ordinea in care voi prezenta principalele grupe de chordate este data (in general) in concordanta cu sistematica cladistica care demonstreaza pe baza synapomorozelor, monofilia diferitelor grupe de chordate (clade). Pentru fiecare grup (clad) vor fi evidentiate principalele caractere diagnostice, synapomorfoze, distributia stratigrafica, exemple de genuri cu semnificatie filogenetica si/sau biostratigrafica(*)
CHORDATA
------ UROCHORDATA
------ CEPHALOCHORDATA
------ (?) CALCICHORDATA
------ CRANIATA:
-------- ----- ------ ------------ HYPEROTRETI (Myxini)
-------- HYPEROATRIA (Petromyzonidae)
----------VERTEBRATA -------- ? EUCONODONTA
-------- "OSTRACODERMI" (Agnathe fosile)
-------- GNATHOSTOMATA (Placodermi, Chondrichtyes, Achanthodii, Actinopterigii, Sarcopterigii, Tetrapodomorphe, Tetrapoda, Reptiliamorpha, AMNIOTA (chelonieni, soparle, dinosauri, pasari, mamifere).
Chordatele - sunt animale coelomate, deuterostomieni, cu simetrie bilaterala, cu organizare anatomica superioara. Majoritatea sunt organisme mobile, inzestrate cu organe adaptate pentru inot si / sau deplasare terestra.
Caractere diagnostice ale chordatelor (adica, caractere care nu se regasesc la celalte grupe de animale studiate anterior):
- faringotremia - existenta unor perforatii in peretele partii anterioare a tubului digestiv (in faringe) = dispozitivul respirator al fantelor branchiale;
- notochordul - un schelet intern sub forma unei baghete rezistente, elastice, dispusa sagital, in tot lungul corpului, situat dorsal, deasupra tubului digestiv; la vertebratele evoluate notochordul va fi inlocuit cu coloana vertebrala. Notochordul consta dintr-un sir de celule, fiecare continand o vacuola centrala larga plina cu lichid limfatic. Celulele sunt inconjurate de o teaca de tesut conjunctiv dens care le mentine turgescenta (fenomenul de "umflare" a celulelor cu lichid limfatic). Functia primara a notochordului este aceea de a rezista la compresie, prevenind astfel scurtarea corpului. Notochordul apare la embrionii tuturor chordatelor dar in stadiile adulte notochordul dispare (la Urochordate) sau este inlocuit (inconjurat de corpurile vertebrale) partial sau total la Vertebrate.
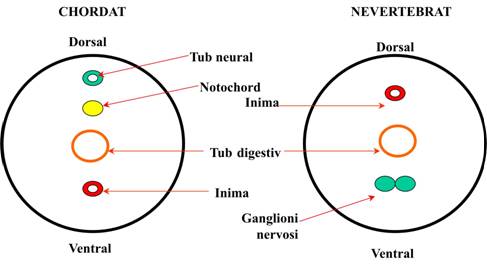
- epineurie - pozitia dorsala a tubului neural, deasupra tubului digestiv si a notochordului - caracteristica distinctiva fata de nevertebrate la care cea mai mare parte a sistemului nervos (cordoane nervoase, ganglioni nervosi scalariformi) sunt situati ventral, sub tubul digestiv (Fig. 1). La vertebrate capatul anterior al tubului neural se dilata si formeaza creierul.
- prezenta unei "cozi" (sau inotatoare codala) in pozitie post-anala sau o prelungire (mai mult sau mai putin dezvoltata) a notochordului si a tubului neural posterior fata de anus. La tunicate si cephalochordate "prelungirea codala post-anala este prezenta numai in stadiile larvare. La pesti eficienta cozii este crescuta prin aparitia radiilor cartilaginoase sau osoase. La alte vertebrate (broaste, oameni) prelungirea codala dispare.
- musculatura segmentata (myomere).
|
|
|
Figura 1. Diferentele dintre planul structural general de organizare al chordatelor si nevertebratelor. |
Planul structural general de organizare al chordatelor pastreaza insa o serie de caractere comune cu cele ale nevertebratelor superioare. De exemplu, simetria bilaterala, axul antero-posterior, coelomul, metamerizarea si cephalizarea. Totusi diferentele dintre nevertebrate si vertebratele primitive (inferior organizate) (Fig. 1) sunt destul de clare si nu este inca posibil sa se stabileasca cu exactitate relatiile de inrudire dintre nevertebrate si chordate.
(*) Ipoteza dupa care echinodermele ar putea furniza informatii asupra stramosului chordatelor este in acest moment cea mai acceptata si se bazeaza pe o serie de caractere pe care cele doua grupe le au in comun: clivajul radial al oului, anusul derivat din blastoporul gastrulei, iar orificiul bucal reprezinta o neoformatie (adica sunt deuterostomieni) si coelomul este rezultat prin enterocoelie (cu exceptia unor vertebrate schizocoelice).
(*) Alte caracteristici ale chordatelor:
- corpul chordatelor este segmentat (metamerizat) prezentand cavitati coelomice perechi (dreapta si stanga) ca si in cazul annelidelor si arthropodelor, dar, spre deosebire de acestea din urma somitele chordatelor sunr evidente numai pe partea dorsala a corpului, pe partea ventrala fiind fuzionate.
- sistemul circulator la chordate este reprezentat printr-un sistem inchis de vase prin care curge sangele. Circulatia sanguina este craniano-caudala prin vasele dorsale si caudalo-craniana prin vasele ventrale, deci un circuit invers fata de nevertebrate. Sangele chordatelor contine oxigen care este transportat de moleculele de hemoglobina.
- fantele faringeale au suferit importante modificari pe parcursul evolutiei chordatelor. La chordatele primitive aceste fante erau utilizate pentru filtrarea particulelor de hrana din apa. La pesti si o parte dintre amfibieni fantele faringeale poarta branchii fiind utilizate pentru schimbul de gaze. Aceste fante faringeale branchiale au luat nastere prin invaginarea ectodermului si evaginarea endodermului faringian. La majoritatea chordatelor actuale, terestre (vertebrate superioare) aceste fante branchiale faringeale sunt prezente numai in stadiile embrionare sau ele nu se mai formeaza, insa locul unde acestea ar fi trebuit sa se dezvolte este marcat de existenta unor adancituri prezente si la embrionii umani. La aceste vertebrate apar plamanii formati din pungi ventrale ale faringelui embrionar;
"AGNATHA" (vertebrate fara maxilare)
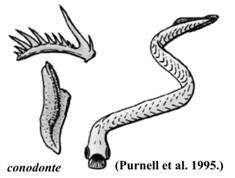
CONODONTA (EUCONODONTA) (Cb2
- T3) (540 -
Sunt reprezentate de obicei in stare fosila prin piese
(denticuli) de talie microscopica "conodonte" alcatuiti in principal din fosfat
de calciu. Primul animal conodont articulat, complet, a fost gasit in
|
|
|
Figura 2. Diferite tipuri de conodonte (stanga) si reconstituirea unui euconodont (dreapta). |
|
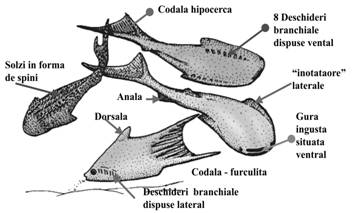
Figura 4. Caractere morfologice generale ale thelodontilor (dupa Janvier, 1996) |
THELODONTI (S - D) - Exoschelet dermic alcatuit din numerosi solzi de dimensiuni reduse, rombici, neimbricati sau in forma de spini alungiti (Fig. 4). Majoritatea aveau corpul aplatizat dorso-ventral cu 8 deschideri branchiale situate pe partea ventrala, cu o deschidere inhalanta mediana larga si o gura mica, ingusta, situata ventral. Alti thelodonti aveau corpul aplatizat lateral, deschiderile branchiale dispuse lateral si o codala mare in forma de furculita. Traiau in medii marine, cu ape putin adanci, din preajma Gondwanei si a Laurasiei. |
|
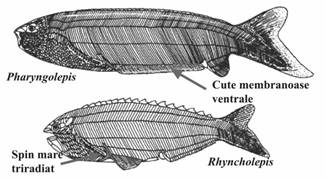
ANASPIDA (S - ?D3) (430-410m.a.) - corp fusiform, comprimat lateral; - 8 - 15 - 20 deschideri branchiale dispuse in diagonala in spatele ochilor; - intre orbite existau 2 orificii: pineal; nazohipofizar; - exoschelet dermic: placi osoase mari, groase, dorsale si dorsolaterale care acopereau numai regiunea capului; - restul corpului esa acoperit numai de solzi mici; - placile si solzii erau alcatuiti din aspidina (tesut osos laminar, acelular, nemineralizat); - inotatoare anala redusa, codala hipocerca; - endoscheletul era probabil cartilaginos; traiau in medii marine costale; Exemple: Birkenia, Jamoytius, Pharyngolepis, Rhyncholepis, Silurian (Fig. 5). |
Figura 5. Caractere morfologice generale ale anaspidelor (dupa Janvier, 1996) |
Gnathostomatele sunt reprezentate prin doua clade exclusiv fosile: Placodermi (S1 - D3) si Acanthodii (O3 - P1) si prin toate celelalte vertebrate care includ: rechinii, pisicile de mare, chimerele, pestii ososi, coelacanthia, si toate vertebratele terestre.
(*) Gnasthomatele cuprind urmatoarele clade majore:
GNATHOSTOMATA
|----- ----- --------- ----- ------- PLACODERMI
`----- ----- --------- ----- ----- CHONDRICHTHYES
`TELEOSTOMI
|----- ----- ----------- ACANTHODII
`OSTEICHTHYES
|----- ----- ------ ACTINOPTERYGII
`----- ----- ------ SARCOPTERYGII
........
...........
PLACODERMI (S -D, "pesti cuirasati") - placodermii sunt considerati ca reprezentand cea mai veche ramura a gnathostomelor sau forme primitive (bazale), aberante ale gnathostomelor. Desi prezinta o larga variabilitate a structurilor interne si externe, ei au totusi cateva trasaturi comune:
- prezentau un invelis exoscheletic format din placi osoase dermice care acopereau complet partea anterioara a corpului (adica acopereau capul si o portiune mai mult sau mai putin extinsa din trunchi) formand un fel de carcasa alcatuita din doua portiuni: un scut cephalic si un scut toracic, articulate printr-un sistem tubercul si foseta; acest sistem era situat pe partile laterale, la limita dintre cele doua scuturi, asigurand o oarecare mobilitate intre cele doua parti; zona de jonctiune dintre scuturi corespundea deschiderii branchiale;
- ochii erau mari, inconjurati de un inel sclerotical format din 1-4 piese, iar globii oculari erau conectati la cutia craniana printr-un mic cartilagiu (la fel ca la rechini);
- inotatoarele pectorale protejate sau complet inchise de spini ososi simpli sau articulati; codala heterocerca; prezentau 1-2 inotatoare dorsale;
- endoschelet relativ bine osificat - numai arcurile neurale si hemale erau osificate, centrumurile nu erau inca osificate; restul scheletului intern era cartilaginos sau alcatuit din tesut osos perichondral;
Desi capul si o parte a trunchiului erau protejate de o armura masiva de oase dermice, restul corpului era destul de vulanerabil, fiind acoperit numai de solzi mici sau era nud. Nu se cunoaste exact care era scopul acestei armuri din apartea anterioara a corpului, dar s-a sugerat faptul ca aceasta ar fi putut constitui un mijloc eficient de protectie impotriva scorpionilor de mare giganti care traiau in apele salmastre in acelsi timp cu placodermii.
La inceputul aparitiei lor in Silurianul inferior placodermii nu erau foarte raspanditi. Incepand insa cu Devonianul inferior acestia au dezvoltat o radiatie evolutiva exploziva, devenind dominanti in special in ecosistemele salmastre sau in cele marine din apropierea tarmului si chiar in apele dulci si in domeniul oceanic dechis. Placodermii au atins cea mai mare diversitate in Devonian (perioada denumita si "Age of Fishes"). Placodermii cuprind atat primele vertebrate care au colonizat apele dulci cat si cele mai vechi vertebrate care colonizat oceanul deschis. In Devonian alaturi de placodermi, traiau de asemenea ostracodermii, primii rechini (Chondrichtyes), primii pesti ososi si primii sarcopterigieni. Multi dintre acesti taxoni au disparut insa la sfarsitul Devonianului, din motive care inca nu sunt bine cunoscute sau intelese. Placodermii au supravietuit de asemena pana la sfarsitul Devonianului, iar exinctia acestora pare sa fi intervenit brusc.
|
Figura 7. Dunkleosteus, unul dintre cei mai mari placodermi, L = aprox. 9m; L partii anterioare conservate - 3,2m Devonian) (J. Conrad, 1999) |
|
|
|
Figura 8. Bothriolepis Devonian sup., L aprox. 20 to 30 cm. |
ACANTHODII (S1 - P1) ("pestii cu spini") - reprezinta cel mai vechi grup de vertebrate gnathostome cunoscut din Silurian prin solzi si spini ososi ("ihtiodoruluite"). Corpul era fusiform (L - 20cm), comprimat lateral, codala heterocerca, acoperit de solzi mici, rombici, grosi (de tip cosmoid - tesut vacuolar, vascularizat); orbitele largi, inconjurate de placi circumorbitale; deschiderile branchiale acoperite de cute membranoase (rasfrangeri ale pielii). Denumirea grupului provine de la faptul ca toate inotatoarele perechi si neperechi (cu exceptia codalei) erau constituite din cute dermice fara schelet intern, in schimb toate erau sustinute de spini ososi robusti, uneori foarte alungiti. La unele genuri spinii erau mobili, la altele acestia erau fixati. Cei primitivi (de exemplu Climatius - Fig. 10.) aveau un numar mare (4-5) de inotatoare suplimentare intermediare (dispuse intre pectorale si pelviene).
|
Figura 10. Climatius (S3 - D1) |
OSTEICHTHYES ("pestii ososi") (C1 - Q) - pesti la care endoscheletul cartilaginos din stadiul embrionar este inlocuit total sau partial in stadiul adult, cu tesut osos. Aceasta evolutie ontogenetica nu este insa invariabil reprodusa in cadrul evolutiei filogenetice a pestilor ososi, deoarece exista grupe (dipnoi, sturionii) la care s-a produs o regresiune a tesutului osos de la formele primitive la cele evoluate, prin pastrarea tesutului cartilaginos embrionar si la indivizii adulti datorata fenomenului de neotenie (capacitatea de reproducere inca din stadiul larvar). Osteichthyes reprezinta primul grup de vertebrate cu oase endochondrale adevarate. Osteichthyes cuprinde doua clade majore:
- ACTINOPTERYGII ( ? P2, T - Q) - cu inotatoarele sustinute pe intreaga lor suprafata de radii osoase (lepidotrichii); Ex. Din grupul Teleostei : Palaeorhynchus humorensis, Oligocen, Romania (vezi explicatia mai jos).
- SARCOPTERYGII (D - Q) - inotatoarele prezinta o parte carnoasa dezvoltata.
SARCOPTERYGII: partea distala a inotatoarelor era moale, carnoasa ; Actinistia (Coelacanthia) (Dmed. -Q) si Rhipidistia (Dinf. - Q) ; Caractere generale: inotatoarele au partea distala carnoasa (gr. : sarcos - moale, pterygium - aripa); prezentau doua inotatoare dorsale; codala heterocerca la formele primitive, cu tendinta spre tipul dificerc la cele evoluate; solzi rombici de tip cosmoid - cu trei strate de tesut osos laminar, dintre care stratul de cosmina este distinctiv. O parte dintre sarcopterygieni traiau in ape dulci, continentale, supuse periodic secarii. Astfel la pestii dipnoi (cu dubla respiratie) din cladul Rhipidistia se observa restrangera la maximum a functiilor vitale. Grupul in general prezinta o adaptare progresiva la viata terestra, prin restructurarea dispozitivelor locomotorii, care a condus la aparitia tetrapodelor terestre.
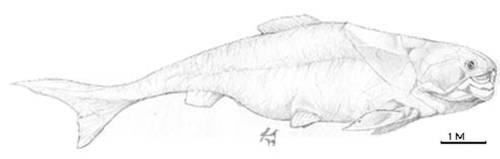
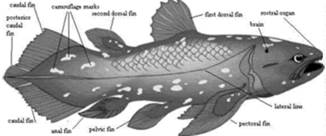
Actinistia (Coelacanthia) (Devonian mediu -Actual) - In timpul Paleozoicului traiau in ape marine si dulci. In Mesozoic si Neozpic traiau numai in ape marine. Genul Latimeria (Fig. 13) este singurul reprezentant actual (a fost descoperit pentru prima data in anul 1938, in apele din preajma insulei Madagascar). Caractere generale: plamanii acestora si-au pierdut functia respiratorie, devenind calcifiati; bolta craniana formata din oase subtiri separate prin largi spatii cartilaginoase; dinti numai in partea anterioara a maxilarelor; notochordul persistent; codala dificerca.
|
Figura 13. Latimeria (Coelacanthia) - Actual |
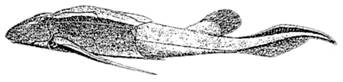
Figura 14. Diplurus newarki, un coelacanth primitiv (Triasic) |
Rhipidistia (Devonian inferior - Actual) - cuprinde doua mari grupe:
- Dipnomorpha(D inf. - Q): Porolepiformes (D), Dipnoi (D - Q)
- Osteolepiformes (Dmed. - P inf.) - au dat nastere vertebratelor tetrapode.
Dintre acestera retineti grupele Dipnoi si Osteolepiformis.
|
Dipnoi (D - Q) - reprezentantii din Devonian traiau in ape dulci din Devonianul continental din Europa, nordul Africii, America de Nord si prezentau urmatoarele caractere generale: solzi grosi de tip cosmoid; endoschelet bine osificat; codala heterocerca (Fig. 15); reprezentantii din intervalul T - Q, se caracterizeaza prin: reducerea grosimii solzilor; reducerea tesutului osos in cadrul craniului si al coloanei vertebrale, tesutul cartilaginos embrionar fiind pastrat si la adult; o flexibilitate sporita a corpului, necesara perioadelor de seceta. |
Figura 15. Scumenacia curta, un sarcopterygian dipnoi primitiv (D sup., Quebec) |
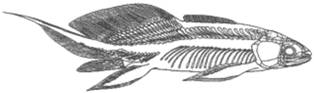
Osteolepiformes (Dmed. - P inf.) - sunt sarcopterygieni cu caractere apropiate de cele ale tetrapodelor primitive (amfibieni): craniul foarte lat si scund; orbitele dispuse dorsal; orificiul pineal dispus in spatele orbitelor; septum internazal gros; structura scheletului de sustinere a inotatoarelor perechi, relativ usor de omologat cu structura membrelor tetrapodelor (Fig. 16).
|
Figura 16. Eusthenopteron foordi, Devonian
superior (Frasnian), Quebec, Canada (vestul Laurasiei). Lungime = |
Tetrapodomorphele sunt vertebrate adaptate primar locomotiei pe uscat, locul inotatoarelor neperechi fiind luat de cele doua perechi de membre. Tetrapodomorphele reprezinta tetrapode primitive, mai cunoscute sub numele de amfibieni. Sunt organisme ectoterme, a caror desprindere de mediul acvatic este doar partiala, dezvoltarea embrionara desfasurandu-se in mediul acvatic, iar adultii traiesc in preajma apelor sau in locuri cu umiditate ridicata. Tetrapodele au evoluat din pestii sarcopterygieni. Cele mai vechi tetrapodomorphe cunoscute sunt genurile Acanthostega (Fig. 17) si Ichthyostega (Fig. 18) din Devonianul din Groenlanda. Aceste doua genuri prezinta o serie de caractere care marcheaza trecerea de la sarcopterygieni la tetrapodele primitive.
|
Figura 17. Acanthostega (D sup.) |
Figura 18. Ichthyostega (D sup.) |
REPTILOMORPHA (C-T)
Reprezinta un grup ancestral de tetrapode care marcheaza tranzitia dintre tetrapodomorphe si reptile. Cuprinde doua clade majore: Anthracosauroidea (C-T) si Batrachosauria (C-P).
Caractere generale: majoritatea sunt forme terestre, dar si acvatice; bolta craniana slab atasata de partile laterale; osul tabular mare atasat la parietal; centuri bine dezvoltate; membrele cu 5 degete; unghi ascutit intre bolta craniana si partile laterale ale craniului, sutura posibil mobila.
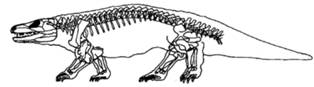
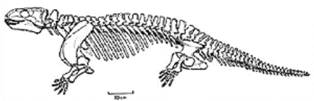
De retinut cladul Bathracosauria: cuprinde doua clade majore: Seymouriamorpha (C-P) si Cotylosauria (C-P).
Seymouriamorpha - (C-P) clad reprezentat prin genul Seymouria (P, SUA) (Fig. 19): Lungime aproximativ 70cm; Prezinta caractere tipice de stegocephali si caractere reptiliene ilustrand in ansamblu modalitatea tranzitiei structurale de la tetrapodomorphe (amfibieni) la reptile.
Cotylosauria (Csup.-Pinf.) - considerate reptile ancestrale (incluse un sistematica veche la Subclasa Anapsida-Cls. Reptilia). Aveau o lungime cuprinsa intre 1,5-3m; unele erau complet terestre, ierbivore (g. Diadectes). (*) Genul Diadectes (Fig. 20) - avea santul otic cu un tympan osificat, 3m lungime; centuri puternice, iar vertebrele aveau articulatii intervertebrale suplimentare; dintii premaxilari si cei anterior dentari erau spatulati.
|
Figura 19. Seymouria (P) - bathrocosaurian cu caractere tipice de stegocephali si caractere reptiliene |
|
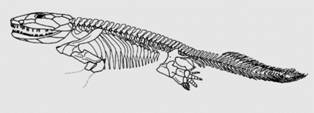
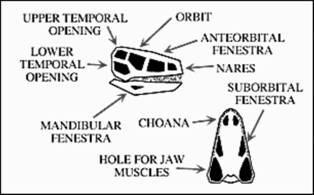
Amniotele sunt animale care depun oua, de milioane de ani. Embrionul se dezvolta in interiorul oului. Oul amniotelor are la exterior o "cochilie", un invelis dur (carbonatic) care previne uscarea, iar in interior embrionul este inconjurat de mai multe membrane. Acest tip de ou este unic, caracteristic amniotelor care includ: broastele testoase, soparlele, dinosaurii, pasarile si mamiferele. Ar putea sa va surprinda faptul ca in aceasta lista de aminote sunt trecute si mamiferele (despre care stim ca in general sunt vivipare, nasc pui vii). Totusi cele mai vechi mamifere (mamiferele primitive) depuneau oua si cateva mamifere actuale (monotreme) mai fac acest lucru si astazi.
Figura 21. Limnoscelis (Pinf.) - probabil un amniot bazal.
(*)In interiorul oului se afla o serie de membrane separate prin fluide, care permit embrionului sa supravietuiasca si sa se dezvolte. Aceste membrane sunt: amnionul, allantoida, sacul yolk si chorionul. In jurul embrionului se afla amnionul care este umplut cu lichid amniotic. Urmeaza allantoida care indeplineste 2 functii foarte importante pentru embrion: permite difuzia gazelor si indeparterea deseurilor. Hrana pentru dezvoltarea embrionului provine din sacul yolk, care se reduce ca marime, pe masura ce embrionul se dezvolta. La exterior, toate aceste membrane sunt acoperite de chorion. In jurul chorionului se afla albumina si o cochilie externa, coaja, protejeaza intregul ou prevenind uscarea, dar in acelasi timp permitand aerului sa ajunga la embrion (prin pori).
Placenta este un "ou modificat". (*)La mamiferele placentare, membranele pe care le gasim la ou, sunt modificate. Embrionul este in continuare inconjurat de fluidul amniotic. Allantoida si sacul yolk se transforma in cordonul ombilical-conexiunea prin care hrana ajunge la fetus si rezidurile sunt indepartate. Aceste membrane impreuna cu o parte din chorion formeaza placenta care este atasata de peretele uterin al mamei.
Tetrapodele primitive erau legate de mediul acvatic prin mecanismul lor reproductiv. Ca majoritatea pestilor acestea aveau fertilizare externa in apa, unde depuneau ouale din care se dezvolta larva acvatica. Prin metamorfoze succesive se transforma treptat in adulti care traiau pe uscat. Unii "amfibieni" actuali au mostenit acest mod de reproducere primitiv. In timpul Carboniferului insa, un grup de tetrapode (Bathracosauria-Cotylosauria) au dezvoltat oul amniotic-care reprezinta un caracter derivat major, comun tuturor amniotelor.
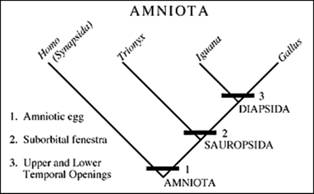
Imediat dupa aparitia primelor amniote pot fi recunoscute doua grupe distincte de amniote:
-Sauropsida - linia de evolutie a pasarilor
-Synapsida - linia de evolutie a mamiferelor.
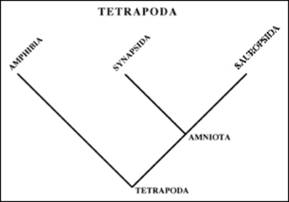
Cladele majore de Tetrapode si principalele lor synapomorfoze sunt (Fig. 22):
-Tetrapode - 4 membre
-Amniote - ou amniotic
-Lissamphia - (amfibienii actuali)- dinti pe piedestale
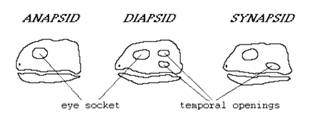
-Synapside- fereastra temporala inferioara
-Sauropside- fereastra suborbitala
|
|
|
||||||
|
Figura 22. Cladogramele tetrapodelor si amniotelor si principalele synapomorfoze. |
Amniotele sunt cunoscute in stare fosila incepand din Carbonifer. Desi nu avem inca nici o dovada directa a oului amniotic inainte de Mesozoic, totusi amniotele paleozoice trebuie sa fi avut un ou similar din moment ce toti descendentii acestora au un astfel de ou. Amniotele sunt divizate in doua mari grupe: SAUROPSIDE - linia pasarilor SYNAPSIDE - linia mamiferelor.
DISTRIBUIE DOCUMENTUL
Comenteaza documentul:Te rugam sa te autentifici sau sa iti faci cont pentru a putea comentaCreaza cont nou Termeni si conditii de utilizare | Contact
| ||||||