| CATEGORII DOCUMENTE |
| Astronomie | Biofizica | Biologie | Botanica | Carti | Chimie | Copii |
| Educatie civica | Fabule ghicitori | Fizica | Gramatica | Joc | Literatura romana | Logica |
| Matematica | Poezii | Psihologie psihiatrie | Sociologie |
Stomatele
Stomatele Stomatium) sunt structuri specializate, localizate la nivelul epidermei, avand rol in schimbul de gaze dintre planta si mediul extern, precum si in degajarea vaporilor de apa (transpiratia).
Ele nu sunt altceva decat elementele epidermice cele mai externe ale tesuturilor aerifere. Cu ajutorul lor mai are loc si patrunderea in corpul plantei a bioxidului de carbon necesar pentru asimilatie si a oxigenului necesar respiratiei, precum si eliminarea din corpul plantelor a diferitelor substante gazoase (oxigen si bioxid de carbon) formate in decursul proceselor amintite mai sus.
Prin organizarea lor se deosebesc de celulele epidermice.
O stomata este formata din doua celule stomatice reniforme, dispuse fata in fata cu suprafata lor concava, lasand intre ele o deschidere numita ostiola.
Celulele stomatice prezinta structura, continut si functii caracteristice.
Fata de celulele epidermice propriu-zise, ele contin multa citoplasma, nucleu clar si cloroplaste dispuse in jurul nucleului.
Pe langa cloroplaste, se disting mici grauncioare de amidon.
Celulele stomatice sunt inconjurate de doua sau mai multe celule anexe, lipsite de cloroplaste: indeplinesc legaturi functionale si de protejare a celulelor stomatice. Fiecare celula stomatica poseda:
v un perete ventral, ingrosat, spre ostiola;
v un perete dorsal, mai subtire, celulozic, spre celula anexa;
v un perete extern, acoperit cu o cuticula mai groasa sau mai subtire, spre exterior;
v un perete intern, mai subtire, celulozic.
Peretii ventrali au doua creste, una externa si alta interna. Uneori sunt mai dezvoltate crestele externe, ingrosate, cutinizate. Alipirea lor poate inchide complet ostiola.
Ambele creste externe delimiteaza atriul superior, reprezentand deschiderea izodiala, iar crestele interne delimiteaza atriul inferior ,
reprezentand deschiderea opistiala.Uneori, celulele stomatice se gasesc scufundate printre celulele epidermice rezultand camera suprastomatica. Rolul ei este de a impiedica o transpiratie puternica. In toate cazurile, sub cele doua celule stomatice se afla delimitat un spatiu intercelular numit camera substomatica. Aceasta se afla in legatura cu toate spatiile intercelulare si constituie un sistem de aerisire. Ansamblul celulelor stomatice cu celulele anexe alcatuiesc aparatul stomatic.
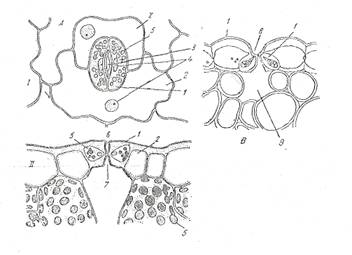
Fig. Alcatuirea stomatei
A - frunza de cimbrisor (Thymus serpyllum); B - stanjenel (Iris germanica); I - vazuta din fata; II - sectiune transversala; 1 - stomate; 2 - celule anexe; 3 - ostiola; 4 - nucleu; 5 - cloroplaste, 6 - atrium superior; 7 - atrium inferior; 8 - camera suprastomatica; 9 - camera substomatica.
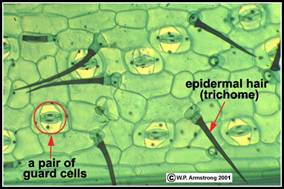
Fig. - Stomatele la Tradescantia pendula (Dupa Armstrong)
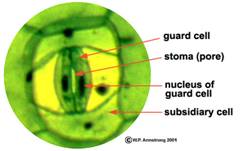
Fig. - Alcatuirea unei stomate de Tradescantia (Dupa Armstrong)
Distributia si numarul stomatelor
Stomatele sunt distribuite in general pe toate organele aeriene ale plantelor, de exemplu frunze, tulpini tinere, sepale, petale, etc. Nu au stomate plantele terestre saprofite, unele plante acvatice submerse, etc.
Pozitia stomatelor pe frunza este dependenta de diverse adaptari ecologice.
Exista frunze amfistomatice, orientate in spatiu vertical sau aproape vertical, cu stomate in epiderma ambelor fete. De exemplu, prezinta astfel de frunze stanjenelul(Iris germanica), porumbul(Zea mays), graul(Triticum aestivum), ovazul(Avena sativa), etc.
Un alt tip de frunze sunt frunzele hipostomatice, orientate in spatiu mai mult sau mai putin orizontal, cu stomate numai in epiderma fetei inferioare. Astfel de frunze se intalnesc la specii precum:fagul(Fagus sylvatica), stejarul(Quercus robur), teiul(Tilia tomentosa), marul(Malus domestica),etc.
Exista si frunze epistomatice, cu stomate numai in epiderma fetei superioare, caracteristice plantelor acvatice cu frunze natante, asa cum sunt de exemplu: nufarul alb(Nymphaea alba), nufarul galben(Nuphar luteum), broscarita(Potamogeton natans) si latele.
In general predomina frunzele amfistomatice, dar si la ele stomatele sunt in numar mai mare pe fata inferioara a frunzelor. Frunzele hipostomatice si epistomatice au un numar relativ restrans de stomate si pe fata superioara cam una pana la doua stomate de mm. Numarul cel mai frecvent de stomate la frunzele plantelor e cuprins intre 100 - 300 pe mm. Sunt cazuri cand stomatele sunt mai numeroase, de exemplu, la rapita sunt 600-700 pe 1 mm, la Spiraea ulmaria sunt 1200 pe mm. O frunza de rapita are 11.000.000 stomate si una de floarea-soarelui 13.000.000.
Plantele cu frunze plutitoare, de ex., nufarul, au stomatele dispuse numai pe fata superioara a frunzelor in numar de 450 pe mm.
Stejarul are 300 pe mm, cerealele au 30 - 90 pe mm, iar Sempervivum tectorum are numai 11-14 pe mm.
Plantele submerse ca Elodea, nu au stomate deloc. Altele, ca Potamogeton si Zannichellia palustris au stomate, dar ele sunt in permanenta inchise.
Deseori frunzele arborilor nu au stomate pe fata superioara. Distributia stomatelor si numarul lor prezinta variatii in functie de mediul unde traiesc plantele si de intensitatea transpiratiei (Shearman, Beard 1972).
Nivelul la care sunt asezate stomatele variaza in functie de conditiile de viata in care traiesc plantele. Numeroase plante au stomatele asezate la nivelul celulelor epidermice si nu poseda camera suprastomatica. De exemplu: spanzul (Helleborus), zambila (Hyacinthus), narcisa (Narcissus), etc. Acestea traiesc in conditii de umiditate normala.
Unele plante ce cresc in astfel de conditii au stomatele usor iesite in afara fata de epiderma asa cum este dovleacul (Cucurbita pepo). Plantele caracteristice regiunilor secetoase, sarace in apa, au stomatele scufundate fata de nivelul celulelor epidermice si, in acest caz, se formeaza o camera suprastomatica care contribuie la micsorarea transpiratiei. Este cazul unor specii precum:Agave, Aloe.
Numarul stomatelor pe limbul frunzei variaza in functie de specie.
Numarul stomatelor variaza si pe frunzele aceluiasi individ, astfel cele superioare au mai multe stomate decat frunzele inferioare (Shearman, Beard 1972).
Stomatele la frunzele cu nervuri paralele (gimnosperme si monocotiledonate) sunt dispuse in randuri paralele, iar la frunzele cu nervuri reticulate stomatele nu sunt dispuse intr-o anumita ordine.
Celulele stomatice pot fi dispuse la acelasi nivel cu celulele epidermice, pot sa iasa deasupra lor sau pot fi distribuite sub nivelul epidermei. La unele plante stomatele se gasesc in niste adancituri ale frunzei (stomate profunde). In aceste cripte (adancituri) se remarca existenta unor perisori epidermici.
Tabelul Numarul stomatelor pe mm la diferite specii de plante (Heller, 1965)
|
Specia |
Frunza |
Specia |
Frunza |
||
|
Suprafata superioara |
Suprafata inferioara |
Suprafata superioara |
Suprafata inferioara |
||
|
Ovaz |
Geranium sp. | ||||
|
Grau |
Liliac | ||||
|
Porumb |
Mar |
| |||
|
Tomate |
Cires | ||||
|
Fasole |
Piersic | ||||
|
Varza |
Plop | ||||
|
Cartofi |
Stejar | ||||
|
Lucerna | |||||
In general, pe un milimetru patrat pot exista 10 stomate ca la Sedum acre sau 1200, ca la cretusca (Filipendula ulmaria). Pe lamina frunzelor de floarea-soarelui (Helianthus annuus) numarul stomatelor este de ordinul milioanelor. In epiderma petalelor numarul stomatelor este foarte scazut. Frunzele submerse nu au stomate asa cum sunt: ciuma apelor (Elodea canadensis), sarmulita apei (Vallisneria spiralis).
Stomatele frunzelor xerofitelor sunt mai mici si mai numeroase pe unitatea de suprafata decat ale mezofitelor si la cestea mai mici si mai numeroase decat la higrofite.
In general, numarul stomatelor este mai mare la plantele lemnoase si la acelea care cresc intr-un mediu uscat si luminat, fata de plantele erbacee si acelea care traiesc intr-un mediu umed si intunecos(Gabriela Serbanescu - Jitaru, C. Toma, 1980).
Forma celulelor stomatice in functie de planul de inchidere a celulelor poate fi semilunara, haltera sau reniforma.
Structura celulelor stomatice
Ele au un protoplast activ format dintr-un nucleu, iar in citoplasma intra: cloroplaste, vacuole, aparatul Golgi, etc. O particularitate extrem de caracteristica a structurii stomatelor o prezinta membrana care are o ingrosare neuniforma. Aceasta este in legatura cu modificarea formei si a volumului celular in procesul de inchidere si de deschidere a stomatelor.
Stratul extern al celulelor stomatice de obicei este cutinizat, iar peretele celular este acoperit cu cuticula. Cuticula uneori acopera celula stomatica si la nivelul ostiolei sau poate sa ajunga pana la nivelul camerei substomatice.
Ultrastructura
In procesul diferentierii celulelor stomatice la Pisum sativum se ingroasa mult peretele tangential inferior si superior, dar si peretele anticlinal care margineste deschiderea stomatei. Ingrosarea peretilor tangentiali se caracterizeaza printr-o dispunere radiara a fibrilelor celulozice in directia deschiderii stomatelor, spre peretele anticlinal.
In citoplasma periferica, langa peretii care se ingroasa, s-a constatat existenta numerosilor microtubuli avand aceeasi distributie radiara ca si fibrilele celulozice. Celulele mame din care se formeaza stomatele se aseamana cu celulele epidermice din punct de vedere ultrastructural. Ele sunt formate din vacuole mici, reticul endoplasmatic (RE), mitocondrii, plastide, dictiozomi si ribozomi. La celulele stomaticein diferentiere densitatea citoplasmei scade, creste numarul de mitocondrii si dictiozomii (Singh, Srivastava 1973). Pe masura diferentierii celulelor stomatice dimensiunile plastidelor cresc formandu-se in interiorul lot tilacoizii, acumulandu-se totodata si amidon. La frunzele sempervirescente, amidonul este inlocuit cu uleiuri grase, tanin sau zaharuri simple.
La maturitate celulele stomatice au una sau mai multe vacuole de marimi variabile. Ele in mod frecvent sunt interconectate si contin incluzii veziculare.
Cercetari de microscopie electronica asupra structurii stomatelor de la Anemia rotundifolia arata ca stomatele deschise sunt insotite de marimea volumului vacuolar si dezorganizarea partiala a citoplasmei (Humbert, 1971). In citoplasma celulelor stomatice se observa numeroase vezicule si cordoane ale reticulului endoplasmatic. Nucleul este dispus oarecum central fiind acoperit de doua membrane de tip elementar. Ca o particularitate citologica esentiala a stomatelor o constituie ingrosarea inegala a peretilor celulari. Peretii laterali ai celulei stomatice sunt subtiri, iar cei ventrali de langa ostiola sunt grosi.
Folosind microscopul electronic in cercetarea structurii peretilor stomatei la Helleborus nigrum s-a constatat o structura poroasa a peretilor ingrosati, iar membrana ventrala a celulei stomatice este prevazuta cu coaste mai mult sau mai putin proeminente.
Originea si ontogenia stomatelor
Pentru cercetarea modului de formare a stomatelor este indicat sa se cerceteze primordiile foliare la plante. Dupa un timp scurt de la aparitia primordiei, unele celule epidermice incep a forma celulele stomatice.
Asadar stomatele isi au originea in celulele epidermice. Prin simpla diviziune a unei celule initiale sau celule mama a stomatei (Teodorescu, 1973) rezulta doua celule inegale: o celula mica, reprezentand celula-mama stomatica este opaca, bogata in citoplasma, continand leucoplaste fara granule de amidon, mitocondrii, multi ribozomi, reticul endoplasmatic granular, vacuole marunte si un nucleu sferic, mai voluminos decat al celulei mari ce va forma celula anexa. Celula-mama stomatica se rotunjeste si, prin aparitia unui perete longitudinal, rezulta cele doua celule stomatice.
Peretele longitudinal se ingroasa capatand forma lenticulara. Ostiola se formeaza pe cale schizogena, prin dezintegrarea substantei pectice a peretelui longitudinal. Ea este o deschidere intercelulara reglabila prin care se face posibil schimbul de gaze intre spatiile intercelulare si mediul extern. Celulele stomatice si celulele anexe pot rezulta din una si aceeasi celula initiala( stomate de tip schizotecheil). In alte situatii celulele anexe se formeaza prin diviziuni ale unor celule epidermice tinere si vecine cu stomatele(Stomate de tip haplocheil).
Dupa modul de diferentiere a stomatelor se deosebesc 4 tipuri:
O singura celula mama (stomatica) din epiderma va forma direct o stomata, iar celulele din vecinatatea ei nu se mai divid. Acestea nu au celule anexe, de exemplu la monocotiledonate (Iris, Hyacinthus). La un moment dat, pe frunza tanara de Iris, unele celule epidermice se divid inegal, formand doua celule, una din ele este mica si alta mare. Celula mica este de fapt celula mama a stomatei, deoarece din ea se vor forma printr-o diviziune celulele stomatei. A doua celula, mare, va functiona ca celula epidermica. Celulele stomatice la Iris sunt lipsite de celule anexe.
La formarea stomatelor la zambila in celula-mama a stomatei se formeaza un perete despartitor circular si astfel are loc formarea unei celule-fiica care apoi se divide in doua, dand nastere la doua celule stomatice.
La Tradescantia, celula mama a stomatei se formeaza ca la Iris, apoi ea se divide si formeaza celulele stomatice, dar spre deosebire de Iris, in timpul formarii stomatei, celulele invecinate se divid formand 4 celule anexe.
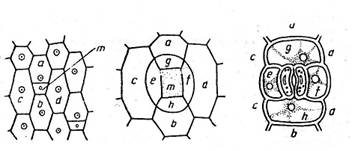
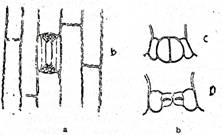
Fig. Originea stomatelor la Tradescantia virginica; stomatele sunt inconjurate de patru celule anexe (dupa Grintescu)
a,b,c,d - celule epidermice; e,f,g,h - celule anexe; m - celule - mama
A treia modalitate arata ca o celula mama a stomatei prin diviziune va forma mai intai celulele anexe si apoi, in ultimul stadiu, se vor diferentia celulele stomatice, exemplu la Camelina.
O celula mama a stomatei se va divide prin patru pereti oblici si va forma 4 celule. Una din celule se va mai divide o data si va forma stomata, iar celelalte 3 se vor transforma in celule anexe. Acest mod de formare a stomatei este caracteristic pentru reprezentantii familiei Cruciferae si Crasulaceae.
Originea si ontogenia celulelor stomatice s-au cercetat cu microscopul electronic la Lactuca sativa (Plant si colab., 1972), Papaver si Fumaria (Kidwai, 1972), Sinapis alba (Leandr, 1972), Ephedra, Gnetum (Imandar, Bhatt, 1972), Avena sativa (Kaufman si colab., 1970), etc.
Initial, celula mama stomatica se afla la acelasi nivel cu celulele epidermice. In timpul diferentierii stomatelor apare schimbarea de pozitie, ridicandu-se sau colaborandu-se fata de nivelul epidermei. Dupa ce s-au diferentiat celulele stomatice are loc formarea ostiolei in mod schizogen, cat si prin gelificarea lamelei mediane dintre cele doua celule stomatice tinere. Apoi se instaleaza procesul de ingrosare inegala a peretilor celulari ai stomatei. Tot acum are loc si formarea camerei substomatice.
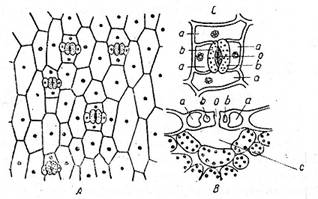
Fig. Tradescantia virginica
A - fragment din epiderma unei frunze cu 5 stomate; B - stomata in sectiune;
C - vazuta de sus; a - celule anexe; b - celule stomatice; c - camera substomatica; o - ostiola (B si C dupa Strasburger).
Pe o frunza stomatele nu apar toate deodata, ci succesiv pe masura cresterii limbului foliar. La plantele cu nervuri paralele care au stomatele dispuse in siruri longitudinale, etapele de dezvoltare a stomatelor sunt observate in zonele de crestere activa a frunzei, iar la frunzele cu nervatiuni reticulate (majoritatea dicotiledonatelor) stadiile diferite ale dezvoltarii stomatelor sunt amestecate ca intr-un mozaic, de exemplu stomatele tinere apar printre cele mature. La criptogame vasculare se intalnesc ambele modalitati.
Dupa modul de repartizare a celulelor anexe exista mai multe tipuri:
Anemocit - celule neregulate de tip ranunculaceu. De fapt stomata este inconjurata de cateva celule care nu se deosebesc dupa forma si marime de restul celulelor epidermice. Prin urmare sunt lipsite de celule anexe tipice.
Anisocit - celulele anexe au marimi diferite (tip crucifer). De data aceasta stomata este inconjurata de 3 celule anexe, din care una este foarte mica.
Paracit - cu celulele anexe paralele (tip Rubiaceae). Asadar stomata este insotita de ambele parti de cate o celula anexa sau mai multe, al caror ax este paralel cu deschiderea ostiolei si a celulelor stomatice.
Diacit - celulele anexe dispuse in cruce (tip Cariofilaceae). Stomata este inconjurata de doua celule anexe care sunt dispuse perpendicular (in unghi drept) pe celulele stomatice.
Encicloid - celulele anexe sunt dispuse sub forma de inel, in jurul celulelor stomatice.
Gradul de deschidere a stomatelor
In actualitate este foarte important de cunoscut gradul de deschidere a stomatelor, deoarece intensitatea transpiratiei depinde de aceasta. Cele mai importante metode de determinare a deschiderii stomatelor sunt:
Metoda microscopica se foloseste mai ales pentru frunzele subtiri si transparente care au stomatele situate la nivelul epidermei, in acest scop utilizandu-se microscopul cu micrometrul ocular.
Metoda infiltratiei se foloseste mai ales pe teren pentru stabilirea gradului de deschidere a stomatelor. In acest scop se aseaza pe fata inferioara a frunzei o picatura de benzina sau benzol si observam frunza spre lumina. Aparitia unei pete de culoare mai inchisa denota ca lichidul respectiv a patruns in spatiile intercelulare ale mezofilului, indicand ca stomatele sunt desschise.
Porometrul este un aparat care permite aprecierea gradului de deschidere a ostiolelor, dupa rezistenta intampinata de aer la strabaterea frunzei.
Functiile celulelor stomatice
La plante celulele stomatice indeplinesc mai multe functii.
Transpiratia la plante se face cu precadere prin stomate, dar mai are loc si prin cuticula, suber si lenticele. Se stie ca difuziunea gazelor prin suprafete mici are loc dupa formula lui Fick:
F - f
V = A ― * S
d
unde V reprezinta viteza de difuziune, A - o constanta a difuziunii, F si f este concentratia substantei respective, d - distanta de difuziune, iar S reprezinta suprafata. Asadar, la aceeasi substanta viteza difuziunii este direct proportionala cu suprafata si cu diferenta de concentratie si invers proportionala cu distanta. In situatia suprafetelor mici (stomate) se remarca abateri de la relatia lui Fick, care constau in aceea ca prin foarte multe suprafete mici difuzeaza o cantitate mai mare de vapori decat prin deschideri mari care au in total aceeasi suprafata. Asa de exemplu, la un vas descoperit vaporii de apa se distribuie in mediul inconjurator in cazul deschiderilor (porilor) mici, prin mijlocul orificiului iese un numar relativ mic de molecule de apa, insa numarul cel mai mare iese pe la marginea orificiului, deoarece aici aerul este mai putin saturat in vapori de apa. In aceasta situatie difuziunea are loc dupa relatia lui Stephan:
F - f
V = 4 * r * A -
p
unde V reprezinta viteza evaporarii, r - raza suprafetei mici, A - constanta de difuziune, F - tensiunea vaporilor saturati, f - tensiunea vaporilor de aer in momentul respectiv, iar p - presiunea atmosferei. Asadar, in jurul fiecarei stomate care are ostiola deschisa vaporii de apa iesiti prin transpiratie formeaza o atmosfera saturata in vapori. Spre a nu se frana functia de transpiratie distanta dintre ostiole este de aproximativ 10 ori mai mare ca diametrul lor.
Asupra deschiderii stomatelor actioneaza anumiti factori. Astfel, stomatele reactioneaza in scurt timp (cateva minute) la actiunea unor factori ai mediului inconjurator, ca de exemplu: intensitatea luminii, lungimea de unda a radiatiilor luminii, starea de aprovizionare cu apa a frunzelor, leziunile de pe frunza, acizii.
Influenta intensitatii luminii
Stomatele, la intuneric, in mod obisnuit sunt inchise. In cazul iluminarii frunzelor au loc miscari de deschidere a ostiolelor (miscare fotonastica). Deschiderea este cu atat mai prompta cu cat lumina este mai intensa.
Actiunea luminii monocromatice
Daca frunzele plantelor sunt puse in lumina albastra si rosie de aceeasi intensitate se constata ca stomatele se deschid mai degraba in lumina albastra decat in cea rosie. Dimpotriva, intensitatea fotosintezei este mai mare in lumina rosie fata de cea albastra, de aici se poate intelege ca la baza mecanismului de deschidere a stomatelor nu este fotosinteza. Asadar, in mecanismul deschiderii stomatelor lumina intervine ca un excitant.
Aprovizionarea cu apa
In cazul unei bune aprovizionari a frunzelor cu apa, stomatele sunt deschise, iar daca se produce un deficit de apa stomatele se inchid inainte de a se instala vestejirea.
Leziunile de pe frunza produc deschiderea stomatelor din apropierea ranii, determinat de faptul ca celulele anexe ranite preseaza mai putin pe celulele stomatice facilitand miscarea pasiva de deschidere a ostiolelor.
Substantele chimice si pH-ul actioneaza asupra deschiderii stomatelor. S-a constatat ca acidul acetic diluat stimuleaza deschiderea stomatelor si ca el actioneaza prin modificarea pH-lui. Stomatele se inchid la un pH intre 5-5,5 si se deschid la o crestere sau o descrestere fata de aceste cifre. In celulele stomatice pH-ul se modifica sub actiunea CO . La lumina cantitatea de CO scade, iar pH-ul din celulele stomatice creste, ceea ce faciliteaza deschiderea ostiolelor.
In general se stie ca stomatele au o functionare ritmica, asa de pilda stomatele de pe frunzele celor mai multe plante se deschid la lumina si se inchid la intuneric. Ziua sunt deschise, spre seara si noaptea sunt inchise.
Exceptie fac unele specii (Anemone silvestris, Bellis perenis), la care raman deschise si in timpul noptii. Mecanismul deschiderii lor consta in hidrolizarea amidonului preexistent din stomate, sub influenta unor enzime care actioneaza la ivirea luminii. Sucul celular al stomatelor devine concentrat in glucoza, substanta osmotic activa. Apa din celulele epidermice vecine trece in celulele stomatice, care devin turgescente.Sub influenta turgescentei, peretii subtiri se intind si trag dupa ei peretii ingrosati ce delimiteaza ostiola, deschizand-o. Spre seara, glucoza se transforma in amidon, substanta osmotic inactiva. Ca urmare, are loc trecerea apei din stomate in celulele epidermice, turgescenta scade si ostiolele se inchid.
Miscarile de deschidere si de inchidere ale stomatelor
Celulele stomatelor executa miscari de apropiere si de indepartare intre ele, modificand prin aceasta dimensiunea ostiolei.
Dupa modul cum celulele stomatice presteaza miscari de inchidere si de deschidere exista 3 tipuri de aparate stomatice:
I. Tipul amarilidaceu este intalnit la Amaryllidaceae, cat si la majoritatea monocotiledonatelor si dicotiledonatelor. Celulele stomatelor de acest tip privite din fata au forma boabelor de fasole prinse intre ele prin peretele lor concav, cum se vede in fig. . In jurul lor se gasesc celulele anexe, care sunt de fapt celule ale epidermei.
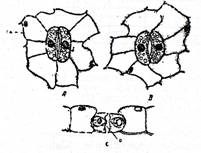
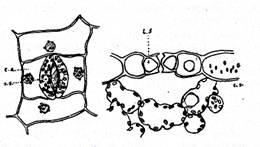
Fig. - Stomate ale frunzei marului A si B vazuta din fata si C, in sectiune transversala; c.a. celule anexe, o - ostiola. (Dupa B. A. Rubin)
Cele doua celule stomatice de forma reniforma (privite de sus) executa miscari de apropiere si de indepartare intre ele intr-un plan paralel cu suprafata epidermei, modificand prin aceasta dimensiunea ostiolei. Membrana celulelor stomatelor este subtire si elstica pe peretele dinspre celulele anexe, cum se vede in fig. , atat la stomata privita din fata A, cat si la cea sectionata transversal B.
A B
Fig. - Stomata de pe fata inferioara a frunzei de Tradescantia virginica. A - vazuta din fata si B - vazuta in sectiune transversala
Aceste stomate au celulele stomatice cu peretii dorsali mai subtiri, iar cei ventrali dinspre ostiola sunt ingrosati mai ales spre partea inferioara si superioara. Aceste doua proeminente separa ostiola intr-o camera anterioara sau vestibul, care comunica cu aerul inconjurator si intr-o camera posterioara sau atriu, care stabileste comunicarea cu camera substomatica. Intre cele doua ingrosari se afla o zona a peretelui ventral subtire. Peretii ingrosati ai celulelor stomatelor sunt cu mult mai putin extensibili decat peretele subtire dinspre celulele anexe.Din cercetari microscopice reiese ca elasticitatea acestor pereti se datoreaza dispunerii microfibrilelor de celuloza care au o orientare radiara. Extensibilitatea maxima este perpendiculara pe directia microfibrilellor.Celulele stomatice functioneaza cu ajutorul apei din sucul vacuolar si al celulelor anexe.
Celulele stomatelor au un strat de protoplasma, care captuseste membrana. In aceasta se gaseste un nucleu si numeroase cloroplaste verzi. In celulele epidermei cloroplastele lipsesc. Cea mai mare parte a volumului celulelor stomatelor, este ocupata de o vacuola, care contine saruri si glucide dizolvate in sucul sau.
Modul de funcsionare al acestui tip de stomate a fost explicat de Schwendener. El se intelege usor cu ajutorul unei stomate artificiale, construita din doi saci de cauciuc, avand peretele intern ingrosat si peretele extern subtiat. Umfland acesti saci cu aer, ei se curbeaza in afara, peretele extern mai extensibil devenind convex, producand o ccurbura concava la peretele intern mai putin extensibil. Prin aceasta, distanta dintre cei doi saci de cauciuc se mareste.
In cazul stomatei, modificarea volumului celulelor se produce cu ajutorul apei, care atunci cand este supta de celulele stomatei din celulele anexe, le mareste volumul si prin aceasta produce incovoierea convexa a peretelui subtire si elastic dinspre celulele anexe si incovoierea concava a peretelui dinspre ostiola, mai gros si mai putin elastic, cum se vede in fig. , IV si VI.
I III VI
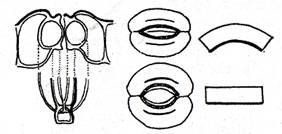
II IV V
Fig. - Stomata de la frunza de Amaryllis formosissima. I - sectiune transversala; II - jumatate a stomatei privita din fata; III - stomata inchisa privita din fata; IV - stomata deschisa privita din fata; V si VI - schema peretilor celulelor stomatei in stare inchisa V, si deschisa VI. (Dupa Schwendener)
Inchiderea stomatei e produsa prin sugerea apei din celulele stomatelor de catre celulele anexe. Prin aceasta celulele stomatelor isi micsoreaza volumul, peretii revin la pozitia initiala, Fig. III si V si ostiola se inchide. Ostiola e incchisa mai repede si mai puternic de catre cele doua celule ale stomatei in portiunea lor centrala. Asa cum a aratat Schwendener (1898), inchiderea stomatelor se realizeaza atat prin portiunea centrala a ostiolei, cat si prin extremitatile lor, la intariturile membranei din aceste regiuni, care fie ca se intrepatrund sau se suprapun. Schwendener a aratat ca la stomatele inchise se scurteaza vestibulul, in portiunea sa centrala, deschiderea ostiolei lipseste complet, adica inchiderea la acest nivel este completa, iar celulele stomatei au intre peretii lor dinspre celulele anexe, o distanta cu mult mai mica decat la stomatele deschise.
Celulele stomatice sunt complet inconjurate de doua sau mai multe celule anexe. Deschiderea celulelor stomatice depinde si de presiunea pe care o exercita celulele anexe asupra celulelor stomatice. Cu cat aceasta presiune e mai mica, cu atat stomatele se deschid mai uor. Prin inteparea celulelor anexe se inlatura presiunea si stomatele se deschid numaidecat larg. Celulele anexe servesc si ca rezervoare de apa pentru celulele stomatice, caci din ele isi absorb apa in timpul ddeschiderii lor si in ele trece apa din celulele stomatelor in timpul inchiderii lor.
II. Tipul gramineu este raspandit la speciile de Poaceae si Cyperaceae.
La graminee si la criptogame stomatele se deschid tot in directie paralela cu suprafata epidermei, insa ele au peretii ventrali si dorsali ingrosati in zona mijlocie. Asadar, celulele stomatice au forma de haltere, unde zona mediana are peretii foarte grosi, iar capetele distale sub forma de vezicula au peretii subtiri.
La cresterea turgescentei peretii subtiri ai capetelor se intind, iar portiunile mijlocii ingrosate si rigide se indeparteaza deschizand ostiola.
Deschiderea ostiolei este determinata de umflarea capetelor stomatice.
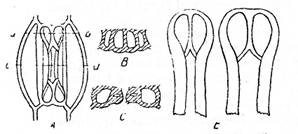
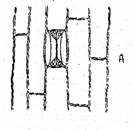
Fig. O stomata de tip Gramineae (dupa Haberlandt)
A - vazuta din fata; B - sectiune transversala in a-b; C - sectiune transversala in c-d; E - mecanismul deschiderii stomatelor.
III. Tipul primitiv (pteridofitic) sau mnium este raspandit la muschi si partial la ferigi.
Modificarea ostiolei are loc prin miscari ale celulelor stomatei, executate intr-un plan perpendicular pe suprafata epidermei. In aceasta situatie peretii dorsali ai celulelor stomatice sunt puternic ingrosati si rigizi, iar cei ventrali sunt subtiri, care au coaste cutinizate. Dar si peretii periclinali sunt subtiri si celulozici. Datorita acestei structuri se poate explica modul de deschidere a celulelor stomatice modificandu-si diametrul lor transversal numai in sens perpendicular pe suprafata epidermei.
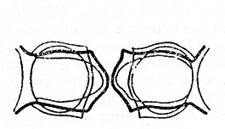
Fig. O stomata de tip 'Mniun' (dupa Haberlandt)
Miscarea de deschidere a stomatelor se face atat paralel, cat si perpendicular pe suprafata epidermei.Celulele stomatice au peretele dorsal subtire, fin, elastic si bombat, iar spre partea externa si interna, prezinta ingrosari sub forma de creste, orientate paralel cu suprafata epidermei. Cresterea turgescentei duce la bombarea peretilor dorsali spre celulele anexe, iar peretii ventrali devin concavi deschizand ostiola.
Fiziologia miscarilor de deschidere si de inchidere a stomatelor
Celulele stomatelor se deosebesc profund de celulele mezofilului frunzelor si e celulele epidermei. Asa, de exemplu, amidonul se acumuleaza la celulele stomatelor la intuneric, ceea ce duce la inchiderea stomatei si dispare la lumina, ceea ce duce la deschiderea stomatei, invers decat cum se intampla la celulele mezofilului si ale epidermei. Si la deshidratare, celulele stomatelor reactioneaza altfel decat ale mezofilului frunzei si anume, la celulele stomatelor deficitul de apa provoaca sinteza amidonului, in timp ce la celulele mezofilului si la celulele epidermei, deshidratarea provoaca hidroliza amidonului. Se cunosc si unele exceptii de la regula stabilita. Asa, de exemplu, sunt stomate care la lumina raman incarcate cu amidon si stomate care in timpul noptii sunt lipsite de amidon sau contin numai urme din aceaasta substanta. La frunzele de muscata s-au observat ca miscarile stomatelor nu insotesc variatia cantitatii de amidon din celulele lor.
In decursul zilei, presiunea osmotica creste mult la celulele stomatelor, in timp ce la celulele epidermei si ale mezofilului frunzei, ea creste numai cu putin, cum se vede in fig. .
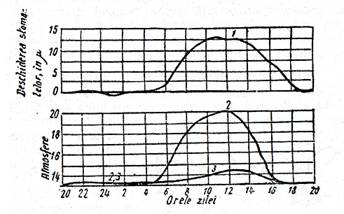
Fig. - Variatia in decursul zilei a deschiderii stomatelor (1) si a presiunii osmotice a celulellor stomatei (2) si ale celulelor epidermei (3).
(Dupa B. A. Rubin)
Cu coloranti vitali, se arata ca celulele stomatelor sunt mai putin permeabile decat celulele anexe, si acestea mai putin permeabile decat celulele epidermei. La celulele omorate, colorantii vitali patrund la fel de repede in toate aceste celule. Celulele stomatelor reduc azotatul de argint, coloranduase in negru. Apa oxigenata coloreaza in brun vacuola celulelor stomatelor, oxidand diferite substante organice. Celulele stomatelor contin enzima fosforilaza, in timp ce celulele epidermei nu cpntin aceasta enzima. Imamura (1943) a aratat ca celulele stomatei acumuleaza ionul potasiu in decursul deschiderii. Celulele stomatelor prezinta o rezistenta mai mare fata de conditiile neprielnice ale mediului inconjurator, ele rezistand mai bine la acizi, la amoniac, la alcool, la eter, la cloroform, decat celulele epidermei. Aceasta deosebire se datoreaza insusirilor protoplasmei. Celulele stomatelor sunt mai rezistente, atat la temperatura ridicata, cat si la temperaturi joase. Dintre toate celulele frunzelor cazute toamna, celulele stomatelor raman mai mult timp in viata. Fata de aceasta regula generala a rezistentei mai mari a celulelor stomatelor, decat a celulelor epidermei, sunt si unele exceptii. Asa, de exemplu, Iljin a aratat ca o seceta puternica vatama mult mai puternic celulele stomatelor, decat celulele epidermei. In timpul iernii, la plantele perene, celulele stomatelor mor in numar mai mare dintre toate celulele frunzelor.
La deschiderea stomatelor, dispare amidonul din celulele stomatelor si reapare la inchiderea lor. Iljin (1914) a aratat ca schimbarea cantitatii de amidon din celulele stomatelor este in stransa legatura cu modificari ale presiunii osmotice. La deschiderea stomatelor creste mult presiunea osmotica din celule, fata de celulele anexe si fata de celulele epidermei, iar la inchidere se revine la situatia anterioara, cand dispar aceste deosebiri. La plante adaptate la lumina puternica, deosebirile intre presiunea osmotica a celulelor stomatelor deschise si inchise sunt cu mult mai mari decat la cele adaptate la lumina scazuta si la umiditate mai mare. Zaharurile, formate prin hidroliza amidonului, sunt polizaharide, dizaharide si monozaharide. Stalfelt a aratat ca presiunea osmotica in celulele stomatelor creste la inceputul expunerii la lumina, dar dupa trei ore ea incepe sa scada. Turgescenta celulelor stomatice creste cu deschiderea lor, asa cum au aratat Ursprung si Blum (1924).
Reactia chimica difera la celulele stomatelor inchise si deschise. Sayr (1926) a gasit ca celulele stomatelor au un pH mai mare in starea deschisa decat in starea inchisa. La lumina pH-ul este cuprins intre 6 si 7,4, pe cand la intuneric el scade sub 5. La celulele epidermei el nu a gasit schimbari. La Rumex acetosa pH-ul celulelor stomatelor la lumina este cuprins intre 5,5 si 6, pe cand la intuneric el este cuprins intre 4 si 5.
Rolul pH-lui in deschiderea stomatelor se explica in modul urmator: la pH 7,0, sub influenta fosforilazei care are un rol insemnat in reglarea schimbului amidon - zahar solubil, fenomenul merge spre dreapta, iar la pH 5,0 el merge spre stanga. La lumina, pH-ul creste repede din cauza scaderii concentratiei CO2, iar la intuneric el scade din cauza formarii CO2. Odata cu cresterea presiunii osmotice, creste si viscozitatea protoplasmei. Aceasta se dovedeste prin forma plasmolizei, care la celulele stomatelor inchise e convexa, iar le celulele stomatelor deschise e concava. Curentii protoplasmici sunt mai mari la celulele stomatelor inchise, ei fiind influentati probabil de viscozitatea protoplasmei. Odata cu modificarea viscozitatii protoplasmei, celulelor stomatei deschise si inchise se schimba si gradul lor de imbibitie cu apa. Probabil ca si modificarile permeabilitatii si ale rezistentei protoplasmei, la conditiile neprielnice, sunt in stransa legatura cu gradul de imbibitie al coloizilor protoplasmei. Probabil se schimba totodata si sarcinile electrice ale coloizilor protoplasmatici. Kiselev a aratat ca celulele stomatelor inchise se coloreaza mai puternic cu coloranti vitali, decat cele deschise si a dedus de aici ca permeabilitatea lor este mai mare. In celulele stomatelor inchise si diastaza poate patrunde in interiorul lor, pe cand, in celulele stomatelor deschise, aceasta enzima nu poate patrunde. Azotatul de potasiu si glucidele patrund mult mai usor in celulele stomatelor inchise. Linsbauer a constatat ca permeabilitatea celulelor stomatelor creste la lumina. Stoeger (1950) a cercetat permeabilitatea mai multor substante, ca amina acetica, glicerina, ureea la stomatele de Tradescantia. El a gasit o permeabilitate mai mare a celulelor stomatelor deschise decat a celor inchise pentru toate substantele cercetate.
Continutul celulelor stomatei se schimba cu deschiderea lor. Cand ele sunt inchise contin de obicei amidon, iar la deschiderea lor amidonul dispare (fig.). O exceptie a gasit Hagen la stomatele frunzelor de ceapa, la care niciodata nu se gaseste amidon. Iljin a aratat ca stomatele deschise nu contin amidon si ca acesta apare cam la ora de la inceputul inchiderii lor. Cloroplastele din celulele stomatelor au mai ales rolul sa inlocuiasca zaharul pierdut in timpul hidrolizei si sa resintetizeze amidonul. Rezistenta celulelor stomatice, in stare deschisa difera de aceea din starea inchisa. Ele rezista mai bine in stare deschisa la temperaturi ridicate, ca si la temperaturi scazute. Rezistenta celulelor stomatice la otravuri este cu mult mai mare in starea deschisa a stomatei decat in starea inchisa. La Vivia faba, la celulele stomatei inchise, nucleii sunt fuziformi, cu ambele capete ascutite. La celulele stomatei deschise, nucleii sunt ovali sau sferici, cu prelungiri in forma de lob. Aceste cercetari s-au facut la epiderma desprinsa de frunze si mai multi cercetatori pun la indoiala modificarile de forma ale nucleului la celulele stomatelor frunzelor intacte.
Mecanismul miscarilor de deschidere si de inchidere a stomatelor a fost explicat din mai multe puncte de vedere:
Teoria fotosintezei
O ipoteza mai veche (Schwendener, 1881) arata ca lumina produce prin intermediul fotosintezei o crestere a presiunii osmotice in celulele stomatice, care faciliteaza patrunderea apei in stomate. Asa s-a crezut ca in celulele stomatice lumina, prin intermediul fotosintezei de la nivelul cloroplastilor, produce o acumulare de glucide in celulele stomatice devenind hipertonice comparativ cu celulele anexe in celulele vecine. In aceasta situatie patrunde apa din celulele anexe in celulele stomatice si datorita elasticitatii inegale a peretilor celulelor stomatice are loc deschiderea ostiolei. Ipoteza fotosintetica nu a rezistat datelor experimentale si anume:
a) La lumina stomatele se deschid intr-un timp scurt, iar pentru sinteza unor cantitati mari de substante organice solubile, capabila sa produca o crestere a presiunii osmotice, este util un timp mai indelungat.
b) S-a aratat ca fotosinteza merge mai intens la lumina rosie, iar asupra deschiderii stomatelor actioneaza mai puternic razele albastre.
c) Celulele stomatice nu realizeaza decat o fotosinteza foarte mica si ca coroplastii ar functiona mai mult ca amiloplaste sintetizand amidon pe seama glucidelor formate in alte parti ale frunzei.
Teoria enzimatica
Arata ca cresterea presiunii osmotice si a turgescentei celulelor stomatice iluminate sunt determinate de enzimele care actioneaza asupra amidonului hidrolizandu-l. In celulele stomatice, la intuneric, enzimele determina sinteza amidonului, care produce o scadere a presiunii osmotice, a turgescentei facilitand inchiderea ostiolelor. Citochimic s-a descoperit ca in celulele stomatice se gaseste fosforilaza, dar aceasta enzima nu se gaseste in celelalte celule epidermice. Acestei enzime i se confera atributul de a hidroliza la pH 6,4 - 7,3, iar cresterea pH-lui este determinata de folosirea CO in fotosinteza, in timp ce la intuneric CO determina o scadere a pH-lui la 5, iar fosforilaza stimuleaza sinteza amilazei.
Teoria enzimatica este sustinuta de urmatoarele date experimentale:
a) Daca se introduce epiderma detasata de pe frunza de Tulipa intr-un extract bogat in amilaza (malt) se obtine o deschidere foarte mare a ostiolelor;
b) Sinteza si hidroliza amidonului se produc intr-un timp foarte scurt, analog cu cel care are loc la deschiderea sau inchiderea stomatelor.
In deschiderea si inchiderea celulelor stomatice intervin miscari fotoactive, hidroactive si hidropasive.
In miscarile fotoactive frunzele sunt aprovizionate suficient cu apa, iar rolul dominant il are lumina, iar lipsa ei produce inchidere.
Miscarile hidroactive sunt determinate de un deficit de apa care stimuleaza actiunea enzimelor din celulele stomatice in sens sintetizant, in timp ce in celulele anexe, epidermice si ale mezofilului enzimele actioneaza in sens hidrolizant. Aceasta situatie duce la o scadere a presiunii osmotice a celulelor stomatice comparativ cu cea din celulele anexe. In acest timp apa este absorbita din celulele stomatice si ostiola se inchide.
Miscarile fotoactive au o importanta mare in fotosinteza prin difuzarea CO spre celulele asimilatoare, in timp ce miscarile hidroactive au rolul sa franeze pierderile de apa prin transpiratie in situatia unui deficit hidric. Dacca assupra frunzei actioneaza concomitent atat lumina, cat si deficitul hidric, domina miscarea hidroactiva inhiband-o pe cea fotoactiva si astfel stomatele se inchid.
Miscarile hidropasive ale stomatelor sunt stimulate de o turgescenta foarte mare a celulelor anexe, care manifesta o presiune asupra stomatelor, facilitand inchiderea ostiolelor.
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 24420
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved