| CATEGORII DOCUMENTE |
| Demografie | Ecologie mediu | Geologie | Hidrologie | Meteorologie |
Parametrii dinamicii populatiilor (relatia input - output). Mortalitate - natalitate; rata mortalitatii - rata natalitatii. Imigrare - emigrare. Crsterea populatiilor, tipuri de rate ale cresterii. Tipuri de modele matematice de crestere a populatiilor. Pareticularitati ale cresterii populatiei
Natalitati si mortalitati excesive. Sunt doua fenomene care la anumite intervale de timp pot afecta semnificativ abundenta populatiilor, aspect care poate fi evidentiat in studiul dinamicii. Practic exista o varietate de cauze care pot influenta cele doua procese. Cele mai credibile sunt cele legate de cantitatea de hrana si daunatori specifici. Alte studii considerau ca variatia ciclica a radiatiei solare, in ritmul anilor cu pete solare (manifestari ale exploziilor si furtunilor solare), este cauza acestor procese biologice. Fapt ulterior infirmat, sau cel putin, pentru anumite specii, el nu a fost dovedit. Indirect toate tipurile de radiatii electromagnetice care ajung pe pamant induc anumite modificari, dar nu influenteaza in asemenea masura fiziologia organismelor. Este adevarat ca plantele isi regleaza ciclurile de dezvoltare in raport cu ciclurile factorilor meteorologici. De exemplu, in zonele nordice, cand este hrana suficienta in verile calde, care alterneaza periodic cu verile reci, productia de fructe a plantelor este maxima. Fiind hrana multa si numarul rozatoarelor este de asemenea mare sau chiar excesiv (Kalela 1961).
In cazul mortalitatii sunt mai multi factori posibili: foamete, factori de mediu excesivi, epidemii, pradatori. La populatii de rozatoare de exemplu, se pare ca nici unul dintre acestia nu are o importanta atat de mare precum relatiile dintre indivizi, de lupta pentru suprematie, dominatie sau existenta (Chitty 1952). Agresivitatea dintre indivizi din perioadele de suprapopulare excesiva pot duce la o diminuare numerica excesiva. La Microtus sp. mortalitatea este mai mare primavara comparativ cu iarna cand populatia este mai mult expusa si frigului si daunatorilor.
Pierderi mici de indivizi sunt frecvente in orice populatie si sunt datorate variatiilor la nivelul factorilor climatici, factori interni (varsta, hrana), factori populationali (relatii intra- si interspecifice), boli. Mortalitatea ale carei cauze au fost sugerate de catre Chitty (1952) nu a fost insa verificata la toate speciile de rozatoare. Pentru alte specii a fost semnalata "boala de soc", un fenomen atipic, in care mortalitatea aparea cauzata nu de afectini evidente ci de un asa-zis fenomen de epuizare subita fara sa fie detactate infectii la nivelul organismului sau alte modificari semnificative le nivelul proceselor fiziologice. Unii specialisti au denumit acest fenomen sindrom adaptiv sau adaptabilitatea organismului (Selye 1956), fiind in realitate vorba de o reactie adecvata a organismelor in urma unor actiuni puternice interne si externe (gen stress) la care organismul se apara in general, pe cale fiziologica-biochimica sau pe cale ecologico-functionala, de-a lungul unui proces constituit din 3 faze importante: faza de alarma, faza de rezistenta si faza de epuizare. (Fig. 5-20).
In populatiile de rozatoare declansarea stress-ului este rezultatul unui comportament agresiv excesiv, fixat genetic si posibil influentat de factorii externi. Suprapopularea determina intensificarea nervozitatii si luptelor pe viata si pe moarte. Stress-ul determina mortalitatea pe urmatoarele cai:
- incapacitatea de aparare pe cale biochimica in faza de epuizare;
- aparitia unor boli incurabile (hipertensiune, scleroza, diabet etc);
- scaderea rezistentei in fata infectiilor;
- diminuarea capacitatii de a mobiliza mecanismele de termoreglare in momente in care factorii climatici devin extrem de nefavorabili
umane si tranzitia demografica.
Parametrii dinamicii populatiilor. Rata natalitatii si rata mortalitatii
Populatia fiind constituita din totalitatea indivizilor unei specii are caracteristici si particularitati dependente de acestia. In timp si spatiu indivizii se nasc si mor, iar intre cele doua extreme, cresc, se dezvolta si se reproduc. Indiferent de aceasta, daca mediul este favorabil populatia va exista.
Indivizii pot intra intr-o populatie prin imigrare sau datorita fenomenului de nastere si parasesc populatia prin mortalitate si emigrare (Fig. 5-23). Daca populatia nu se schimba prin emigrari si imigrari atunci dinamica sa este caracterizata de urmatorii parametrii: rata natalitatii, rata mortalitatii, rata cresterii numerice si ecuatia de crestere.
Orice rata este un raport intre o crestere sau scadere si marimea intreaga fata de care se face raportarea, intr-o unitate de timp.

Fig. 5-23. Reprezentarea schematica a factorilor care modeleaza marimea unei populatii (dupa Brewer)
Daca excludem imigrarea si emigrarea (lucru usor de realizat in laborator), atunci marimea populatiei este dependenta de natalitate si mortalitate. Numarul de indivizi care apar intr-o populatie, ca rezultat al reproducerii (nastere, germinare, diviziune, inmugurire rtc), raportat la o anumita durata de timp constituie rata natalitatii (nx). Numarul de indivizi care dispar din populatie, prin moarte, intr-o anumita unitate de timp constituie rata mortalitatii (qx).
Intr-o populatie, mortalitatea poate fi cauzata de: varsta, vitalitatea scazuta, accidentele (ex. proasta functionare a mecanismelor de maparlire,deformari la aripile adultilor, etc), conditiile fizico-chimice, boli naturale, pradatori, paraziti, agenti patogeni, hrana, uscaciunea, canibalismul, diminuarea capacitatii de refugiu in fata factorilor nefavorabili.
In cercetarile
efectuate asupra populatiilor animale sunt importante datele furnizate de
studiul ciclului biologic sau tabelul de viata (life table) care
reprezinta si o ilustrare a structurii de varsta a populatiei la un moment dat.
Este in realitate un "tabel al natalitatii si mortalitatii" deoarece cuprinde
date statistice referitoare la mortalitatea si supravietuirea in populatie.
Altfel spus este o transpunere tabelara a parametrilor statistici ai populatiei
(Smith, 1974; Collier si colab.,1974; Brewer, 1988; Stiling, 1996) considerand mortalitatea ca probabilitate
de viata sau ca rata a mortalitatii (
Cresterea populatiei. Rata intrinseca si rata finita a cresterii
Definim o populatie ca un grup de organisme care apartin unei aceleeasi specii si ocuun anume spatiu la un moment dat. Atunci cand este necesar a compara diferite populatii, este nevoie de evaluarea si estimarea unor atribute si parametrii care caracterizeaza o anume populatie. Cei mai importanti sunt:
☼ - densitatea = marimea populatie raportata la o unitate de spatiu;
☼ - natalitatea sau rata natalitatii = rata la care sunt adusi in populatie noi indivizi, prin reproducere;
☼ - mortalitatea sau rata mortalitatii = nivelul de indivizi care sunt pierduti din populatie ca urmare a mortalitatii;
☼ - dispersia (dispersal) = rata in care indivizii imigreaza intr-o populatie sau emigreaza din populatie;
☼ - distributia (dispersion) = modul in care indivizii sunt distribuiti in spatiu [am amintit deja de cele 3 tipuri clasice de distributii: distributia randomizata (intamplatoare); distributia uniforma; distributia grupata (in agregate)];
☼ - repartitia claselor de varsta = proportia indivizilor de diferite virste in populatie
In conditiile in care cercetatorul dispune de suficiente date referitoare la ciclul biologic si cunoaste varsta specifica a reproducerii, pot fi determinate o serie de caracteristici ale cresterii populatiilor. Cresterea sau declinul populatiei depinde de distributia ratei natalitatii si mortalitatii in relatie cu varsta, plus o distributie aditionala care reprezinta frecventa relativa a indivizilor din populatie, tot in relatie cu varsta (distributia varstei).
☻- rata intrinseca de crestere sau rata finita (infinitesimala de crestere (r), este diferenta dintre rata natalitatii si rata mortalitatii
Rata intrinseca de crestere este rata de crestere per unitate in conditii fizice specifice, in spatiu nelimitat unde nu este nevoie sa se ia in considerare efectul cresterii densitatii. Multi autori s-au referit la acest concept sub denumirea de potential biotic (prin combinarea ratelor fecunditatii, sexelor si supravietuirii), indice de mediu (ca o masura relativa a stabilitatii mediilor, dar fara a prezenta rata de crestere in diferite conditii de mediu) sau indice combinat (insumand numarul de oua depuse, rata de supravietuire a stadiilor imature, rata de dezvoltare si rata sexelor)(aut.cit in birch
Studiul lui r s-a facut prin metoda de determinare directa, la specii de insecte cu o singura perioada de ovipozitare si care depun toate ouale repede (birch
Orice studiu despre rata de crestere a populatiilor este incomplet fara referire la distributia varstelor in populatie (si legat de aceasta, a capacitatii femelelor de a produce urmasi). O populatie are in realitate un anumit program al varstei atat pentru fecunditate cat si pentru mortalitate. Daca ar exista o populatie cu un program constant al varstei pentru fecunditate si pentru mortalitate, populatia va creste la o rata dN/dt = rN (deci parametrul r se refera la o populatie cu o distributie stabila a varstei). Ideea aceasta a fost vitala in statistica. In oricare alt fel de populatie, rata cresterii variaza in timp pana se obtine o distributie stabila a varstei. Referitor la aceasta, se poate pune intrebarea: care este rata de crestere pentru N0 adulti proaspat emersi, intr-un spatiu nelimitat?. Aceasta rata va varia in timp pana cand populatia va avea o distributie stabila a varstelor. Pe de alta parte, maximul ratei care se poate mentine pe o perioada nedefinita de timp este data de rata de crestere intr-o populatie cu o distributie stabila a varstelor. Aceasta rata este deci o capacitate intrinseca de crestere a organismului.
Toate acestea au determinat pe unii specialisti ecologi in studiul populatiilor de insecte sa respinga forma exponentiala a ecuatiei de crestere, abordand rata de crestere ca un fenomen discontinuu. Totusi, rata de crestere ramane expresia cea mai buna pentru studiul cresterii populatiilor in spatiu nelimitat, dar ea singura nu este totusi satisfacatoare. Astfel, o etapa urmatoare a fost calcularea ratei de crestere tinand cont de varsta specifica a fecunditatii si rata de supravietuire, observata in conditii distincte (rata fecunditatii se considera proportia in care sunt depuse ouale: sunt inca discutii daca fecunditatea reprezinta numarul de oua depuse de o femela sau reprezinta numarul de oua depuse + numarul de oua din ovare, inregistrate la moarte). Considerand fecunditatea ca numar de oua depuse, apar alte aspecte: unele sunt nefertile, altele nu eclozeaza (in acest caz, acestea vor fi incluse in rata mortalitatii stadiului de ou). De fapt, unii ecologi si folosesc expresia de rata fertilitatii pentru rata natalitatii. Apoi, fecunditatea si rata de supravietuire variaza cu densitatea, deci valoarea lui r se poate calcula si la diferite densitati. Semnificatia particulara a lui r este in situatia unei fecunditati si rate de supravietuire, maxime, atunci si densitatea este optima si exista un maxim posibil al ratei de crestere, in cadrul unor conditii fizice definite. Aceasta zona poate fi considerata ca o zona optima (acceptiunea lui optim este uneori arbitrara; optim nu implica intotdeauna si un avantaj in crestere, la un maxim posibil al ratei). Rata maxima intrinseca de crestere are importanta din doua puncte de vedere:
- necesara 'ca termen', in ecuatiile matematice ale populatiilor (Lotka, Gausse, Voltera);
-importanta practica (pentru studiul temperaturii, umiditatii, densitatii, competitiei interspecifice, care modeleaza zona optima)
☻- rata finita de crestere (λ), este necesara in studiul populatiilor de insecte, ea reprezentand numarul de timpi necesari pentru multiplicarea populatiei pe o anumita unitate de timp.
Deci, rata finita (limitata ) de crestere este antilogaritmul natural al ratei intrinseci de crestere (r) sau reciproca, rata intrinseca de crestere este logaritmul natural al lui λ.
Cresterea exponentiala.
Daca in timpul 0 se porneste de la un individ care se tot multiplica din timp in timp (1, 2, 4, 8, 16, 32, 64, 128.), multiplicarea este de tip exponential. Daca intr-o populatie exista acest tip de crestere, in care populatia creste mereu, este vorba de o crestere exponentiala (vezi Fig. 5-25; 5-26).
Acest tip de crestere depinde de potentialul biotic al speciei, de marimea populatiei (cand potentialul este redus si rata de crestere este redusa). Cresterea exponentiala este rara (cazul bacteriilor sau a altor microorganisme).
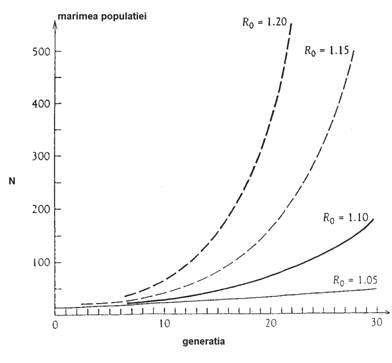
Fig. 5-26. Exemple de crestere exponentiala
(geometrica) a populatiei. Generatii distincte, rata reproductiva
Un model de crestere de tip exponential caracterizeaza de exemplu bacteriile si protozoarele. Dar aceasta nu poate decurge la infinit, existand si o rata mare a mortalitatii. Daca nu ar fi asa, celulele de bacterii se pot divide tot la 20 minute, astfel ca in x ore ar atinge un numar de 23x celule. Admitand ca volumul unei bacterii este de 1μ3 in 43 ore s-ar atinge volumul Terrei !.
Logaritmarea ordonatei permite liniarizarea iar forma curbei exponentiale astfel obtinuta este ilustrata in Fig. 5-27.
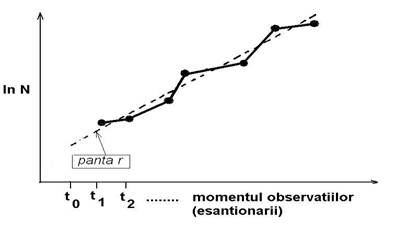
Fig. 5-27. Determinarea directa a ratei de crestere r (panta unei drepte) dupa liniarizarea functiei exponentiale de crestere (dupa Frontier si Pichod-Vial 1998).
2.Cresterea logistica
Se mai numeste si cresterea Verhulst. Cand curba de crestere la multe populatii este lenta - in relatia marimea populatiei si timp - curba care se obtine are forma literei "S" si reprezinta cresterea sigmoida (logistica ) (Fig. 5-28).
Interpretarea simplista este urmatoarea: in prima faza cresterea este activa, de tip exponential; apoi se atenueaza si are loc dupa un model liniar; partea terminala arata o stabilizare, cresterea este de tip logaritmic si arata ca oricat ar creste timpul, populatia nu mai creste sau creste foarte putin.
La limita superioara a curbei unde aceasta are o portiune dreapta, se gaseste capacitatea portanta = capacitate de suportare a mediului = capacitate de sustinere a mediului (carrying capacity) (K), care poate fi interpretata in 4 moduri diferite si reprezinta:
- limita impusa de abilitatea mediului de a suporta populatia la un nivel de supravietuire redus; nu exista suficienta hrana pentru o crestere optima a corpului, vigoarea si sanatatea populatiei; orice schimbare, cat de mica, de la aceasta stare poate duce la catastrofe (Dasmann 1964);
- densitatea optima la care populatia are hrana si alte conditii adecvate; indivizii se apropie de maxim in greutatea corpului, vigoare si sanatate; populatia ramane insa putin timp la acest nivel deoarece apar daunatori iar populatia este controlata de teritorialitate sau de natalitate;
- un prag de securitate; daca densitatea unei populatii este deasupra unui anumit nivel exista o instabilitate in populatie (o parte din indivizi pot pleca din populatie iar altii pot fi afectati de pradatori); in acest caz, densitatea de securitate este corespunzatoare unei densitati tolerante, or unei densitati optime (Errington 1946);
- densitate toleranta; este o densitate de saturatie (numarul sau biomasa de organisme pe care le poate suporta un anumit habitat) (Odum 1993).
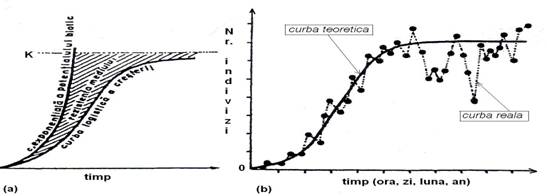
Fig. 5-28. (a). Reprezentarea schematica a modelului curbei logistice de crestere si a curbei exponentiale a potentialului biotic. (b). Curba logistica, teoretica si reala (dupa Stan 1994 (1995).
Aplicarea acestui concept, la diferite nivele de organizare ierarhica in ecologie (populatie, biocenoza, ecosistem) este totusi foarte dificila. Se pune des intrebarea care poate fi limita superioara a lui K in conditiile cresterii unei populatii care poate fi sustinuta de un mediu de viata dat ?. Specialistii iau in considerare existenta a doua nivele: (a) un maxim sau o densistate de subzistenta sau un maxim al numarului de indivizi care poate sa-si prelungasca existenta in habitat; (b) un nivel optim sau "sigur" la o densitate mai mica si la care exista o mai mare siguranta pentru indivizi, in ceea ce priveste hrana, rezistenta la pradatori sau fluctuatia periodica a resurselor (Odum 1993). Daca urmarim curba cresterii logistice, unde este marcat K ca reprezentand nivelul maxim, un nivel optim poate fi apreciat ca localizat intre acest maxim si un punct de inflexiune unde rata cresterii populatiei este cea mai mare iar densitatea este cam jumatate sau 2/3 ca marime.
Ca exemplu oate fi citat cazul speciilor de Cervidae (Cervus sp., Capreolus sp. etc) Astfel, populatia atinge limita K, in absenta pradatorilor, printr-o inmultire rapida; rezultatul acestei cresteri este un pasunat excesiv iar vegetatia nu mai poate oferi un suport de existenta ca in cazul populatiei incipiente sau anterioare. Experimental s-a trecut apoi la o mutare de exemplare sau printr-un fenomen experimental de predatorism pana la o rata care sa mentina populatia la un nivel optim. Pe baza unor studii de durata s-a evidentiat ca valoarea pentru K maxim este de 90 indivizi/mila2, iar K optim de 50 indivizi/mila2 (Mc Cullough 1979).
Aceste principii sau concepte sunt valabile atat pentru populatiile de animale cat si pentru cele de plante. Astfel, in cazul unor specii de plante (ex. Pinus sp.) un numar de 100 exemplare au crescut relativ bine impreuna pe un lot experimental. Cu timpul insa, cresterea s-a incetinit si a crescut mortalitatea. Reducerea densitatii la ½ a modificat radical situatia; a crescut calitatea si fiecare individ a prezentat o valoare economica mare.
In impactul lor cu mediul insa, reactia indivizilor se poate inscrie pe un spectru larg. Fenomenul este cu atat mai vizibil si interesant referitor la populatia umana. In azul acesteia, pe langa resursele ca de suport de viata mai intervine un al doilea parametru - intensitatea de folosinta. In acest caz, capacitatea de suportare a mediului (K), in cazul populatiei umane, a fost definita ca volumul si intensitatea de folosinta a resurselor (Catton 1987) (autorul sugereaza chiar legarea conceptului K de Legea minimului lui Liebig, pentru estimarea limitelor superioare).
Atentie.
Este important de retinut ca in ceea
ce priveste capacitatea de suportare a mediului (K), aceasta nu este o
In cazul ecosistemelor naturale, la nivel global, capacitatea de suportare a mediului (K) ar putea fi aproape de o constanta, atat timp cat nivelul si calitatea energiei solare (input) ramane constanta.
Cele 2 relatii importante ale cresterii populatiilor sunt:
a. pentru
cresterea exponentiala: ![]() (1)
(1)
b. pentru
cresterea logistica (ecuata lui Verhulst): ![]() (2)
(2)
Formele diferentiale ale acestor ecuatii pot fi prezentate in urmatoarea forma:
(1): rata de crestere = rata reproductiva r ∙ numar N
(2): rata de crestere = rata reproductiva r ∙ numar N ∙ factorul propriu de limitare (rezistenta mediului)- [(K-N)/K]
Cele doua constante importante sunt: K - nivelul capacitatii superioare de suportare a mediului si r - rata intrinseca de crestere a populatiei intr-un mediu nelimitat.
r-K selection. In medii de viata in care nu exista o aglomerare de indivizi ai vreunei specii (ex. cazul colonizarii unui agroecosistem dupa ce o anume cultura a fost abandonata, popularea incipienta a unor noi medii de viata etc), presiunea selectiei naturale va favoriza intotdeauna speciile cu un potential mare al ratei reproductive, deci cu un numar mare de urmasi. Opus, in medii aglomerate (ex. o padure matura si stabila) selectia naturala va favoriza speciile cu rata redusa de crestere, dar adaptate bine la aceste conditii si capabile sa utilizeze cu eficienta maxima resursele disponibile. Ceeea ce am prezentat succint aici este cunoscutul principiu al selectiei r-K (r-K selection), situatii in care speciile manifesta o strategie r ("generalisti") sau o strategie K ("specialisti") (Odum 1993; Rejmanek si Spitzer 1980; Spitzer si Rejmanek 1982)
Relatia dintre cresterea populatiei si densitate.
Un prim caz este acela in care, cu cat densitatea creste, rata cresterii descreste. Este tipul de relatie invers dependenta de densitate (Fig. 5-29).
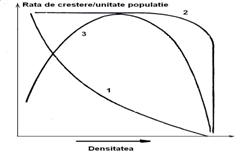
Fig. 5-29. Trei modele ale cresterii populatiei in relatie cu densitatea (curba 1 = tipul de relatie invers dependenta de densitate; curba 2 = tipul independent de densitate; curba 3 - tipul Allee (dupa Odum 1993).
O alta situatie este aceea cand rata cresterii ramane la nivel ridicat pana ce densitatea va creste de asemenea la un nivel mare si apar alti factori externi limitativi. Cand exista o serie de factori externi a caror actiune este relativ slaba (ex. pradatori, lipsa de hrana sau hrana putina, factori fizici ai mediului etc), aceste specii pot suferii oscilatii pronuntate ale densitatii si cresterii, cu efecte imprevizibile sau negative asupra factorului uman. Este cazul speciilor care in literatura de specialitate sunt considerate "daunatoare". Unele pot fi daunatori permanenti datorita unui nivel constant mare al populatiilor. Sunt daunatori tipici, mai ales in cazul acelor specii unde planta gazda este o planta cultivata. Alte specii au densitati relativ constant-reduse dar sunt tot daunatoare (ex. lepidopterul Mamestra brassicae). Un alt caz este acela al unor specii cu nivel constant mare al populatiilor dar nu produc daune decat cu totul intamplator, dar cand densitatea creste exploziv pot fi daunatori ocazionali. Este cazul multor specii extrem polifage (ex. Xestia c-nigrum) Acesta este tipul independent de densitate (Fig. 5-29; curba 2).
In cazul speciilor de animale sociale sau plante coloniale, rata cresterii atinge nivelul cel mai mare la o densitate intermediara. Atat densitatea mica cat si cea mare determina o scadere a ratei de crestere. Este asa numita forma de crestere a lui Allee (Allee 1951) (Fig. 5-29; curba 3).
Mai este cunoscut cazul speciilor caracterizate de explozii populationale (dupa terminologia "boom and bust"). Adica, are loc o crestere exploziva, cu atingerea unei densitati mari, dupa care se constata o revenire la normal printr-o scadere a nivelului populatiilor, la fel de pronuntata (ex. ciclul de 4 ani al lemingilor, unele specii de insecte - gandacii de scoarta, lacustele etc).
Curba logistica este o curba teoretica. Curba dupa care are loc relatia dintre numarul de indivizi si timp are in realitate un trend ferestruit. Aceasta este curba reala. Intervalul (spatiul) dintre curba exponentiala a potentialului biotic si curba logistica reprezinta rezistenta mediului (Fig. 5-28).
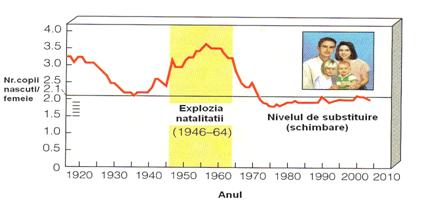
Cresterea populatiei umane. Populatia umana a cunoscut si modele extreme de crestere. Un model de crestere negativa a fost inregistrat prin sec. XIV, cand ciuma bubonica ("moartea neagra") a redus populatia Europei cu 25% (Freedman si Berelson 1974). In anumite secole, populatia umana a crescut incet, comparativ cu altele. Un model de acest gen este prezentat in Fig. 5-31.
Se observa ca in perioada 1930-1940 a urmat un declin dupa care o explozie demografica pe intervalul 1946-1964, iar aceasta a fost continuata de o stabilizare (nivel de substituire), pe perioada 1975-2005.
Evolutia ratei natalitatii si mortalitatii pentru populatia umana din diferite tari si zone de pe glob arata ca rata natalitatii este deosebit de mare tocmai in tarile foarte sarace si scade progresiv odata cu crestere nivelului de trai, gradul de civilizatie si puterea economica, industriala (Fig. 5-31; 5-32-B).
Perioadele de crestere accentuata si rapida au fost legate de procurarea resurselor de energie:
- prima crestere pronuntata a fost dependenta de dezvoltarea agriculturii (ceea ce a determinat si o crestere a capacitatii de suportare a mediului, fiind realizata acum circa 8000 de ani);
- a doua a fost asociata de revolutia industriala (acum circa 200 ani; ceea ce a determinat o colonizare sustinuta si accentuata a continentelor, descresterea mortalitatii si cresterea numarului de membrii prin cresterea ratei natalitatii si prelungirea duratei de viata ca rezultat al progreselor din medicina si sanatate) (procentajul de crestere al populatiei umane a fost de 80%, numai in ultimele 2 secole; azi atinge 5-6 miliarde locuitori/Terra; demografi cu experienta sustin ca in aceste conditii ale unei cresteri logistice, in urmatorul secol, populatia va fi de 9-14 miliarde locuitori !).
 O
modalitate sugerata de specialisti pentru a descrie cresterea populatiei umane
este termenul de dublarea timpului (= numarul de ani necesari pentru dublarea
densitatii). Cresterea anuala este exprimata in numarul de persoane adagate la
o suta existenta (sau exprimata procentual) (ex. o crestere de 2 procente
annual inseamna 2 persoane la 100, respectiv 200 la 1000 deja existente).
Pentru estimarea aproximativa a dublarii timpului t, pentru o rata de crestere r
de 2% (exprimata in fractie zecimala), exista urmatoarea relatie:
O
modalitate sugerata de specialisti pentru a descrie cresterea populatiei umane
este termenul de dublarea timpului (= numarul de ani necesari pentru dublarea
densitatii). Cresterea anuala este exprimata in numarul de persoane adagate la
o suta existenta (sau exprimata procentual) (ex. o crestere de 2 procente
annual inseamna 2 persoane la 100, respectiv 200 la 1000 deja existente).
Pentru estimarea aproximativa a dublarii timpului t, pentru o rata de crestere r
de 2% (exprimata in fractie zecimala), exista urmatoarea relatie: ![]()
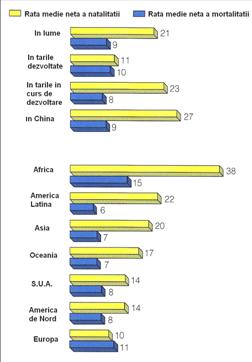
Fig. 5-31. Rata fertilitatii pentru populatia umana din SUA (evaluata pentru perioada 1917-2005) (stanga) si reprezentarea valorilor medii brute ale ratei natalitatii si mortalitatii pentru diferite tari si zone de pe glob (dreapta) (dupa Population Reference Bureau & US Census Bureau; 2006).
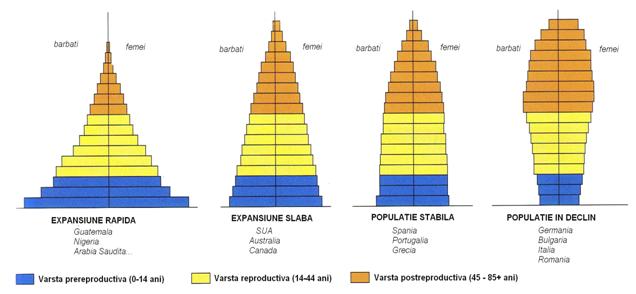
Fig. 5-32-A. O reprezentare sintetica prin diagrame a relatiei varsta - structura populatiei pentru 4 situatii reprezentative: crestere rapida, crestere lenta, stare stabila si populatie in declin (dupa Environmental Science - NowTM; Miller 2006).
Studiul relatiei dintre varsta sexelor pentru 4 categorii reprezentative de evolutie a nivelului populatiei (Fig. 5-32-A) a evidentiat ca tocmai populatiile care au o larga pondere a varstei prereproductive (0-14 ani) sunt si populatiile cu cresterea cea mai rapida a populatiei. Ori acest lucru este caracteristic din pacate tocmai pentru tarile sarace. In tarile cu nivel de viata crescut cresterea este slaba sau stabila iar aici incepe sa domine varsta reproductiva si postreproductiva. Un caz interesant este cel prezentat de a 4-a diagrama, pentru populatiile in declin. Se observa nivelul scazut pentru populatia de varsta prereproductiva, dar o dominare a varstei postreproductive. Aici sunt tarile care au mai multi batrani decat tineri. Sunt tari in curs de dezvoltare iar aici se pare ca exista mai multi factori de influenta.
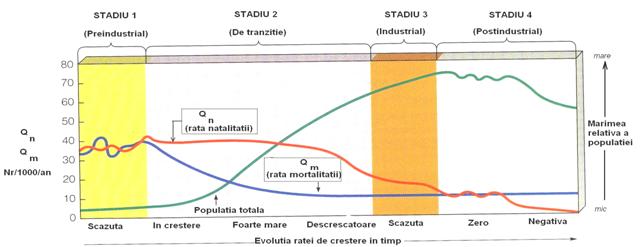
Fig. 5-33. Reprezentarea generalizata a modelului tranzitiei demografice. Este un model care se aplica tarilor avansate din punct de vedere industrial si intelectual. Sunt sugerate 4 stadii iar trendul curbelor care ilustreaza rata natalitatii si rata mortalitatii, este deosebit de interesant (dupa Environmental Science - NowTM; Miller 2006).
Un procent de 2% nu pare a fi prea mult, dar la aceasta rata inseamna ca populatia orasului Cluj, creste de 2 ori pe durata de viata a unei persoane (la ora actuala, pe Terra, rata de crestere este de 1,8%; in unele tari subdezvoltate este de 3% ceea ce inseamna ca timpul de dublare va fi peste 23 de ani !; in tarile dezvoltate si supradezvoltate, rata este <1% iar dublarea timpului va fi peste 70 ani sau mai mult; la polul opus sunt unele tari unde rata este in jur de 0% !). La nivel mondial, rata de crestere este cuprinsa intre una joasa (o medie in jur de 0,8%) si una mare (media de 2,5%) (Brown si Jacobson 1986).
Legat de cresterea populatiei umane sunt o serie de pareri contraversate. Unii sociologi, economisti si ecologi sustin fenomenul de tranzitie demografica (Fig. 5-33) (pornind de la ideea ca populatia umana va incetini ritmul de crestere ca popor fiind in viitor mai putin dependenta de copii lor; va creste tendinta de a avea putini copii si a folosi resursele pentru cresterea calitatii vietii - adica a trecere spre un stil de viata de forma K). Din acest punct de vedere, cresterea populatiei este o problema de economie (McNamara 1982). Se para insa ca lucrurile sunt mult mai complicate, fiind importante nu numai aspectele economice dar si cele de dezvoltare calitativa, de constiinta si educatie, de controlul natalitatii etc (Brown si Johnson 1986).
Cresterea Von Bertalanffy
Este ilustrata de ecuati von Bertalanffy si ecuatia Ivlev. Este o ecuatie folosita in cazul cresterii individuale si in particular in cazul cresterii in lungime. In acest caz, din curba logistica se ia in considerare doar portiunea dreapta (exponentiala negativa). Aceasta este de forma:
Cresterea Gompertz
Reprezinta o curba de forma celei logistice doar ca este disimetrica, edificata cu ecuatia Gompertz. In partea sa initiala prezina o crestere de tip exponential pozitiv, de durata insa foarte scurta comparativ cu restul. Punctul de inflexiune este atins la un nivel inferior care reprezinta jumatatea nivelului limita maxima (Fig. 5-35, a).
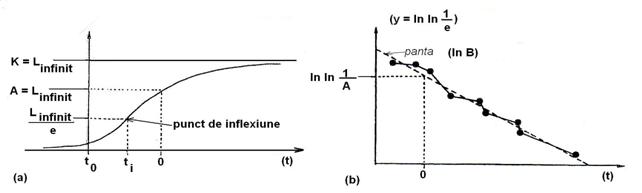
Fig. 5-35. Curba normala de crestere Gompertz (a) si reprezentarea liniarizata a curbei prin logaritmare (dupa Frontier si Pichod-Vial 1998).
|
Politica de confidentialitate | Termeni si conditii de utilizare |

Vizualizari: 3745
Importanta: ![]()
Termeni si conditii de utilizare | Contact
© SCRIGROUP 2025 . All rights reserved